Review

ABSTRACT
INTRODUCTION
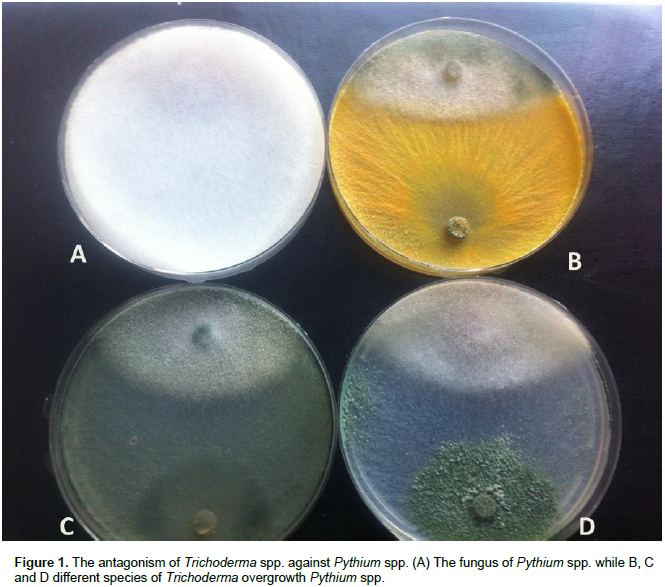
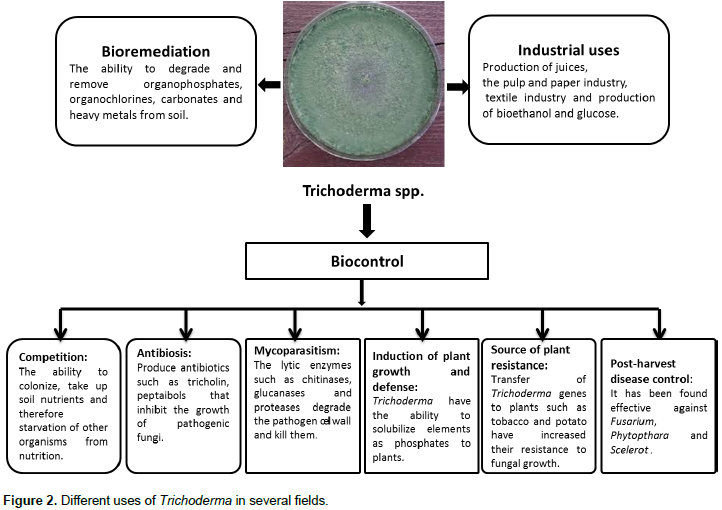
OTHER APPLICATIONS OF TRICHODERMA
TOOLS FOR GENETIC IMPROVEMENT OF TRICHODERMA
CONCLUSION
CONFLICT OF INTERESTS
REFERENCES
|
Abbasi S, Safaie N, Shamsbakhsh M (2014). Evaluation of gamma-induced mutants of Trichoderma harzianum for biological control of charcoal rot of melon (Macrophomina phaseolina) in laboratory and greenhouse conditions. J. Crop. Prot. 3 (4):509-521. |
|
|
Agrawal T, Kotasthane AS (2012). Chitinolytic assay of indigenous Trichoderma isolates collected from different geographical locations of Chhattisgarh in Central India. Springerplus. 1:73. |
|
|
Arfarita N, Imai T, Kanno A, Yarimizu T, Xiaofeng S, Jie W, et al (2013). The Potential use of Trichoderma Viride Strain FRP3 in Biodegradation of the Herbicid Glyphosate. Biotechnol. Biotechnol. Equip. 27:3518-3521. |
|
|
Babu AG, Shim J, Bang KS, Shea PJ, Oh BT (2014). Trichoderma virens PDR-28: a heavy metal-tolerant and plant growth- promoting fungus for remediation and bioenergy crop production on mine tailing soil. J. Environ. Manage. 132:129-134. |
|
|
Baharvand A, Shahbazi S, Afsharmanesh H, Ebrahimi MA, Askari H (2014). Investigation of gamma irradiation on morphological characteristics and antagonist potential of Trichoderma viride against M.phaseolina. Intl. J. Farm. Alli. Sci. 3(11):1157-1164. |
|
|
Balasubramanian N, Priya VT, Gomathinayagam S, Lalithakumari D (2012). Fusant Trichoderma HF9 with Enhanced Extracellular Chitinase and Protein Content. Appl. Biochem. Microbiol. 48:409-415. |
|
|
Bareen FE, Shafiq M, Jamil S (2012). Role of plant growth regulators and a saprobic fungus in enhancement of metal phytoextraction potential and stress alleviation in pearl millet. J. Hazard. Mater. 237-238:186-193. |
|
|
Bartinicki-Garcia S (1968). Cell wall chemistry, morphogenesis and taxonomy of fungi. Annu. Rev. Microbiol. 22:87-109. |
|
|
Benítez T, Rincón AM, Limón MC, Codón AC (2004). Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 7:249-260 |
|
|
Bisby GR (1939). Trichoderma viride Pers. ex Fries and notes on Hypocrea. Trans. Br. Mycol. Soc. 23(2):149-168. |
|
|
Błaszczyk L, Siwulski M, Sobieralski K, Lisiecka J, Jędryczka M (2014). Trichoderma spp. application and prospects for use in organic farming and industry. J. Plant. Protect. Res. 54(4). |
|
|
Brunner K, Zeilinger S, Ciliento R, Woo SL, Lorito M, Kubicek CP, et al. (2005). Improvement of the Fungal Biocontrol Agent Trichoderma atroviride To Enhance both Antagonism and Induction of Plant Systemic Disease Resistance. Appl. Environ. Microbiol. 71:3959-3965. |
|
|
Chet I, Inbar J (1994). Biological control of fungal pathogens. Appl. Biochem. Biotechnol. 48:37-43. |
|
|
Chet I, Inbar J, Hadar I (1997). Fungal antagonists and mycoparasites. In: Wicklow DT, Söderström B (eds) The Mycota IV: Environmental and microbial relationships. Springer-Verlag, Berlin. pp. 165-184. |
|
|
Dana MM, Pintor-Toro JA, Cubero B (2006). Transgenic Tobacco Plants Overexpressing Chitinases of Fungal Origin Show Enhanced Resistance to Biotic and Abiotic Stress Agents. Plant. Physiol. 142:722-730. |
|
|
El-Hassan SA, Gowen SR, Barbara Pembroke (2013). Use of Trichoderma hamatum for biocontrol of lentil vascular wilt disease: efficacy, mechanisms of interaction and future prospects. J. Plant. Prot. Res. 53(1). |
|
|
Farkas V (1990). Fungal cell walls: their structure, biosynthsis and biotechnological aspects . Acta. Biotechnol. 10:225-238. |
|
|
Ferre FS, MP Santamarina (2010). Efficacy of Trichoderma harzianum in suppression of Fusarium culmorum. Ann. Microbiol. 60:335-340. |
|
|
Gajera H, Domadiya R, Patel S, Kapopara M, Golakiya B (2013). Molecular mechanism of Trichoderma as bio-control agents against phytopathogen system. Curr. Res. Microbiol. Biotechnol. 1(4):133-142 |
|
|
Ghisalberti EL, Narbey MJ, Dewan MM, Sivasithamparam K (1990). Variability among strains of Trichoderma harzianum in their ability to reduce take-all and to produce pyrones. Plant. Soil. 121:287-291. |
|
|
Giczey G, KereËnyi Z, Dallmann G, ËHornok L (1998). Homologous transformation of Trichoderma hamatum with an endochitinase encoding gene, resulting in increased levels of chitinase activity. FEMS. Microbiol. Lett. 165:247-252. |
|
|
Harmam GE, Petzoldt R, Comis A, Chen J (2004a). Interactions Between Trichoderma harzianum Strain T22 and Maize Inbred Line Mo17 and Effects of These Interactions on Diseases Caused by Pythium ultimum and Colletotrichum graminicola. Phytopathology 94:147-153. |
|
|
Harman GE, Howell CR, Viterbo A, Chet I, Lorito M (2004b). Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2(1):43-56. |
|
|
Harris AD, Ramalingam C (2010). Xylanases and its Application in Food Industry. J. Exp. Sci. 1(7):01-11. |
|
|
Hasan S (2016). Potential of Trichoderma sp. In Bioremediation: A Review. J. Basic. Appl. Eng. Res. 776-779. |
|
|
Hermosa R, Viterbo A, Chet I, Monte E (2012). Plant-beneficial effects of Trichoderma and of its genes. Microbiology 158:17-25. |
|
|
Howell CR (1999). Selective isolation from soil and separation in vitro of P and Q strains of Trichoderma virens with differential media. Mycologia 91:930-4. |
|
|
Howell CR (2003). Mechanisms employed by Trichoderma species in the biological control of plant diseases: the history and evolution of current concepts. Plant. Dis. 87:4-10. |
|
|
Junaid JM, Dar NA, Bhat TA, Bhat AH, Bhat MA (2013). Commercial Biocontrol Agents and Their Mechanism of Action in the Management of Plant Pathogens. Int. J. Mod. Plant. Anim. Sci. 1(2):39-57. |
|
|
Khandoker N, Al Mamun A, Nafiz TN, Khan SN, Hoq MM (2013). Strain Improvement of Trichoderma Viride through Mutation for Enhanced Production of Cellulase. Bangladesh. J. Microbiol. 30:43-47. |
|
|
Kowsari M, Motallebi M, Zamani RM (2014a). Construction of new GFP-tagged fusants for Trichoderma harzianum with enhanced biocontrol activit. J. Plant. Protect. Res. 54(2). |
|
|
Kowsari M, Motallebi M, Zamani M (2014b). Protein Engineering of Chit42 Towards Improvement of Chitinase and Antifungal Activities. Curr. Microbiol. 68:495-502. |
|
|
Kumar S (2013). Trichoderma: a biological weapon for managing plant diseases and promoting sustainability. Int. J. Agric. Sci. Vet. Med. 106-121. |
|
|
Limon MC, Pintor-Toro JA, Benítez T (1999). Increased antifungal activity of Trichoderma harzianum transformants that overexpress a 33- kDa chitinase, Phytopathology 89:254-261. |
|
|
Lorito M, Woo SL, Fernandez IG, Coluccii G, Harman GE, Pintor-toro JA, Filipponei E, Muccifora S, Lawrence CB, et al. (1998). Genes from mycoparasitic fungi as a source for improving plant resistance to fungal pathogens. Proc. Natl. Acad. Sci. 95:7860-7865. |
|
|
Mayo S, Gutiérrez S, Malmierca MG, Lorenzana A, Campelo MP, Hermosa R, et al (2015) Influence of Rhizoctonia solani and Trichoderma spp. in growth of bean (Phaseolus vulgaris L.) and in the induction of plant defense-related genes. Front. Plant Sci. 6:685. |
|
|
Mendoza-Mendoza A, Pozo MJ, Grzegorski D, Martınez P, Garcıa JM, Olmedo-Monfil V, Cortes C, Kenerley C, Herrera-Estrella A (2003) Enhanced biocontrol activity of Trichoderma through inactivation of a mitogen-activated protein kinase. PNAS. 100(26):15965-15970. |
|
|
Mohamed HAA, Haggag WM (2010). Mutagenesis and inter-specific protoplast fusion between Trichoderma koningii and Trichoderma reesei for biocontrol improvement. Am. J. Sci. Ind. Res. 1(3):504-515. |
|
|
Mukherjee PK (2011) Genomics of biological control – whole genome sequencing of two mycoparasitic Trichoderma spp. Curr. Sci. 101(3):268 |
|
|
Naher L, Yusuf U, Ismail A, Hossain k (2014). Trichoderma SPP: A biocontrol agent for sustainable management of plant diseases. Pak. J. Bot. 46(4):1489-1493. |
|
|
Nevalainen H, Penttilä M (1995). Molecular biology of cellulolytic fungi. In: The Mycota II. Genetics and Biotechnolog. Edt. U. Kuck. Springer-Verlag, Berlin, pp. 303-319. |
|
|
Pal KK, Gardener BM (2006). Biological Control of Plant Pathogens. The Plant Health Instructor, Pandya JR, Sabalpara AN, Chawda SK (2011). Trichoderma: a particular weapon for biological control of Phytopathogens. J. Agric. Technol. 7(5):1187-1191. |
|
|
Patil AS, Lunge AG (2012). Strain improvement of Trichoderma |
|
|
harzianum by UV mutagenesis for enhancing its biocontrol potential against aflotoxigenic Aspergillus species. Experiment 4:228-242. |
|
|
Persoon CH(1794). Neuer Veersuch einer systematischen Eintheilung der Schwamme. Neues Magazin für die Botanik. 1:63-128. |
|
|
Polizeli MLTM, Rizzatti ACS, Monti R, Terenzi HF, Jorge JA, Amorim DS (2005). Xylanases from fungi: properties and industrial applications. Appl. Microbiol. Biotechnol. 67:577-591. |
|
|
Prabavathy VR, Mathivanan N, Sagadevan E, Murugesan K, Lalithakumari D (2006). Self-fusion of protoplasts enhances chitinase production and biocontrol activity in Trichoderma harzianum. Bioresour. Technol. 97:2330-2334. |
|
|
Rashmi S, Maurya S, Upadhyay RS (2016). The improvement of competitive saprophytic capabilities of Trichoderma species through the use of chemical mutagens. Braz. J. Microbiol. 47(1):10-17. |
|
|
Rao GS, Reddy NNR, Surekha C (2015). Induction of plant systemic resistance in legumes Cajanus cajan, Vignaradiata, Vigna mungo against plant pathogens Fusarium oxysporum and Alternaria alternata - a Trichoderma viride mediated reprogramming of plant defense mechanism. Int. J. Rec. Sci. Res.6:4270-428. |
|
|
Rifai MA (1969) A revision of the genus Trichoderma. Mycol. Pap. 116:1-56. |
|
|
Sadykova VS, Kurakov AV, Kuvarina AE, Rogozhin EA (2015). Antimicrobial Activity of Fungi Strains of Trichoderma from Middle Siberia. Appl. Biochem. Microbiol. 51:355-361. |
|
|
Saravanakumar K, Fan L, Fu K, Yu C, Wang M, Xia H, Sun J, Li Y, Chen J (2016). Cellulase from Trichoderma harzianum interacts with roots and triggers induced systemic resistance to foliar disease in maize. Sci. Rep. 6:35543. |
|
|
Seidl V, Seiboth B (2010). Trichoderma reesei: genetic approaches to improving strain efficiency. Biofuels 1(2):343-354. |
|
|
Sharma RA (2012). brief review on mechanism of Trichoderma fungus use as biological control agents. Int. J. Innov. Bio-Sci. 2: 200-210. |
|
|
Shi M, Chen L, Wang XW, Zhang T, Zhao PB, Song XY, Sun CY, Chen XL, Zhou BC, Zhang YZ. (2012). Antimicrobial peptaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 158:166-175. |
|
|
Singh R, Maurya S, Upadhyay RS (2016). The improvement Of competitive saprophytic capabilities of Trichoderma species through the use of chemical mutagens. Braz. J. Microbial. 47:10-17. |
|
|
Singh R, Maurya S, Upadhyay RS (2010). Improvement of antagonistic capability of Trichoderma harzianum by UV irradiation for management of Macrophomina phaseolina. Arch. Phytopathol. Plant. Prot. 43:1579-1588. |
|
|
Sivan A, Chet I (1989). Biological control of Fusarium spp. in cotton, wheat and muskmelon by Trichoderma harzianum. J. Phytopathol. 116:39-47. |
|
|
Solgi T, Moradyar M, Zamani MR, Motallebi M (2015). Transformation of canola by chit33 gene towards improving resistance to Sclerotinia sclerotiorum. Plant. Protect. Sci. 51:6-12. |
|
|
Srivastava PK, Vaish A, Dwivedi S, Chakrabarty D, Singh N, Tripathi RD (2011). Biological removal of arsenic pollution by soil fungi. Sci. Total Environ. 409(12):2430-2442 |
|
|
Teng Y, Luo Y, Ma W, Zhu L, Ren W, Luo Y, et al (2015). Trichoderma reesei FS10-C enhances phytoremediation of Cd-contaminated soil by Sedum plumbizincicola and associated soil microbial activities. Front. Plant Sci. 6:438. |
|
|
Vinale F, Sivasithamparamb K, Ghisalbertic EL, Marraa R, Woo SL, Loritoa M (2008). Trichoderma–plant–pathogen interactions. Soil. Biol. Biochem. 40:1-10. |
|
|
Vinale F, Sivasithamparam K, Ghisalberti EL, Woo SL, Nigro M, Marra R, et al (2014). Trichoderma Secondary Metabolites Active on Plants and Fungal Pathogens. Mycol. J. 8:127-139. |
|
|
Walunj AA, John P (2015). Adaptability of UV Irradiated mutant of Trichoderma harizianum to carbendazim. The Bioscan 10(4):1869-1871. |
|
|
Weindling R (1932). Trichoderma lignorum as a parasite of other fungi. Phytopathology 22:837. |
|
|
Woo SL, Ruocco M, Vinale F, Nigro M, Marra R, Lombardi N, et al (2014). Trichoderma-based Products and their Widespread Use in Agriculture. The Open Mycol. J. (Suppl-1, M4):71-126 71. |
|
|
Yazdani M, Yap CK, Abdullah F, Tan SG (2009) Trichoderma atroviride as a bioremediator of Cu pollution: an in vitro study. Toxicol. Environ. Chem. 91(7):1305-1314. |
|
|
Zeng X, Su S, Jiang X, Li L, Bai L, Zhang Y (2010). Capability of pentavalent and Biovolatilization of Three Fungal Strains under Laboratory Conditions. Clean. Soil. Air. Water. 38: 238-241. |
|
|
Zhang F, Ruan X, Wang X, Liu Z, Hu L, Li C (2016). Overexpression of a Chitinase Gene from Trichoderma Asperellum Increases Disease Resistance in Transgenic Soybean. Appl. Biochem. Biotechnol. 180:1542-1558. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


