Review

ABSTRACT
INTRODUCTION
PLANT ALUMINIUM ION TOXICITY AVOIDANCE MECHANISMS
POSSIBLE ALUMINIUM TOXICITY, HETEROTIC AND EPIGENETIC MECHANISM MODELS
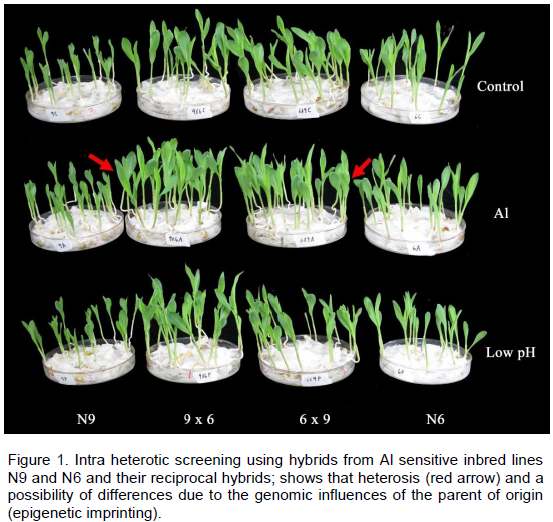
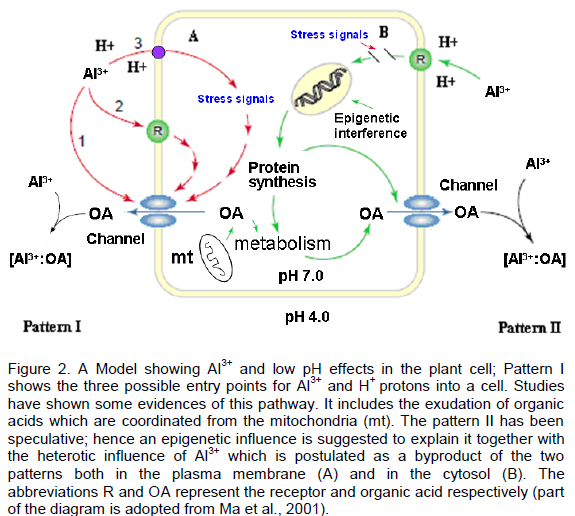
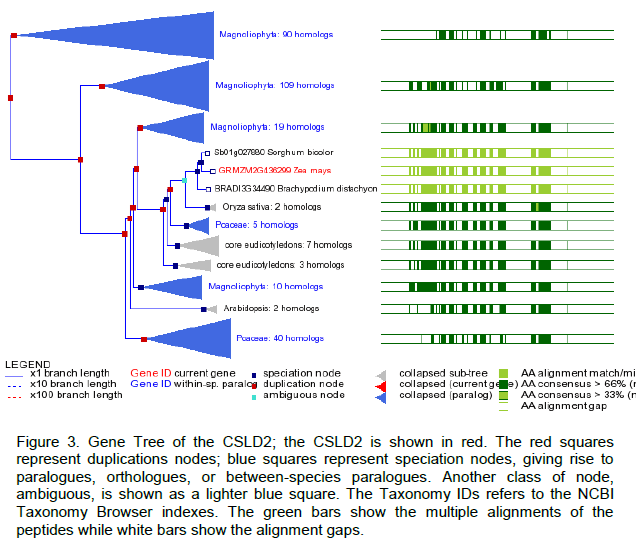
CONFLICT OF INTERESTS
The author(s) did not declare any conflict of interest.
REFERENCES
|
Abate E, Hussien S, Laing M, Mengistu F (2013). Aluminium toxicity tolerance in cereals: Mechanisms, genetic control and breeding methods. Afr. J. Agric. Res. 8(9):711-722. |
|
|
|
|
|
Alleman M, Sidorenko L, McGinnis K, et al. (2006). An RNA dependent RNA polymerase is required for paramutation in maize. Nature 442: 295-298. |
|
|
|
|
|
Bennetzen JL, Hake SC (2009). (eds.), Handbook of Maize: Its Biology, Springer Science, p. 291. |
|
|
|
|
|
Borrero JC, Pandey S, Ceballos H, et al. (1995). Genetic variances for tolerance to soil acidity in a tropical maize population. Maydica 40: 283-288. |
|
|
|
|
|
Chinnusamy V, Zhu J (2009). Epigenetic regulation of stress responses in plants. Curr. Opin. Plant Biol. 12:1-7. |
|
|
|
|
|
Cubas P, Vincent C, Coen E (1999). An epigenetic mutation responsible for natural variation in floral symmetry. Nature 401: 157-161. |
|
|
|
|
|
Dana CD, Radhika D, Jennifer RW, et al. (2007). Metastable Epialleles, Imprinting, and the fetal origins of adult diseases. Pediatr. Res. 61: 30-37. |
|
|
|
|
|
Delhaize E, Ryan PR (1995). Aluminum toxicity and tolerance in plants. Plant Physiol. 107:315–321. |
|
|
|
|
|
Duvick DN, Cassman KG (1999). Post-green revolution trends in yield potential of temperate maize in the North-Central United States. Crop Sci. 39: 1622-1630. |
|
|
|
|
|
Eckardt NA (2009). Deep sequencing maps the maize epigenomic landscape. Plant Cell 21:1024-1026. |
|
|
|
|
|
Ezaki B, Tsugita S, Matsumoto H (1996). Expression of a moderately anionic peroxidase is induced by aluminum treatment in tobacco cells: possible involvement of peroxidase isozymes in aluminum ion stress. Physiol. Plant 96:21-28. |
|
|
|
|
|
Fieldes M, Schaeffer S, Krech M, et al. (2005). DNA hypomethylation in 5-azacytidine-induced early flowering lines of flax. Theor. Appl. Genet. 11:136-149. |
|
|
|
|
|
Guo M, Rupe MA, Yang X, Crasta O, Zinselmeier C, Smith OS, Bowen B (2006). Genome-wide transcript analysis of maize hybrids: allelic additive gene expression and yield heterosis. Theor. Appl. Genet. 113:831-845. |
|
|
|
|
|
Hamel F, Breton C, Houde M (1998). Isolation and characterization of wheat aluminum-regulated genes: possible involvement of aluminum as a pathogenesis response elicitor. Planta 205:531-538. |
|
|
|
|
|
He G, He H, Deng XW (2013). Epigenetic Variations in Plant Hybrids and Their Potential Roles in Heterosis. J. Genet. Genomics 40(5): 205-210. |
|
|
|
|
|
Jiang H, Kang DC, Alexandre D, Fisher PB (2000). RaSH, a rapid subtraction hybridization approach for identifying and cloning differentially expressed genes. Proc. Natl. Acad. Sci. USA 97: 12684-12689. |
|
|
|
|
|
Jones DL, Kochian LV, Gilroy S (1998). Aluminum induces a decrease in cytosolic calcium concentration in BY-2 tobacco cell cultures. Plant Physiol. 116:81-89. |
|
|
|
|
|
Kakutani T (2002). Epi-alleles in plants: inheritance of epigenetic information over generations. Plant Cell Physio. 43:1106–1111. |
|
|
|
|
|
Kimatu JN, Jiang L, Ngezahayo F, Songdi C, Quan-yuan Y, Pang J, Liu B (2013). Alteration in cytosine DNA methylation patterns and levels induced by aluminium toxicity stress in Maize varieties. Int. J. Mod. Agric. 2(1):9-25. |
|
|
|
|
|
Kinraide T, Parker D (1989). Assessing the phytotoxicity of mononuclear hydroxy-aluminum. Plant Cell Environ. 12:478-487. |
|
|
|
|
|
Kisinyo PO, Opala PA, Gudu SO, Othieno CO, Okalebo JR, Palapala V, Otinga AN (2014). Recent advances towards understanding and managing Kenya acid soils for improved crop production. Adv. J. Agric. Res. 9(31):2397-2408. |
|
|
|
|
|
Kochian LV, Hoekenga OA, Pi-eros MA (2004). How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu. Rev. Plant Biol. 55:459-493. |
|
|
|
|
|
Kochian LV (1995). Cellular mechanisms of aluminum toxicity and resistance in plants. Annu. Rev. Plant Biol. 46:237-260. |
|
|
|
|
|
Kollmeier M, Dietrich P, Bauer CS, Horst WJ, Hedrich R (2001). Aluminum activates a citrate-permeable anion channel in the Alsensitive zone of the maize root apex: a comparison between an Al-sensitive and an Altolerant cultivar. Plant Physiol. 126:397-410. |
|
|
|
|
|
Kuleshov NN (1933). World's diversity of phenotypes of maize. J. Agron. 25:688-700. |
|
|
|
|
|
Lukens LN, Zhan S (2007). The plant genome's methylation status and response to stress: implications for plant improvement. Curr. Opin. Plant Biol. 10:317-322. |
|
|
|
|
|
Ma JF, Ryan PR, Delhaize E (2001). Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci. 6:273-278. |
|
|
|
|
|
Magnavaca R, Gardner C, Clark R (1987). Evaluation of inbred maize lines for aluminum tolerance in nutrient solution. In: Gabelman HLB, ed. Genetic aspects of plant mineral nutrition. Dordrecht, the Netherlands: Martinus Nijhoff, pp. 255-265. |
|
|
|
|
|
Mao C, Yi K, Yang L, Zheng B, Wu Y, Liu F, Wu P (2004).Identification of aluminiumregulated genes by cDNA-AFLP in rice (Oryza sativa L.): aluminium-regulated genes for the metabolism of cell wall component. J. Exp. Bot. 55:137.143. |
|
|
|
|
|
Mariano ED, Keltjens WG (2003). Evaluating the role of root citrate exudation as a mechanism of aluminium resistance in maize genotypes. Plant Soil 256:469-479. |
|
|
|
|
|
Maron LG, Kirst M, Mao C, Milner MJ, Menossi M, Kochian LV (2008). Transcriptional profiling of aluminum toxicity and tolerance responses in maize roots. New Phytol. 179:116-128. |
|
|
|
|
|
Mena S, Ortega A, Estrela JM (2009). Oxidative Stress and Mechanisms of Environmental Toxicity. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 674:36-44. |
|
|
|
|
|
Miller TE, Iqbal N, Reader SM et al. (1997). A cytogenetic approach to the improvement of Aluminium tolerance in wheat, in: Putting Plant Physiology on the Map: Genetic Analysis of Development and Adaptive Traits. Proceedings of the Second New Phytologist Symposium, Bangor, UK. 137: 93–98. |
|
|
|
|
|
Mona MA (2008). Physiological Aspects of Aluminium Toxicity on Some Metabolic and Hormonal Contents of Hordeum Vulgare Seedlings. Aust. J. Basic Appl. Sci. 2:549-560. |
|
|
|
|
|
Moore DP (1974). Physiological effects of pH on roots. In: The plant root and the environment [M], Ed. E. W. Carson, University Press of Virginia, Charlottesville, pp. 135-151. |
|
|
|
|
|
Ninamango-Cárdenas FE, Guimaraes CT, Martins PR, et al. (2003). Mapping QTLs for aluminum tolerance in maize. Euphytica 130: 223–232. |
|
|
|
|
|
Pandey S, Ceballos H, Magnavaca R, et al. (1994). Genetics of tolerance to soil acidity in tropical maize. Crop Sci. 34:1511–1514. |
|
|
|
|
|
Pi-eros MA, Shaff JE, Manslank HS, Alves VM, Kochian LV (2005). Aluminum resistance in maize cannot be solely explained by root organic acid exudation; A comparative physiological study. Plant Physiol. 137:231-241. |
|
|
|
|
|
Pi-eros MA, Cançado GMA, Maron LG, Lyi SM, Menossi M, Kochian LV (2007). Not all ALMT1-type transporters mediate aluminum-activated organic acid responses: the case of ZmALMT1 – an anion-selective transporter. Plant J. 53:352-367. |
|
|
|
|
|
Pi-eros MA, Magalhaes JV, Carvalho AVM, Kochian LV (2002). The physiology and biophysics of an aluminum tolerance mechanism based on root citrate exudation in maize. Plant Physiol. 129:1194-1206. |
|
|
|
|
|
Rakyan VK, Chong S, Champ ME, Cuthbert PC, Morgan HD, Luu KV, Whitelaw E (2003). Transgenerational inheritance of epigenetic states at the murine Axin(Fu) allele occurs after maternal and paternal transmission. Proc. Natl. Acad. Sci. USA 100: 2538–2543. |
|
|
|
|
|
Rao IM, Zeigler RS, Vera R, Sarkarung S (1993). Selection and breeding for acid soil tolerance in crops: upland rice and tropical forages as case studies. BioScience 43:454-465. |
|
|
|
|
|
Rapp RA, Wendel JF (2005). Epigenetics and plant evolution. New Phytol. 168:81–91. |
|
|
|
|
|
Richards KD, Schott EJ, Sharma YK, Davis KR, Gardner RC (1998). Aluminum induces oxidative stress genes in Arabidopsis thaliana, Plant Physiol. 116:409-418. |
|
|
|
|
|
Sasaki T, Yamamoto Y, Ezaki B (2004). A wheat gene encoding an aluminum-activated malate transporter. Plant J. 37:645-653. |
|
|
|
|
|
Schmitz RJ, Schultz MD, Urich MA, Nery JR, Pelizzola M, Libiger O, Alix A, McCosh RB., Chen H, Schork NJ, Ecker JR (2013). Patterns of population epigenomic diversity. Nature 495:193-198. |
|
|
|
|
|
Shen H, He H, Li J, Chen W, Wang X, Guo L, Peng Z, He G, Zhong S, Qi Y, Terzaghi W, Deng XW (2012). Genome-wide analysis of DNA methylation and gene expression changes in two Arabidopsis ecotypes and their reciprocal hybrids. Plant Cell 24:875-892. |
|
|
|
|
|
Sivaguru M, Baluška F, Volkmann D, Felle HH, Horst WJ (1999). Impacts of aluminum on the cytoskeleton of the maize root apex. Short-term effects on the distal part of the transition zone. Plant Physiol. 119:1072-1082. |
|
|
|
|
|
Sivaguru M, Horst W (1998). The distal part of the transition zone is the most aluminum-sensitive apical root zone of maize. Plant Physiol. 116:155-163. |
|
|
|
|
|
Tani E, Polidoros AN, Nianiou-Obeidat I, Tsaftaris AS (2005). DNA methylation patterns are differently affected by planting density in maize inbreds and their hybrids. Maydica 50:19-23. |
|
|
|
|
|
Tice KR, Parker DR, DeMason DA (1992). Operationally defined apoplastic and symplastic aluminium fractions in root tips of aluminiumintoxicated wheat. Plant Physiol. 100:309-318. |
|
|
|
|
|
Raymet, G.E. and Probert, M.E.) (Kluwer Academic Publishers, Dordrecht, The Netherlands. pp. 5-19. |
|
|
|
|
|
von Uexku¨ll HR, Mutert E (1995). In: Plant-Soil Interactions at Low pH: Principles and Management [M] (eds. Date, R.A. Grundon, N.J. Waterland RA, Jirtle RL (2004). Early nutrition, epigenetic changes at transposons and imprinted genes, and enhanced susceptibility to adult chronic diseases. Nutrition 20:63-68. |
|
|
|
|
|
Wenzl P, Patino GM, Chaves AL, Mayer JE, Rao IM (2001). The high level of aluminum resistance in signalgrass is not associated with known mechanisms of external aluminum detoxification in root apices. Plant Physiol. 125:1473-84. |
|
|
|
|
|
Wood S, Sebastian K, Scherr SJ (2000). In: Pilot Analysis of Global Ecosystems: Agroecosystems [M] (ed. Rosen, C.) (International Food Policy Research Institute and the World Resources Institute, Washington, D.C. pp. 45-54. |
|
|
|
|
|
Xiong LZ, Xu CG, Shagi-Maroof MA, Zhang Q (1999). Patterns of cytosine methylation in an elite rice hybrid and its parental lines, detected by a methylation-sensitive amplification polymorphism technique. Mol. Genet. Genomics 261:439-446. |
|
|
|
|
|
Zhang C, Hsieh T (2013). Heritable Epigenetic Variation and its Potential Applications for Crop Improvement. Plant Breed. Biotechnol. 1(4):307-319. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


