Review

ABSTRACT
INTRODUCTION
The transmission network for central grid power lines of at least 220 kV covers about 300,000 km in Europe (European Network of Transmission System Operators for Electricity, 2012) and 250,000 km in the USA (Abraham, 2002). Power lines above 220 kV may only constitute a small proportion of the total grid. About 200,000 and 450,000 km of overhead power lines carrying various voltages exist in Norway (Statistics Norway, 2011a) and Sweden (Grusell and Miliander, 2004), respectively.
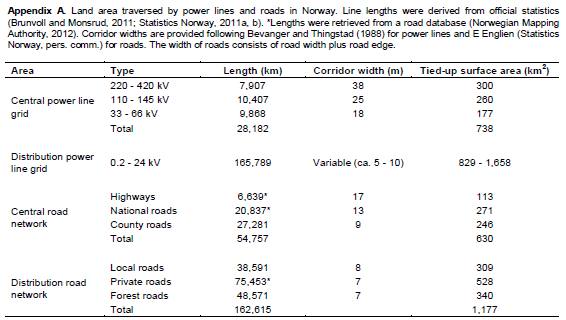
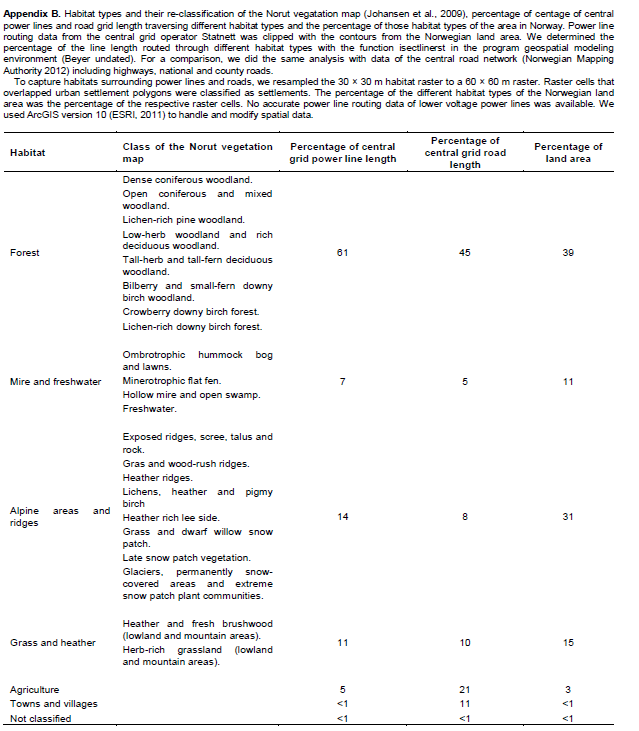
In Norway, the central power line grid covers over 20% more surface than the central road network due to required corridor widths (740 versus 630 km2), even though it is only half as long (28,000 versus 55,000 km) (Appendix A). The footprints of the distribution power line grid and road network are similar (Appendix A). We estimated that over 60% of central grid power lines traverse forests, while circa 40% of the Norwegian land area is covered by forest (Appendix B).The Norwegian central grid operator Statnett is planning to increase the construction of power lines to 300 km per year by 2020 (Statnett, 2013). 3,600 km of new power lines are planned in Germany until 2023 (German Transmission System Operators, 2013).
An extensive body of research on potential effects of roads on wildlife is available (reviewed in: Forman and Alexander (1998), Spellerberg (1998), Seiler (2001), Coffin (2007), Fahrig and Rytwinski (2009), Benitez-Lopez et al., (2010)), but knowledge on power line effects is scattered. Power lines may influence wildlife through disturbance, clearing of forest habitat under power lines, edge, barrier and corridor effects (Willyard and Tikalsky, 2004; Ball, 2012). Fragmentation by power lines could induce genetic drift, as for roads (Epps et al., 2005, Kuehn et al., 2007), and reduce population productivity and persistence (Griffen and Drake 2008; Haanes et al., 2013). Wide-ranging and mobile species as some ungulates will likely encounter power lines within their home ranges (Tables 1 to 7).
If disturbance by power lines is analogous to predation risk, it can cost energy for fleeing, increased vigilance, lost resources in habitats associated with danger and impaired mating and parental investment (Frid and Dill, 2002). If ungulates are disturbed by power lines, they can be expected to avoid power lines to reduce these costs. However, cleared areas under power lines (rights-of-way or ROW) are maintained as early- to mid-successional habitats (Bramble and Byrnes, 1982) and provide benefits through additional browse for forest ungulates (Bramble and Byrnes, 1972). This may result in attraction effects towards power line ROW.
The aim of this article is specifically to review the factors that possibly influence avoidance and attraction effects of power lines on ungulates (Tables 1 to 7), set into a conceptual framework (Figure 1). The review is based on both peer-reviewed and grey literature found through the Web of Science (isiknowledge.com) and Google Scholar, proceedings from the symposia on Environmental Concerns in Rights-of-Way Management (http://rights-of-way.org/1content.htm) and reviews on road ecology (see above). We searched for the keywords power line or transmission line in combination with ungulate, deer, elk, reindeer, sheep or cow; and subsequently scanned the papers for relevant references that did not show in the search engine. Research has mainly been undertaken on reindeer (Rangifer tarandus tarandus) in open alpine areas in Norway and on other ungulates in forests in Canada, USA and Norway (Table 1 to 7).
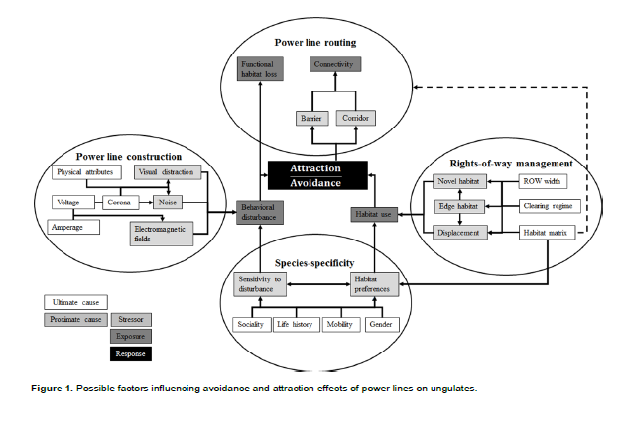
We identified proximate and ultimate causes of avoidance and attraction effects resulting from power line routing, construction, ROW management and species-specific effects (Figure 1). Power line construction may induce behavioural disturbance. ROW management and routing is expected to affect the use of ROW habitat. Sensitivity to disturbance and habitat preferences, which are species-specific, should further affect behavioural disturbance and habitat use. Both behavioural disturbance and habitat use determine avoidance and attraction effects. Therefore, power line routes could function as barriers and/or corridors, with consequences for connectivity and functional loss of habitats. Finally, we suggest how to mitigate avoidance effects through power line routing and ROW vegetation management.
EFFECTS OF POWER LINES ON UNGULATES
Behavioural disturbance from power line constructions
Power lines could disturb ungulates because they are artificial structures that can emit noise, light and electromagnetic fields. Frid and Dill (2002) reported that disturbance should be analogous to predation risk. Ungulates can be expected to alter their behaviour close to power lines if they are disturbed by power lines.
Noise
Electrical discharge by power lines produces crackling or hissing corona noise (Straumann, 2011). Wind can produce Aeolian noise though vibrations of the physical structure (Tsujimoto et al., 1991). An audiogram suggests that reindeer can hear corona noise from power lines (300 and 420 kV) up to 79 m (Flydal et al., 2010). Although little is known on the effects of corona noise, noise of a 500 kV transmission line did not significantly influence cattle behaviour (Ganskopp et al., 1991).
Electromagnetic fields
Power line electromagnetic fields are suspected to disturb the hypothesized magnetic alignment of cattle and roe deer (Capreolus capreolus) (Burda et al., 2009). However, ungulates may align themselves in the direction of power lines (Burda et al., 2009), supposedly interrupted in their north south alignment (Begall et al., 2008), for other reasons that were not accounted for such as wind and solar conditions for thermoregulation (Hetem et al., 2011). Fluctuations in electric fields of a 500 kV power line did not influence cattle behaviour (Ganskopp et al., 1991). Domestic-tame reindeer in enclosures became more restless and moved away from power lines (132 and 300 kV) when transmission load increased, although these results were ambiguous (Flydal et al., 2009).
Visual distraction
Visual distraction of power lines in the absence of noise and electromagnetic fields has apparently not been tested. Reindeer are suspected to see ultraviolet (UV) light (Hogg et al., 2011) and consequently corona flashes from power lines (Tyler et al., 2014). Reindeer’s sensitivity to UV light is suspected to aid in detecting predators and forage in arctic environments (Hogg et al., 2011). The strongest emission of UV light by power lines was centred on 337 nm wavelength according to a patent application for a corona detector (Le et al., 1994). The eye lens of ungulates however blocked the largest proportion of light at this wavelength (Douglas and Jeffery, 2014). This may explain the lack of rentinal response towards UV light of 325 nm wavelength by other ungulates (Jacobs et al., 1994).
The lack of behavioural disturbance under power lines in general may indicate that the sight of power lines was not a source of disturbance. Deer (Odocoileus spp.), elk (Cervus canandensis) and other ungulates fed in a power line ROW (500 kV, 41 m wide) without signs of disturbance apart from a five-minute motionless period when entering the ROW (Goodwin Jr, 1975). Also semi-domestic reindeer in an enclosure experiment did not clearly avoid power lines (132 and 300 kV) (Flydal et al., 2009).
White-tailed deer (Odocoileus virginianus) (Bramble and Byrnes, 1972; Doucet et al., 1979; Doucet et al., 1983), moose (Alces alces) (unpublished data), bighorn sheep (Ovis canadensis canadensis) and elk (Goodwin Jr, 1975) bedded under power lines. Feeding positions and activity of cattle were similar in pens with and without power lines (Ganskopp et al., 1991). Other studies indicate that the vegetation in power line ROW rather than disturbance by power lines influences the use of those areas by ungulates (Goodwin Jr, 1975; Morhardt et al., 1984). Energy spent in response to indifferent stimuli may be wasted (Reimers and Colman, 2009).
Although the given examples provide little evidence for the disturbance of ungulate behaviour by power lines, it does not necessarily mean that ungulates are not impacted by power lines. Human disturbance increased cardiac rates of bighorn sheep without changing their behaviour (MacArthur et al., 1979, 1982). Chronic stress can have adverse effects on reproductive, immune and neural systems and suppress growth in the absence of behavioural changes (Wingfield et al., 1997).
Altered habitat use at power line rights-of-way
Displacement from rights-of-way
Besides possible disturbance effects of power lines, forest ungulates may be displaced from cleared habitats under power lines because they lack canopy cover (Rieucau et al., 2007) and forage in the first years after clearing (Bramble and Byrnes, 1982; Lamothe and Dupuy, 1984; Garant and Doucet, 1995; Ricard and Doucet, 1999; Hydro-Québec, 2013) (Table 2).
Moose tracks and white-tailed deer tracks and pellets were less abundant in power line ROW (220 and 735 kV; 90 to 140 m wide) as compared to forests at 2 km distance (Joyal et al., 1984) and forests adjacent to ROW (120 - 735 kV, 30 - 150 m wide), respectively (Doucet et al., 1979; Lamothe and Dupuy, 1984; Jackson and Hecklau, 1995). White-tailed deer abandoned significantly more food provided in feeders in a power line ROW (30 m wide) as compared to adjacent forest when regeneration was absent (Rieucau et al., 2007). White-tailed deer browsed a smaller proportion of stems despite higher availability in power line ROW as compared to adjacent forest, except where the abundance of browsed stems exceeded that of the forest approximately six-fold (Mayer, 1976).
These results indicate that forest ungulates may be displaced by power line ROW (Table 2), especially when food, cover or both are lacking (Joyal et al., 1984). Increased food abundance may however compensate for the lack of cover (Mayer, 1976; Rieucau et al., 2007).
Rights-of-way as novel habitat
Following regrowth, habitats in power line ROW can also create novel habitats for forest ungulates through the provision of attractive feeding opportunities (Bramble and Byrnes, 1979; Ricard and Doucet, 1999; Hydro-Québec, 2013) (Table 3). White-tailed deer deposited more pellet groups, foraged more intensely and left more signs in power line ROW as compared to forests adjacent to ROW or control forest (Bramble and Byrnes, 1972; Cavanagh et al., 1976). The ROW provided more stems for browsing. Black-tailed deer (Odocoileus hemionus columbianus) used a power line ROW significantly more than adjacent mature forest, indicated by pellet groups (Loft and Menke, 1984). Deer use increased with shrub and herbaceous cover as well as foraging plants. These results indicate that food availability in power line ROW habitat is important for the use of that habitat by forest ungulates.
Not only the amount of forage, but also its composition may influence the use of power line ROW for browsing (Milligan and Koricheva, 2013). Moose and white-tailed deer browsing intensity in power line ROW appeared to be influenced by the proportion of preferred browse species rather than browse availability (Garant et al., 1987; Ricard and Doucet, 1999).
Trees that have been cut in power line ROW could provide higher quality browse because they prioritize growth instead of defence against herbivore damage through secondary metabolites (Rea and Gillingham, 2001). However, the increased availability of light in power line ROW clearings may promote both growth and defence (Nybakken et al., 2013). Herbs in a power line ROW provided higher concentrations of protein and minerals and contained less fiber as compared to woody browse (Bramble and Byrnes, 1972). Forbs in power line ROW contained more protein and minerals as compared to grasses and woody browse (Harlow et al., 1995). The quality of the forage can be expected to influence the attractiveness of power line ROW habitat for forest ungulates.
.png)
Edge habitat along rights-of-way
Forest ungulates can benefit from the increased availability of forage not only inside power line ROW but also along edges (Bramble and Byrnes, 1979). Stem availability within 3-10 m from power line ROW edges was elevated as compared to forests at further distances from the edge (Luken et al., 1991; Luken et al., 1992; Rieucau et al., 2007; Powell and Lindquist, 2011). 18 of 20 shrub species were significantly more likely to be found at the edge of a 60 m wide power line ROW as compared to its centre (Brisson et al., 1997).
Lamothe and Dupuy (1984) noted more white-tailed deer tracks along the edge of a power line ROW (twin power lines, 735 kV, 150 m wide) as compared to the ROW and lateral forest (Table 4). However, fewer pellets were found in the ecotones between cleared areas and lateral forests as compared to further inside the forests (Lamothe and Dupuy, 1984; Jackson and Hecklau, 1995), indicating that deer may have spent more time in areas of better cover. The abundance of stems along power line ROW edges as compared to ROW and adjacent woods and the proportion of those stems that were browsed by white-tailed deer did not follow a consistent pattern across study areas (Mayer, 1976). Shrub availability along power line ROW edges may favour ungulate browsing and habitat use along those edges but a link between the two has, as far as we know, not been established.
Functional loss of habitat
Disturbance by power lines may not only affect the use of areas directly under power lines but also habitats adjacent to it. Power lines contributed to a reduction in area use of wild female reindeer within 1 km from pitfall traps and hunting blinds (Panzacchi et al., 2013). The density of semi-domesticated reindeer was significantly (73%) lower within 4 km of a power line (132 kV) than further away during calving in areas of rugged terrain (Vistnes and Nellemann, 2001) (Table 5). However, more favourable snow conditions and lower predation rates at higher elevations further away from power lines may have influenced this result (Reimers and Colman, 2009).
Wild reindeer were significantly less abundant than expected within 2.5 km of power lines (300 and 420 kV) in six of eight sampling years (Nellemann et al., 2001) (Table 5). Areas transected by power lines (66 - 420 kV) were also used less than expected (Vistnes et al., 2001). However, the accessibility of lichen forage, provided by an index of snow depth and hardness, was approximately three times lower in areas transected by power lines and other infrastructure (Vistnes et al., 2001). The influence of forage accessibility, although not significantly different between areas, can be discussed.
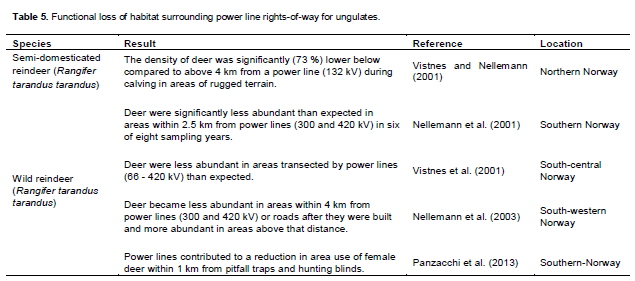
Wild reindeer became less abundant within 4 km from power lines (300 and 420 kV) or roads after they were built and more abundant beyond this distance (Nellemann et al., 2003). However, the shift in abundance coincides with the flooding of an area close to power lines and roads following the construction of a dam (Nellemann et al., 2003).In contrast, counts of wild reindeer were dispro-portionately high within 5 km of power lines and minor roads above 1,400 m a.s.l. in summer (Vistnes et al., 2008). There was no clear evidence for aversion by wild reindeer along a 66 kV power line indicated by lichen measurements (Reimers et al., 2007). Moose did not avoid moving towards central grid power lines except in certain habitats during autumn (Bartzke et al., in press). Hydro-Québec (2013) reported that the use of winter feeding grounds by white-tailed deer was not inhibited by power line ROW (120 - 735 kV, 30 - 164 m wide) close to them.
Connectivity and power line routing
Power line routes as barriers
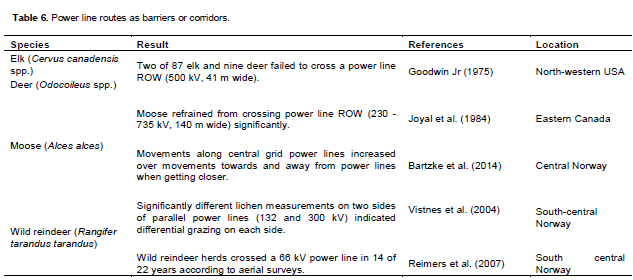
Disturbance by power lines and the lack of canopy cover in power line ROW may prevent animals from crossing power lines. Vistnes et al. (2004) concluded that wild reindeer refrained from crossing power lines based on lichen measurements on two sides of parallel power lines (132 and 300 kV) indicating differential grazing. However, these power lines were routed along a dam in the northern part of the study area that could have impeded crossings. The side that was apparently less grazed in the southern part of the study area was closer to a main road and urban settlements at lower elevations. Reimers et al. (2007) suggested that harvesting along a summer open road close to power lines could have been another alternative explanation for reduced area use.
Wild reindeer crossed a 66 kV power line in 14 of 22 years according to aerial surveys (Reimers et al., 2007). Strand et al. (2001) hypothesized that wild reindeer cross barriers when the need to migrate is extra-large based on the difference in the availability and accessibility of forage, disturbances, predation risk and distance between alternative feeding areas. Moose did not refrain from crossing power lines (735 kV) with ROW that were 90 m wide (Joyal et al., 1984). Neither did white-tailed deer refrain from crossing power line (120 - 735 kV) ROW of 30 - 146 m width (Hydro-Québec, 2013). Only two animals of 87 elk and nine deer (Odocoileus spp.) failed to cross a power line ROW (500 kV, 41 m wide) (Goodwin Jr, 1975) (Table 6).
However, white-tailed deer reduced crossings away from a planted area within a power line ROW (120 kV, 30 m wide) (Doucet et al., 1983). Moose refrained from crossing power line (230 - 735 kV) ROW that were 140 m wide (Joyal et al., 1984). The need to cross power lines, the size of the power line construction, transmission load, the width of the corridor and the availability of cover may influence the willingness of ungulates to cross power lines.
Power line routes as corridors
Species-specific ultimate and proximate causes
Sociality
Reindeer in Norway may in general be more sensitive to power lines than other ungulates like moose or white-tailed deer because they live in large groups in open alpine habitat experiencing comparatively low human use but intense seasonal hunting. These attributes were shown to increase ungulate flight responses (Stankowich, 2008). In open habitats, ungulates should detect and react to disturbances at greater distances because there are no escape habitats to seek refuge (Stankowich, 2008).
The ability to quickly detect and react to disturbances could be an evolutionary advantage to protect against real predators but a disadvantage if the source of the disturbance is not lethal. Then animals loose time and energy in being unnecessarily disturbed. The degree of reindeer domestication may also influence their sensitivity to disturbances (Flydal et al., 2009; Reimers et al., 2012).
Mobility
The lack of power line avoidance by forest ungulates may also in part be explained by the mobility of the species of concern. Stationary species and species with small home ranges may not have alternative habitats available, or the costs of reaching those habitats outweigh the costs of remaining close to power lines (Gill et al., 2001). This could occur in times or areas of resource limitations. White-tailed deer increased stationary browsing time in a power line ROW from 7 to ~40% in cold winter as compared to a mild winter (Doucet et al., 1987). Possibly forest ungulates cannot afford to avoid power line ROW when overall food availability is insufficient in relation to their densities (Ytrehus et al., 1999; Lamoureux et al., 2001). Hagen et al. (2007) speculated that reindeer will also react less to disturbances when the population size increases.
Gender and life history
Throughout a species’ life cycle, its disturbance tolerance towards power lines may vary with life history traits and states such as gender, age, reproductive status, social status and/or season (for example, rut) (Frid and Dill, 2002; Stankowich, 2008).Several, although not all (Frid, 2003; Mahoney et al., 2011), studies indicate that groups with calves (Aastrup, 2000), females with young (Ciuti et al., 2008) and females in times of calving or raising calves (Maier et al., 1998; Vistnes and Nellemann 2001; Bartzke et al., in press) are most sensitive to human disturbance (Wolfe et al., 2000). However, no clear evidence for gender-specific effects towards power lines was found (Vistnes and Nellemann, 2001; Bartzke et al., in press). Possibly, power lines are not disturbing enough to promote gender-specific effects in contrast do direct harassment by humans or motorized vehicles.
Potential demographic impacts
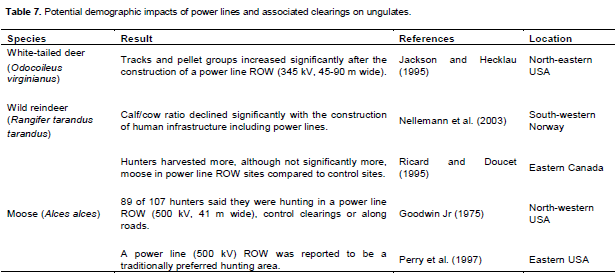
Fragmentation effects by power lines have been argued to contribute to population decline (Nellemann et al., 2003), impair migration (Vistnes et al., 2004) and could induce genetic drift, as for roads (Epps et al., 2005; Kuehn et al., 2007). Yet we found no evidence that the construction of power lines alone influenced population dynamics of ungulates. Tracks and pellet groups of white-tailed deer increased significantly after the construction of a power line (345 kV, 45-90 m wide) (Jackson and Hecklau, 1995) (Table 7).
Although power line ROW can provide additional forage, they could also be an “ecological trap” (Battin, 2004). Ecological traps are thought to occur when the attractiveness of a habitat (through increased browse) increases disproportionately relative to its value for survival and reproduction. An increased rate of disturbance in connection with an increased rate of predator encounters can result in a reduction of population size (Frid and Dill, 2002) due to increased antipredator investment (stress) (Ydenberg and Dill, 1986; Cassirer et al., 1992; Maier et al., 1998; Rumble et al., 2005), reduced net energy intake (Stockwell et al., 1991) and body condition (Bradshaw et al., 1998; Luick et al., 2011). Power line ROW along with access roads may provide access for hunters (Goodwin Jr, 1975; Ricard and Doucet, 1995; Perry et al., 1997). Natural predators were reported to travel along power line ROW (Foster, 1956 in Ball, 2012; Paquet et al., 1996; Gurarie et al., 2011). Power lines provide nesting and perching opportunities for golden eagle (Aquila chrysaetos) (Steenhof et al., 1993; Prather and Messmer 2010), which preys on ungulates (Nybakk et al., 2002; Norberg et al., 2006; Johnsen et al., 2007; Hamel and Côté, 2009; Nadjafzadeh et al., 2013).
Increased predation risk, coupled with disturbance and fragmentation induced by power lines, could affect the demography of ungulate populations. The availability of forage in power line ROW may however favour forest ungulate populations locally.
KNOWLEDGE GAPS
MITIGATION MEASURES
Implications for power line routing
Implications for power line ROW management
Disclosure or conflict of interest
The Norwegian central power line grid operator Statnett and Energy Norway, a non-profit industry organization representing companies involved in the production, distribution and trading of electricity, contributed to the funding of this study. Neither Statnett nor Energy Norway could determine or change the contents of this review.
ACKNOWLEDGEMENTS
We acknowledge our funding and cooperating partners in the Centre for Environmental Design of Renewable Energy (CEDREN): the Research Council of Norway, the Norwegian Water Resources and Energy Directorate, the Norwegian Directorate for Nature Management, Statnett and Energy Norway. We thank G. J. Doucet for providing literature and P. A. Aarestad for his help with translations.
REFERENCES
|
Aastrup P (2000). Responses of West Greenland caribou to the approach of humans on foot. Polar Res. 19:83-90. Crossref |
||||
| Abraham S (2002). National transmission grid study. Report from U.S. Department of Energy, Washington, USA. 93 pp. | ||||
|
Andreassen HP, Gundersen H, Storaas T (2005). The effect of scent-marking, forest clearing, and supplemental feeding on moose-train collisions. J. Wildl. Manage. 69:1125-1132. Crossref |
||||
| Ball SK (2012). Capitalizing on Conservation: The Ecological Benefits of Transmission Line Rights-of-Way. Pages 249-272 in Evans JM, Mahoney JWG, Mutrie D, Reinemann J, editors. Proc. Ninth Int. Symp. Environmental Concerns Rights-of-Way Manage., Portland, Oregon, USA. International Society for Arboriculture, Champaign, Illinois, USA. | ||||
| Ballard BD, Nowak CA, Abrahamson LP, Neuhauser EF, Finch KE (2002). Integrated Vegetation Management on Electrical Transmission Rights-of-Way using Herbicides: Treatment Effects Over Time. Pages 47-55 in Goodrich-Mahoney JW, Mutrie D, Guild C, editors. Proc. Seventh Int. Symp. on Environ. Concerns in Rights-of-Way Manage., Calgary, Alberta, Canada. Elsevier Science Ltd., Amsterdam, Netherlands. | ||||
| Bartzke GS, May R, Stokke S, Røskaft E, Engen S (in press). Comparative effects of power lines and roads on moose (Alces alces) habitat selection. in Proc. Tenth Int. Symp. on Environ. Concerns in Rights-of-Way Manage., Phoenix, Arizona, USA. | ||||
| Bartzke G, May R, Røskaft E (2014). Differential barrier and corridor effects of power lines, roads and rivers on moose (Alces alces) movements. in Bartzke GS. Effects of power lines on moose (Alces alces) habitat selection, movements and feeding activity. PhD thesis. NTNU, Trondheim, Sør-Trøndelag, Norway. | ||||
|
Battin J (2004). When Good Animals Love Bad Habitats: Ecological Traps and the Conservation of Animal Populations. Conserv. Biol. 18:1482-1491. Crossref |
||||
|
Begall S, ÄŒervený J, Neef J, VojtÄ›ch O, Burda H (2008). Magnetic alignment in grazing and resting cattle and deer. Proc. Natl. Acad. Sci. USA. 105: 13451-13455. Crossref |
||||
|
Benitez-Lopez A, Alkemade R, Verweij PA (2010). The impacts of roads and other infrastructure on mammal and bird populations: A meta-analysis. Biol. Conserv. 143: 1307-1316. Crossref |
||||
| Bergerud AT, Jakimchuk RD, Carruthers DR (1984). The Buffalo of the North: Caribou (Rangifer tarandus) and Human Developments. Arctic. 73: 7-22. | ||||
| Bevanger K, Thingstad PG (1988). Forholdet fugl - konstruksjoner for overføring av elektrisk energi. En oversikt over kunnskapsnivået. Report 1988:1 from Økoforskutredning, Trondheim, Norway.pp.133. | ||||
| Beyer HL (undated). Geospatial Modelling Environment. | ||||
|
Beyers DW (1998). Causal inference in environmental impact studies. J. N. Am. Benthol. Soc. 17: 367-373. Crossref |
||||
|
Bjørneraas K, Solberg EJ, Herfindal I, Moorter BV, Rolandsen CM, Tremblay JP, Skarpe C, Sæther B-E, Eriksen R, Astrup R (2011). Moose Alces alces habitat use at multiple temporal scales in a human-altered landscape. Wildl. Biol. 17: 44-54. Crossref |
||||
|
Bjornlie DD, Garrott RA (2001). Effects of Winter Road Grooming on Bison in Yellowstone National Park. J. Wildl. Manage. 65: 560-572. Crossref |
||||
|
Bradshaw CJA, Boutin S, Hebert DM (1998). Energetic implications of disturbance caused by petroleum exploration to woodland caribou. Can. J. Zool.-Rev. Can. Zool. 76: 1319-1324. Crossref |
||||
| Bramble WC, Byrnes WR (1972). A Long-term Ecological Study of Game Food and Cover on a Sprayed Utility Right-of-Way. Report 885 from Purdue University, West Lafayette, Indiana, USA. 20 pp. | ||||
|
Bramble WC, Byrnes WR (1979). Evaluation of the Wildlife Habitat Values of Rights-of-Way. J. Wildl. Manage. 43: 642-649. Crossref |
||||
| Bramble WC, Byrnes WR (1982). Development of Wildlife Food and Cover on an Electric Transmission Right-of-Way Maintained by Herbicides: A 30-Year Report. Report from Purdue University, West Lafayette, Indiana, USA. 24 pp. | ||||
| Brisson J, Meilleur A, Fortin M-J, Bouchard A (1997). Edge Effects on Vegetation in Rights-of-Way. Pages 25-33 in James RW, Goodrich-Mahoney JW, Wisniewski JR, Wisniewski J, editors. Proc. Sixth Int. Symp. Environ. Concerns Rights-of-Way Manage., New Orleans, Louisiana, USA. Elsevier Science Ltd., Oxford, UK; New York, USA; Tokyo, Japan. | ||||
|
Bruggeman JE, Garrott RA, Bjornlie DD, White PJ, Watson FGR, Borkowski J (2006). Temporal variability in winter travel patterns of Yellowstone bison: The effects of road grooming. Ecol. Appl. 16:1539-1554. Crossref |
||||
|
Bruggeman JE, Garrott RA, White PJ, Watson FGR, Wallen R (2007). Covariates Affecting Spatial Variability in Bison Travel Behavior in Yellowstone National Park. Ecol. Appl. 17: 1411-1423. Crossref |
||||
| Brunvoll F, Monsrud J (2011). Samferdsel og miljø 2011: Utvalgte indikatorer for samferdselssektoren. Report 21/2011 from Statistics Norway, Oslo, Norway. 163 pp. | ||||
|
Burda H, Begall S, ÄŒervený J, Neefa J, NÄ›mec P (2009). Extremely low-frequency electromagnetic fields disrupt magnetic alignment of ruminants. Proc. Natl. Acad. Sci. USA. 106: 5708-5713. Crossref |
||||
| Cassirer EF, Freddy DJ, Ables ED (1992). Elk Responses to Disturbance by Cross-Country Skiers in Yellowstone National Park. Wildl. Soc. Bull. 20: 375-381. | ||||
| Cavanagh JB, Olson DP, Macrigeanis SN (1976). Wildlife Use and Management of Powerline Rights-of-Way in New Hampshire. Pages 276-294 in Tillman R, editor. Proc. First Nat. Symp. Environ. Concerns Rights-of-Way Manage., Starkville, Mississippi, USA. Mississippi State University, Starkville, Mississippi, USA. | ||||
|
Ciuti S, Pipia A, Ghiandai F, Grignolio S, Apollonio M (2008). The key role of lamb presence in affecting flight response in Sardinian mouflon (Ovis orientalis musimon). Behav. Processes. 77: 408-412. Crossref |
||||
|
Clair CCS, Forrest A. (2009). Impacts of vehicle traffic on the distribution and behaviour of rutting elk, Cervus elaphus. Behav. 146: 393-413. Crossref |
||||
|
Coffin AW (2007). From roadkill to road ecology: A review of the ecological effects of roads. J. Transp. Geogr. 15: 396-406. Crossref |
||||
| Colman JE, Jacobsen BW, Reimers E (2001). Summer response distances of Svalbard reindeer Rangifer tarandus platyrhynchus to provocation by humans on foot. Wildl. Biol. 7: 275-284. | ||||
|
Conti R, Giorgi A, Rendina R, Sartore L, Sena EA (2003). Technical Solutions To Reduce 50 Hz Magnetic Fields from Power Lines. in IEEE Bologna Power Tech Conf. Proc., Bologna, Italy. Crossref |
||||
| Doucet GJ, Brown DD, Lamothe P (1983). White-tailed deer response to conifer plantation as a mitigation measure in a power line right-of-way located in Quebec deer yard. Pages 150-156 in Yahner RH, editor. Trans. Northeast Sect. Wildl. Soc., West Dover, Vermont, USA. Northeast Section, the Wildlife Society, West Dover, Vermont, USA. | ||||
| Doucet GJ, Brown DT, Lamothe P (1987). Deer Behaviour in a Powerline Right-of-Way Located in a Northern Wintering Yard. Pages 7-12 in Byrnes WR, Holt HA, editors. Proc. Fourth Symp. Environ. Concerns Rights-of-Way Manag., Indianapolis, Indiana, USA. Purdue University, West Lafayette, Indiana, USA. | ||||
| Doucet GJ, Stewart RW, Morrison KA (1979). The Effect of a Utility Right-of-Way on White-Tailed Deer in a Northern Deer Yard. in Tillman R, editor. Proc. Second Nat. Symp. Environ. Concerns Rights-of-Way Manag., Ann Arbor, Michigan, USA. Mississippi State University, Starkville, Mississippi, USA. | ||||
|
Douglas RH, Jeffery G (2014). The spectral transmission of ocular media suggests ultraviolet sensitivity is widespread among mammals. Proc. R. Soc. B. 281: 20132995. Crossref |
||||
|
Dunne BM, Quinn MS (2009). Effectiveness of above-ground pipeline mitigation for moose (Alces alces) and other large mammals. Biol. Conserv. 142: 332-343. Crossref |
||||
|
Epps CW, Palsboll PJ, Wehausen JD, Roderick GK, Ramey RR, McCullough DR (2005). Highways block gene flow and cause a rapid decline in genetic diversity of desert bighorn sheep. Ecol. Lett. 8: 1029-1038. Crossref |
||||
| ESRI (2011). ArcGIS Desktop Version 10.1. Redlands, California, USA. | ||||
| European Network of Transmission System Operators for Electricity (2012). Excel attachment "YS & AR 2012 Table 1 operational data 2012" of: Yearly Statistics & Adequacy Retrospect 2012. Report from ENTSOE, Brussels, Belgium. 62 pp. | ||||
| Fahrig L, Rytwinski T (2009). Effects of roads on animal abundance: an empirical review and synthesis. Ecol. Soc. 14: 21. | ||||
|
Flydal K, Kilde IR, Enger PS, Reimers E (2010). Reindeer (Rangifer tarandus tarandus) perception of noise from power lines. Rangifer. 23:21-24. Crossref |
||||
|
Flydal K, Korslund L, Reimers E, Johansen F, Colman J (2009). Effects of Power Lines on Area Use and Behaviour of Semi-Domestic Reindeer in Enclosures. Int. J. Ecol. 1-14. Crossref |
||||
|
Forman RTT, Alexander LE (1998). Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 29:207-231. Crossref |
||||
|
Forman RTT, Deblinger RD (2000). The Ecological Road-effect Zone of a Massachusetts (USA) Suburban Highway. Conserv. Biol. 14: 36-46. Crossref |
||||
| Foster CHW (1956). Wildlife use of utility rights-of-way in Michigan. Master thesis. University of Michigan, Michigan, USA. | ||||
|
Frid A (2003). Dall's sheep responses to overflights by helicopter and fixed-wing aircraft. Biol. Conserv. 110:387-399. Crossref |
||||
| Frid A, Dill L (2002). Human-caused Disturbance Stimuli as a Form of Predation Risk. Conserv. Ecol. 6:11. | ||||
|
Ganskopp D, Raleigh R, Schott M, Bracken TD (1991). Behavior of cattle in pens exposed to +/-500-kV DC transmission-lines. Appl. Anim. Behav. Sci. 30:1-16. Crossref |
||||
| Garant Y, Doucet GJ (1995). An experimental winter cut in a powerline ROW located in a white-tailed deer yard Pages 266-268 in Doucet GJ, Séguin C, Giguère M, editors. Proc. Fifth Int. Symp. Environ. Concerns Rights-of-Way Manage., Montréal, Québec, Canada. Vice Présidence Environnement Hydro-Québec, Montréal, Québec, Canada. | ||||
| Garant YB, Doucet GJ, Hayeur G (1987). Winter Deer Browse Production and Use in a Powerline Right-of-Way Six Years After Initial Clearing. Pages 56-63 in Byrnes WR, Holt HA, editors. Proc. Fourth Symp. Environ. Concerns Rights-of-Way Manag., Indianapolis, Indiana, USA. Purdue University, West Lafayette, Indiana, USA. | ||||
| German transmission system operators (2013). Conclusion of the GDP 2013, second draft. | ||||
|
Gill JA, Norris K, Sutherland WJ (2001). Why behavioural responses may not reflect the population consequences of human disturbance. Biol. Conserv. 97:265-268. Crossref |
||||
| Global Wildlife Management LLC (2013). Hinge Cutting. | ||||
| Goodwin Jr JG (1975). Big Game Movement Near a 500-kV Transmission Line in Northern Idaho. Report from Western Interstate Commission for Higher Education (WICHE), Resource Development Internship Program (RDIP), Boulder, Colorado, USA. 56 pp. | ||||
|
Griffen BD, Drake JM (2008). A review of extinction in experimental populations. J. Anim. Ecol. 77:1274-1287. Crossref |
||||
| Grusell E, Miliander S (2004). GIS-baserad identifiering av artrika kraftledningsgator inom stamnätet. Report 1960900 from Svenska Kraftnät. 19 pp. | ||||
|
Gurarie E, Suutarinen J, Kojola I, Ovaskainen O (2011). Summer movements, predation and habitat use of wolves in human modified boreal forests. Oecologia. 165: 891-903. Crossref |
||||
|
Haanes H, Markussen SS, Herfindal I, Røed KH, Solberg EJ, Heim M, Midthjell L, Sæther BE (2013). Effects of inbreeding on fitness-related traits in a small isolated moose population. Ecol. Evol. 3: 4230-4242. Crossref |
||||
| Hagen D, Bevanger K, Hanssen F, Thomassen J (2007). The dialog project "Mutual politics in the mountain areas". Baseline knowledge on infrastructure development, land use and disturbance in the reindeer herding areas in Selbu, Tydal, Røros and Holtålen municipalities. Report 225 from NINA, Trondheim, Norway. 77 pp. | ||||
| Haggie MR, Johnstone RA, Allen HA (2008). Tree, Shrub, and Herb Succession and Five Years of Management Following the Establishment of a New Electric Transmission Right-of-Way through a Wooded Wetland. Pages 47-59 in Goodrich-Mahoney JW, Abrahamson LP, Ballard JL, editors. Proc. Eighth Int. Symp. Envir. Concerns Rights-of-Way Manage., Saratoga Springs, New York, USA. Elsevier, Amsterdam, Netherlands. | ||||
|
Hamel S, Côté SD (2009). Maternal Defensive Behavior of Mountain Goats Against Predation by Golden Eagles. West. N. Am. Nat. 69:115-118. Crossref |
||||
| Harlow RF, David C. Guynn J, Davis JR (1995). The Effect of Management Treatments on the Biomass, Nutritive Quality, and Utilization of Deer Forages on Utility Rights-of-Way. Pages 284-289 in Doucet GJ, Séguin C, Giguère M, editors. Proc. Fifth Int. Symp. Environ. Concerns Rights-of-Way Manage., Montréal, Québec, Canada. Vice Présidence Environnement Hydro-Québec, Montréal, Québec Canada. | ||||
|
Hetem RS, Strauss WM, Heusinkveld BG, de Bie S, Prins HHT, van Wieren SE (2011). Energy advantages of orientation to solar radiation in three African ruminants. J. Therm. Biol. 36:452-460. Crossref |
||||
| Hjeljord O, Hövik N, Pedersen HB (1990). Choice of feeding sites by moose during summer, the influence of forest structure and plant phenology. Holarct Ecol. 13:281-292. | ||||
|
Hogg C, Neveu M, Stokkan KA, Folkow L, Cottrill P, Douglas R, Hunt DM, Jeffery G (2011). Arctic reindeer extend their visual range into the ultraviolet. J. Exp. Biol. 214:2014-2019. Crossref |
||||
| Hydro-Québec (2013). Ravages du cerf de Virginie - Synthèse des connaissances environnementales pour les lignes et les postes. Report 2013E0789-17 from Hydro-Québec, Montréal, Québec, Canada. 19 pp. | ||||
| Jackson W, Hecklau J (1995). Construction effects of a 345 kV electric corridor on New York deer. Pages 290-299 in Doucet GJ, Séguin C, Giguère M, editors. Proc. Fifth Int. Symp. Environ. Concerns Rights-of-Way Manage., Montréal, Québec, Canada. Vice Présidence Environnement Hydro-Québec, Montréal, Québec, Canada. | ||||
|
Jacobs GH, Deegan JF, Neitz J, Murphy BP, Miller KV, Marchinton RL (1994). Electrophysiological measurements of spectral mechanisms in the retinas of 2 cervids - white-tailed deer (Odocoileus virginianus) and fallow deer (Dama-Dama). J. Comp. Physiol. A Sens Neural Behav. Physiol. 174:551-557. Crossref |
||||
| Jaeger JAG, Fahrig L, Ewald KC (2005). Does the configuration of road networks influence the degree to which roads affect wildlife populations? Pages 151-163 in Irwin C, Garrett P, McDermott K, editors. Proc. 2005 Int. Conf. Ecol. Transp., San Diego, California, USA. Center for Transportation and the Environment, North Carolina State University, Raleigh, North Carolina, USA. | ||||
| Johansen B, Aarrestad PA, Øien DI (2009). Vegetasjonskart for Norge basert på satellittdata; Delprosjekt 1: Klasseinndeling og beskrivelse av utskilte vegetasjonstyper. Report 387 from NORUT/NINA/NTNU, Trondheim, Norway. 34 pp. | ||||
| Johnsen TV, Systad GH, Jacobsen KO, Nygard T, Bustnes JO (2007). The occurrence of reindeer calves in the diet of nesting Golden Eagles in Finnmark, northern Norway. Ornis Fenn. 84:112-118. | ||||
|
Joyal R, Lamothe P, Fournier R (1984). L'utilisation des emprises de lignes de transport d'énergie électrique par l'orignal (Alces alces) en hiver. Can. J. Zool.-Rev. Can. Zool. 62:260-266. Crossref |
||||
|
Kalén C, Bergquist J (2004). Forage availability for moose of young silver birch and Scots pine. For. Ecol. Manage. 187: 149-158. Crossref |
||||
|
Kalhor HA, Zunoubi MR (2005). Mitigation of Power Frequency Fields by Proper Choice of Line Configuration and Shielding. Electromagnetics. 25:231-243. Crossref |
||||
|
Krojerová-Prokešováa J, BaranÄekováa M, Šustrb P, Heurich M (2010). Feeding patterns of red deer Cervus elaphus along an altitudinal gradient in the Bohemian Forest: effect of habitat and season. Wildl. Biol. 16: 173-184. Crossref |
||||
|
Kuehn R, Hindenlang KE, Holzgang O, Senn J, Stoeckle B, Sperisen C (2007). Genetic Effect of Transportation Infrastructure on Roe Deer Populations (Capreolus capreolus). J. Hered. 98:13-22. Crossref |
||||
|
Kuijper DPJ, Cromsigt JPGM, Churski M, Adam B, Jędrzejewska B, Jędrzejewski W (2009). Do ungulates preferentially feed in forest gaps in European temperate forest? For. Ecol. Manage. 258: 1528-1535. Crossref |
||||
| Lamothe P, Dupuy P (1984). Special Consideration for Implanting Two 735 kV Lines in the Hill Head Deer Yard; Near Montreal. Pages 581-591 in Crabtree AF, editor. Proc. Third Int. Symp. Environ. Concerns Rights-of-Way Manage., San Diego, California, USA. Mississippi State University, Starkville, Mississippi, USA. | ||||
| Lamoureux J, Crête M, Bélanger M (2001). Effects of Reopening Hunting on Survival of White-tailed Deer, Odocoileus virginianus, in the Bas-Saint-Laurent region, Québec. Can. Field-Nat. 115:99-105. | ||||
| Le QT, Murray WH, Kinsella NS (1994). Corona detector with narrow-band optical filter. | ||||
|
Loft ER, Menke JW (1984). Deer Use and Habitat Characteristics of Transmission-Line Corridors in a Douglas-Fir Forest. J. Wildl. Manage. 48:1311-1316. Crossref |
||||
|
Luick BR, Kitchens JA, White RG, Murphy SM (2011). Modeling energy and reproductive costs in caribou exposed to low flying military jet aircraft. Rangifer. 16:209-212. Crossref |
||||
|
Luken JO, Hinton AC, Baker DG (1991). Forest edges associated with power-line corridors and implications for corridor siting. Landscape Urban Plann. 20:315-324. Crossref |
||||
|
Luken JO, Hinton AC, Baker DG (1992). Response of woody plant communities in power-line corridors to frequent anthropogenic disturbance. Ecol. Appl. 2:356-362. Crossref |
||||
|
MacArthur RA, Geist V, Johnston RH (1982). Cardiac and Behavioral Responses of Mountain Sheep to Human Disturbance. J. Wildl. Manage. 46:351-358. Crossref |
||||
|
MacArthur RA, Johnston RH, Geist V (1979). Factors influencing heart rate in free-ranging bighorn sheep: a physiological approach to the study of wildlife harassment. Can. J. Zool.-Rev. Can. Zool. 57:2010-2021. Crossref |
||||
|
Mahoney SP, Mawhinney K, McCarthy C, Anions D, Taylor S (2011). Caribou reactions to provocation by snowmachines in Newfoundland. Rangifer. 21:35-43. Crossref |
||||
|
Maier JAK, Murphy SM, White RG, Smith MD (1998). Responses of Caribou to Overflights by Low-Altitude Jet Aircraft. J. Wildl. Manage. 62: 752-766. Crossref |
||||
| Mayer TD (1976). An Evaluation of Chemically-Sprayed Electric Transmission Line Rights-of-Way For Actual and Potential Wildlife Use. Pages 288-294 in Tillman R, editor. Proc. First Nat. Symp. Environ. Concerns Rights-of-Way Manage. Starkville, Mississippi, USA. Mississippi State University, Starkville, Mississippi, USA. | ||||
|
Miller SG, Knight RL, Miller CK (2001). Wildlife responses to pedestrians and dogs. Wildl. Soc. Bull. 29:124-132. Crossref |
||||
|
Milligan HT, Koricheva J (2013). Effects of tree species richness and composition on moose winter browsing damage and foraging selectivity: an experimental study. J. Anim. Ecol. 82: 739-748. Crossref |
||||
|
Montgomery RA, Roloff GJ, Millspaugh JJ (2012). Importance of visibility when evaluating animal response to roads. Wildl. Biol. 18:393-405. Crossref |
||||
| Morhardt JE, Coulston PJ, Moock S (1984). Comparative Use of Transmission Line Corridors and Parallel Study Corridors by Mule Deer in the Sierra Nevada Mountains of Central California. Pages 614-622 in Crabtree AF, editor. Proc. Third Int. Symp. Environ. Concerns Rights-of-Way Manage., San Diego, California, USA. Mississippi State University. | ||||
| Mysterud A, Ostbye E (1995). Roe deer Capreolus capreolus feeding on yew Taxus baccata in relation to bilberry Vaccinium myrtillus density and snow depth. Wildl. Biol. 1: 249-253. | ||||
|
Nadjafzadeh M, Hofer H, Krone O (2013). The Link Between Feeding Ecology and Lead Poisoning in White-Tailed Eagles. J. Wildl. Manage. 77:48-57. Crossref |
||||
|
Nellemann C, Vistnes I, Jordhøy P, Strand O (2001). Winter distribution of wild reindeer in relation to power lines, roads and resorts. Biol. Conserv. 101: 351-360. Crossref |
||||
|
Nellemann C, Vistnes I, Jordhøy P, Strand O, Newton A (2003). Progressive impact of piecemeal infrastructure development on wild reindeer. Biol. Conserv. 113: 307-317. Crossref |
||||
| Crossref | ||||
|
Niering WA, Goodwin RH (1974). Creation of Relatively Stable Shrublands with Herbicides: Arresting "Succession" on Rights-of-Way and Pastureland. Ecology. 55: 784-795. Crossref |
||||
|
Norberg H, Kojola I, Aikio P, Nylund M (2006). Predation by golden eagle Aquila chrysaetos on semi-domesticated reindeer Rangifer tarandus calves in Northeastern Finnish Lapland. Wildl. Biol. 12: 393-402. Crossref |
||||
| Norwegian Mapping Authority (2012). VBASE: Alle kjørbare veier. | ||||
| Nybakk K, Kjelvik A, Kvam T, Overskaug K, Sunde P (2002). Mortality of semi-domestic reindeer Rangifer tarandus in central Norway. Wildl. Biol. 8:63-68. | ||||
|
Nybakken L, Selas V, Ohlson M (2013). Increased growth and phenolic compounds in bilberry (Vaccinium myrtillus L.) following forest clear-cutting. Scand. J. For. Res. 28: 319-330. Crossref |
||||
|
Panzacchi M, Van Moorter B, Jordhøy P, Strand O (2013). Learning from the past to predict the future: using archaeological findings and GPS data to quantify reindeer sensitivity to anthropogenic disturbance in Norway. Landscape Ecol. 28: 847-859. Crossref |
||||
| Paquet PC, Wierzchowski J, Callaghan. C (1996). Effects of Human Activity on Gray Wolves in the Bow River Valley, Banff National Park, Alberta. Pages 7-i - 7.A-11 in Green JC, Cornwell LP, Bayley S, editors. Ecological Outlooks Project. A Cumulative Effects Assessment and Futures Outlook of the Banff Bow Valley. Department of Canadian Heritage, Ottawa, Ontario, Canada. | ||||
| Perry MC, Osenton PC, Fallon FW, Fallon JE (1997). Optimal Management Strategies for Biodiversity Within a Powerline Right-of-Way. Pages 133-139 in Williams JRR, Goodrich-Mahoney JW, Wisniewski JR, Wisniewski J, editors. Proc. Sixth Int. Symp. Environ. Concerns Rights-of-Way Manage., New Orleans, Louisiana, USA. Elsevier Science Ltd., Oxford, UK; New York, USA; Tokyo, Japan. | ||||
|
Powell AS, Lindquist ES (2011). Effects of Power-line Maintenance on Forest Structure in a Fragmented Urban Forest, Raleigh, NC. Southeast. Nat. 10: 25-38. Crossref |
||||
|
Prather PR, Messmer TA (2010). Raptor and Corvid Response to Power Distribution Line Perch Deterrents in Utah. J. Wildl. Manage. 74:796-800. Crossref |
||||
|
Rea RV, Gillingham MP (2001). The impact of the timing of brush management on the nutritional value of woody browse for moose Alces alces. J. Appl. Ecol. 38:710-719. Crossref |
||||
|
Reimers E, Colman JE (2009). Reindeer and caribou (Rangifer tarandus) response towards human activities. Rangifer. 26:55-71. Crossref |
||||
|
Reimers E, Dahle B, Eftestol S, Colman JE, Gaare E (2007). Effects of a power line on migration and range use of wild reindeer. Biol. Conserv. 134:484-494. Crossref |
||||
|
Reimers E, Miller FL, Eftestol S, Colman JE, Dahle B (2006). Flight by feral reindeer Rangifer tarandus tarandus in response to a directly approaching human on foot or on skis. Wildl. Biol. 12:403-413. Crossref |
||||
|
Reimers E, Roed KH, Colman JE (2012). Persistence of vigilance and flight response behaviour in wild reindeer with varying domestic ancestry. J. Evol. Biol. 25:1543-1554. Crossref |
||||
| Ricard JG, Doucet GJ (1995). Moose (Alces alces) Harvest by Recreational Hunting Near Powerline Rights-of-Way in Québec. Pages 323-324 in Doucet GJ, Séguin C, Giguère M, editors. Proc. Fifth International Symposium on Environmental Concerns in Rights-of-Way Management, Montréal, Québec, Canada. Vice Présidence Environnement Hydro-Québec, Montréal, Québec, Canada. | ||||
| Ricard JG, Doucet GJ (1999). Winter Use of Powerline Rights-of-Way by Moose (Alces alces). Alces. 35:31-40. | ||||
|
Rieucau G, Vickery WL, Doucet GJ, Laquerre B (2007). An innovative use of white-tailed deer (Odocoileus virginianus) foraging behaviour in impact studies. Can. J. Zool.-Rev. Can. Zool. 85: 839-846. Crossref |
||||
| Rumble MA, Benkobi L, Gamo RS (2005). Elk Responses to Humans in a Densely Roaded Area. Intermountain J. Sci. 11: 10-24. | ||||
| Seiler A (2001). Ecological Effects of Roads: A Review. Introductory Research Essay. Department of Conservation Biology, Uppsala University, Uppsala, Uppland, Sweden. 40 pp. | ||||
| Semmler M, Straumann U, Roero C, Teich TH (2005). Tonale Schallemissionen von Hochspannungsfreileitungen: Mechanismus und Reduktionsmassnahmen. Bulletin SEV/VSE 15/05 from ETH Zürich, Zürich, Switzerland. 17 pp. | ||||
|
Spellerberg IF (1998). Ecological effects of roads and traffic: a literature review. Global Ecol. Biogeogr. 7:317-333. Crossref |
||||
|
Stankowich T (2008). Ungulate flight responses to human disturbance: A review and meta-analysis. Biol. Conserv. 141: 2159-2173. Crossref |
||||
| Statistics Norway (2011a). Electricity statistics, annual; Lines at the end of the year, by type of line and voltage. 2009. Km1. Statistics Norway (2011b). Helårsbilveier og sommerbilveier. Total veilengde . | ||||
| Statnett SF. 2013. Nettutviklingsplan 2013. Steenhof K, Kochert MN, Roppe J (1993). Nesting by Raptors and Common Ravens on Electrical Transmission Line Towers. J. Wildl. Manage. 57: 271-281. | ||||
|
Stockwell CA, Bateman GC, Berger J (1991). Conflicts in national parks: A case study of helicopters and bighorn sheep time budgets at the grand canyon. Biol. Conserv. 56: 317-328. Crossref |
||||
| Strand O, Jordhøy P, Solberg EJ (2001). Villreinen og effekter av Rv7 over Hardangervidda. Report 666 from NINA, Trondheim, Norway. 24 pp. | ||||
|
Straumann U (2011). Mechanism of the tonal emission from ac high voltage overhead transmission lines. J. Phys. D: Appl. Phys. 44: 1-8. Crossref |
||||
|
Taraborelli P, Gregorio P, Moreno P, Novaro A, Carmanchahi P (2012). Cooperative vigilance: The guanaco's (Lama guanicoe) key antipredator mechanism. Behav. Processes. 91: 82-89. Crossref |
||||
|
Tarlow EM, Blumstein DT (2007). Evaluating methods to quantify anthropogenic stressors on wild animals. Appl. Anim. Behav. Sci. 102:429-451. Crossref |
||||
| Teich TH, Weber HJ (2002). Origin and abatement of tonal emission from high voltage transmission lines. e&i Elektrotechnik und Informationstechnik. 119:22-27. | ||||
|
Tsujimoto K, Furukawa S, Shimojima K, Yamamoto K (1991). Development of ns-tacsr with extremely suppressed aeolian noise and its application to 500-kV overhead transmission-line. IEEE Trans. Power Delivery. 6:1586-1592. Crossref |
||||
| Tyler N, Stokkan K-A, Hogg C, Nellemann C, Vistnes A-I, Jeffery G (2014). Ultraviolet Vision and Avoidance of Power Lines in Birds and Mammals. Conserv. Biol. 00:1-2. | ||||
|
Vistnes I, Nellemann C (2001). Avoidance of Cabins, Roads, and Power Lines by Reindeer during Calving. J. Wildl. Manage. 65: 915-925. Crossref |
||||
|
Vistnes I, Nellemann C (2008). The matter of spatial and temporal scales: a review of reindeer and caribou response to human activity. Polar Biol. 31: 399-407. Crossref |
||||
|
Vistnes I, Nellemann C, Jordhøy P, Støen OG (2008). Summer distribution of wild reindeer in relation to human activity and insect stress. Polar Biol. 31:1307-1317. Crossref |
||||
|
Vistnes I, Nellemann C, Jordhøy P, Strand O (2001). Wild reindeer: impacts of progressive infrastructure development on distribution and range use. Polar Biol. 24: 531-537. Crossref |
||||
|
Vistnes I, Nellemann C, Strand O (2004). Effects of Infrastructure on Migration and Range Use of Wild Reindeer. J. Wildl. Manage. 68:101-108. Crossref |
||||
| Willyard CJ, Tikalsky SM (2004). Research Gaps Regarding the Ecological Effects of Fragmentation Related to Transmission-Line Rights-of-Way. Pages 521-527 in Goodrich-Mahoney JW, Abrahamson LP, Ballard J, editors. Proc. Eighth Int. Symp. on Environ. Concerns in Rights-of-Way Manage., Saratoga Springs, New York, USA. Elsevier Ltd., Amsterdam, Netherlands. | ||||
| Wingfield JC, Hunt K, Breuner C, Dunlap K, Fowler GS, Freed L, Lepson J (1997). Environmental stress, field endocrinology, and conservation biology. Pages 95-131 in Clemmons J, Buchholz R, editors. Behavioral Approaches to Conservation in the Wild. Cambridge University Press, Cambridge. | ||||
|
Wolfe SA, Griffith B, Wolfe CAG (2000). Response of reindeer and caribou to human activities. Polar Res. 19: 63-73. Crossref |
||||
| Wood P, Wolfe ML (1988). Intercept Feeding as a Means of Reducing Deer-Vehicle Collision. Wildl. Soc. Bull. 16: 376-380. | ||||
|
Ydenberg RC, Dill LM (1986). The Economics of Fleeing from Predators. Adv. Study Behav. 16:229-249. Crossref |
||||
|
Ytrehus B, Skagemo H, Stuve G, Sivertsen T, Handeland K, Vikoren T (1999). Osteoporosis, bone mineralization, and status of selected trace elements in two populations of moose calves in Norway. J. Wildl. Dis. 35:204-211. Crossref |
||||
APPENDIX
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


