Full Length Research Paper

ABSTRACT
Secondary forests may act as buffer area and serve as reservoir for biotic components that are lost from primary forest due to anthropogenic disturbances. This study investigated the floristic composition, diversity and community structure of Cocoa Research Institute of Nigeria (CRIN), Idi-Ayunre, Ibadan, Nigeria. Twenty-five main plots (each 50 m × 50 m) were randomly mapped out to enumerate tree species, five 10 m × 10 m sub-plots were systematically mapped out within each main plot to enumerate shrubs and three quadrats (1 m × 1 m) were laid in each sub-plot to enumerate herbaceous species in Wet Season (WS) and Dry Season (DS). Relative Importance Value (RIV), Taxa, Individuals, Dominance, Shannon-Wiener, Equitability and Jaccard similarity index were determined. A total of 181 plant species from 145 genera and 54 families which included 63 trees, 33 shrubs and 85 herbaceous species were enumerated. In wet season, Triplochiton scleroxylon, Lonchocarpus griffonianus and Chromolaena odorata had the highest RIV while in dry season, Terminalia superba, Lonchocarpus cyanescens and C. odorata were the highest for trees, shrubs and herbs, respectively. Low dominance but high equitability and Shannon-Weiner values indicated inter-specificity among trees, shrubs and herbs. It was only in herbs that Jaccard-similarity was less than 100% across seasons. Resilience for keystone species conservation is possible due to flora species heterogeneity of the study site.
Key words: Forest ecosystem, biodiversity, anthropogenic activities, relative importance value, ecosystem services.
INTRODUCTION
Secondary forests constitute a large and growing component of forest cover and are crucial in the provision of various goods and services in the ecosystem (Jenkins and Schaap, 2018). Forest resources exploitation and associated industries create employment and revenue base which suggests its roles in socio-economic development policies (World Bank, 2016). Forest ecosystems provide habitat for biodiversity, provides food stuff and other important resources for humans to survive on land (Atomsa and Dibbisa, 2019). Since biodiversity occupies a crucial role in provision of goods and services for forest ecosystems, human activities through exploitation of some of the available goods and services symbolizes ecosystem degradation which is inimical to biodiversity conservation. The increasing exploitation of tropical rainforests for silviculture and anthropogenic activities such as habitation, agriculture and industry are devastating and of a great concern due to their ecological impacts (Monarrez-Gonzalez et al., 2020). It has been reported that conversion and degradation of forest for agriculture has destroyed about 55% of the world's original area of tropical moist forest leading to biodiversity loss across the world (Green et al., 2005; Awodoyin et al., 2013). Impact of forest conversions includes changes in soil, vegetation, hydrological functions, climate change and depletion of natural resources and biodiversity (Jenkins and Schaap, 2018). Halting anthropogenic activities such as timber harvesting, logging, agriculture and promotion of sustainable forestry management practices would contribute to the resilience of a secondary forest in biodiversity and other ecosystem services (Olajuyigbe and Jeminiwa, 2018). However, adequate monitoring, protection and sustainable management of a tropical secondary rainforest require a good knowledge of flora component, diversity and structure (Fang Zeh et al., 2019). Information on the floristic composition and structure of the secondary forest is key to understanding the larger dynamics of forest area and finding key elements of plant diversity (Akinyemi and Oke, 2014; Van Rooyen et al., 2016). Therefore, changes in species distributions, composition and abundance are measured through biological components of an ecosystem (O’Connor et al., 2017). Awodoyin et al. (2013) enumerated flora species composition, diversity and community structures in Cocoa plots in Nigeria, as an inventory of Nigeria flora status due to agriculture. Studies on floristic assessment will assist in the current state and future prediction of an ecosystem. This study enumerated composition, diversity and community structure of herbs, shrubs and trees in secondary forest of Cocoa Research Institute of Nigeria (CRIN), Idi-Ayunre, Ibadan, Nigeria.
MATERIALS AND METHODS
The study was carried out in the secondary Forest of Cocoa Research Institute of Nigeria (CRIN), Idi-Ayunre, Ibadan, Oyo State, Nigeria (Figure 1). The CRIN was established by the Federal Government of Nigeria through the Nigeria Research Institute in 1964 (Atanda, 1977). The CRIN estate falls within the transitory rainforest-savanna vegetation of Nigeria and located within coordinates 07° 12.157’ and 07° 13.260’ North and 03° 51.093’ and 03° 52.290’ East (Obatolu, 2014). Sampling was conducted in 25 quadrats that were randomly located and established using Global Positioning System (GPS; Garmin Map78 model).
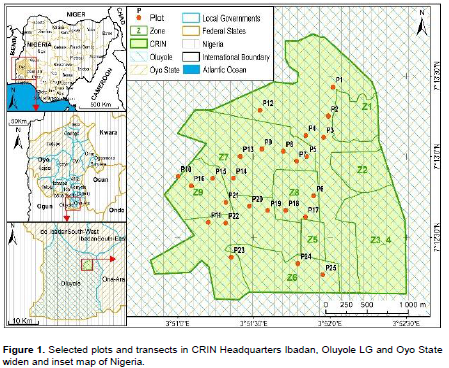
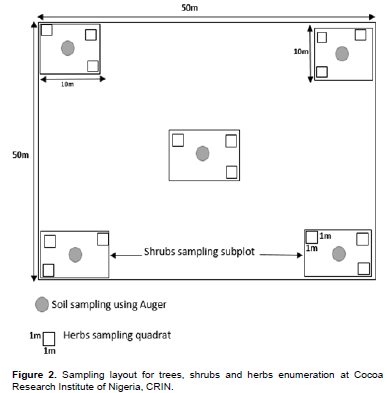
The geographical coordinates of each main quadrat (50 m × 50 m) (Figure 2) were recorded using Garmin map 78 GPS in a total area of 482.63 ha of CRIN stratified zones. The number of plots selected in each zone depended upon area of the forest fragment, and it was 2, 3, 4, 5 or 6 sampled plots per zone. A total of 25 main quadrats (50 m × 50 m) were established per season. The quadrat size established for the enumeration depended on plant life form. Trees were enumerated within each main quadrat (50 m × 50 m). Shrub species were enumerated in five sub-quadrats (10 m × 10 m each) laid within each main quadrat. The sub-quadrats were located at the four corners and one at the centre of the main quadrat. A total of 125 sub-quadrats per season were established for enumeration of shrubs. Three sub-sub-quadrats of 1 m × 1 m each were located randomly in each sub-quadrat to enumerate herbaceous species. Hence, a total of 375 sub-sub-quadrats were laid to collect data on herbaceous species per season. A complete inventory of plant species was done in the main quadrat (trees), sub-quadrat (shrubs) and sub-sub-quadrats (herbaceous) and was carried out between July and August for the wet season and January to February for the dry season. The trees and shrub plants were identified by their local and scientific names with the help of a forester who speaks the local language (Yoruba) and the herbaceous were identified with the help of a flora on West African weeds (Akobundu et al., 2016) and those that could not be identified were collected each day, preserved and taken to the CRIN Laboratory for identification with the help of a taxonomist.
Data analysis
From the data, measure of relative importance value for each species in each site was determined as mean of relative density and relative frequency for each species following the methods in Kent and Coker (1992) and Awodoyin and Egberongbe (2010).
[(RD + RF)/2] × 100 (1)
where RD=Relative Density and RF=Relative Frequency.
RD = (d/D) × 100 (2)
where d = the density of a species; D = total density of all species.
RF = (f/F) × 100 (3)
where f = the frequency of a species; F = total frequency of all species.
Also, the biological community structure as informed by the ecological diversity of the plant species was determined by alpha and beta diversity. The alpha diversity, which is the diversity of species within a particular community, was determined by Species Richness (R), Shannon-Wiener (H`), Evenness (J) and Dominance (D) indices using Paleontological Statistics (PAST) software version 3.0 (Hammer, 2011). The beta diversity, which is the expression of between-habitat diversity, was determined by the Jaccard index of similarity (Spellerberg, 1993).
Species richness is the total number of species occurring within a specified area of the community.
The Shannon-Wiener index of species diversity is calculated as:
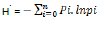
where pi is the proportion which is the number of individuals in a species (n) in relation to the total number of all individuals in a community (N):
pi = n/N (5)
ln is the Naperian logarithm ln = 2.303×log10 (6)
The species equitability index (J) is calculated as:
J = H`/lnS (7)
where H` is Shannon-Wiener index and S is total number of species in the community. The Dominance index (D) is calculated as:
D = ∑ (n/N)2 (8)
where n is number of individuals of a particular species and N is the total number of individuals found in the community.
Jaccard index of community similarity (SCj) is calculated as:
SCj (%)=W/(A+B-W)×100 (9)
where W is the number of common species, A is the number of species in community A, and B is the number of species in community B.
RESULTS AND DISCUSSION
The result of enumeration gave a total number of 181 plant species including 85 herbaceous (Table 1), 33 shrubs (Table 2) and 63 trees (Table 3) belonging to 145 genera and 54 families (21 for herbaceous, 17 for shrubs and 29 for trees) in both wet and dry seasons. Among these families, Fabaceae had the highest number of species (25) followed by Poaceae (20 species), Asteraceae and Rubiaceae had 12 species each, Euphorbiaceae (11 species), Malvaceae (10 species), Apocynaceae (seven species), Moraceae and Solanaceae families had six species each, Cyperaceae (five species), Acanthaceae and Commelinaceae had four species each, Combretaceae, Nyctaginaceae, Sapindaceae and Ulmaceae had three species each.

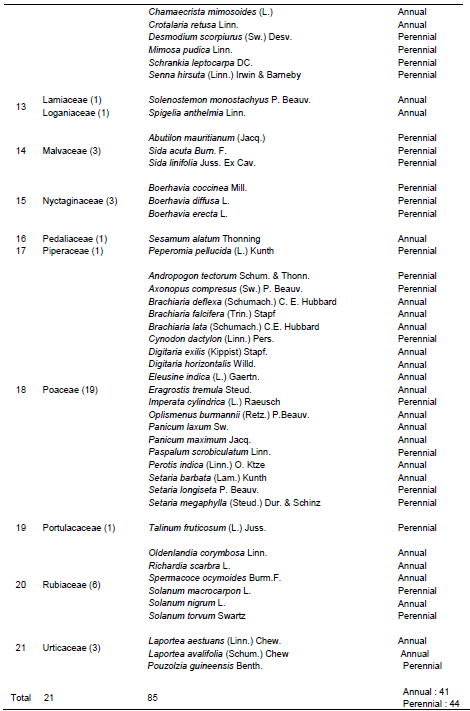


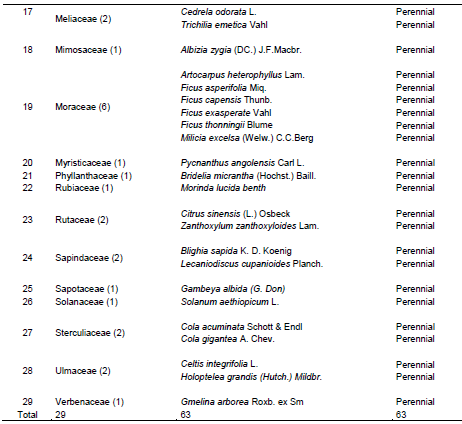
Eleven families namely Amaranthaceae, Boraginaceae, Connaraceae, Convolvulaceae, Irvingiaceae, Meliaceae, Rutaceae, Tiliaceae, and Verbenaceae had two species each while the remaining 23 families like Agavaceae, Anacardiaceae, Annonaceae, Bignoniaceae, Bombacaceae, Capparidaceae, Clusiaceae, Cucurbitaceae, Dennstaedtiaceae, Dioscoreaceae, Icacinaceae, Lamiaceae, Loganiaceae, Myristicaceae, Meliaceae, Musaceae, Ochidaceae, Pedaliaceae, Phyllanthaceae, Piperaceae, Portulacaceae, Sapindaceae, and Sapotaceae had one species each.
The micro-habitat of CRIN secondary forest was closely similar in vegetation types and species composition but varied slightly in individual species (structure) enumerated in the wet and dry seasons. The species composition in shrubs and trees species was similar in the wet and dry season, but there was difference in the herbaceous species composition in the wet and dry seasons. The finding of this study indicated high species richness and abundance for herbs, shrubs and trees species but herbaceous had the highest species richness and abundance. This was similar to the findings of Komolafe et al. (2017) who opined that Ibodi forest of Southwest Nigeria contains 47 herbs and 93 trees while Tang et al. (2010) reported high species composition of 222 plant species in secondary vegetation communities of China comprising 113 herbaceous species, 109 woody species (trees, shrubs, and lianas) with 79 families and 183 genera. The high species composition and abundance may be attributed to human perturbations in the CRIN forest. This was similar to the assertion of Mishra et al. (2008) that herbaceous species dominance over shrubs and trees composition in a disturbed forest may be attributed to the presence of anthropogenic activities including the harvest of food tree plantations (Oluwatosin and Jimoh, 2016). Also, Kessler et al. (2005) attributed the presence of high number of herbaceous flora in a recovery forest to the presence of few mature trees, which allows the penetration of light thereby promoting the growth of light-loving plants known as heliophytes (Bobo et al., 2006; Awodoyin et al., 2013). The most abundant number of herbaceous species was found in Poaceae and Asteraceae in both seasons while Rubiaceae and Fabaceae contained the most abundant shrub species and Fabaceae, Moraceae and Malvaceae were the most abundant tree species. The abundance of Asteraceae in both wet and dry season conforms to the study of Awodoyin et al. (2013) who reported that Asteraceae was found to be among the highest families of herbs in cocoa agroforest of tropical rainforest zone of Nigeria and Ahmed et al. (2015) who reported same for semi-desert region of Egypt. Notwithstanding the number of species identified for herbs, shrubs and trees, the abundance of individuals for most herb species was higher in the wet season than the dry season, which may be attributed to adequate moisture availability.
Relative importance values of herbaceous species in wet and dry seasons of CRIN Secondary Forest, Ibadan, Nigeria
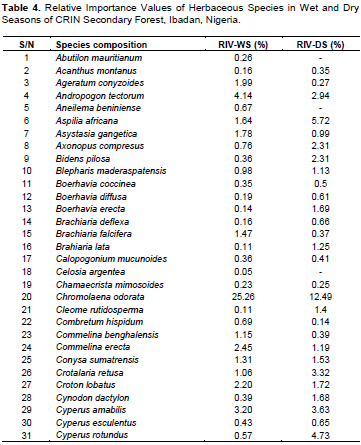
The result showed the herbaceous species that were enumerated (85 species in wet season and 69 species in dry season). Among the herbaceous species identified, Chromolaena odorata had the highest Relative Importance Value (RIV) of 25.26% (WS) and 12.27% (DS), followed by Cyperus rotundus (8.13%) in wet, Aspilia africana (5.72%) in dry, Setaria megaphylla (5.14% in wet) and (1.31% in dry) (Table 4). The lowest RIV belongs to Panicum maximum with 0.03% in wet season and Combretum hispidum with 0.14% in dry season.
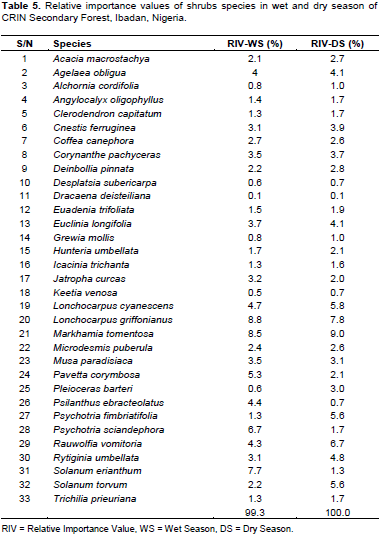
Table 5 shows the shrub species in both seasons. Lonchocarpus griffonianus had 8.8% RIV (the highest in wet season) and 7.8% of RIV in dry season (Table 5). Lonchocarpus cyanescens had 9.0% of RIV (the highest in dry season) and 8.5% of RIV in wet season. Solanum erianthum had 7.7 and 1.33% of RIV in wet and dry season while Rauwolfia vomitoria had 6.7 and 4.3% of RIV in dry and wet season, respectively. The lowest RIV belonged to Dracaena deisteiliana with 0.1% of RIV in wet and in dry season followed by Keetia venosa 0.5 and 0.7% of RIV in wet and dry season, respectively.

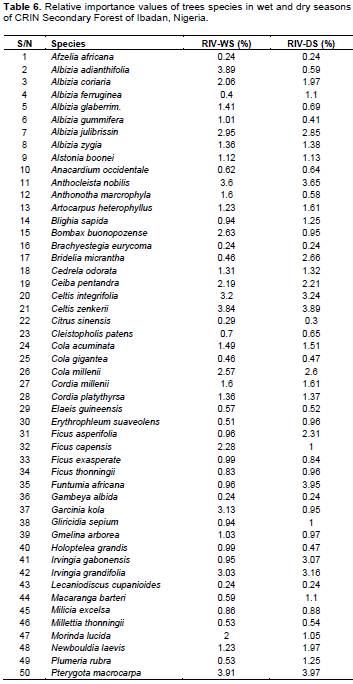
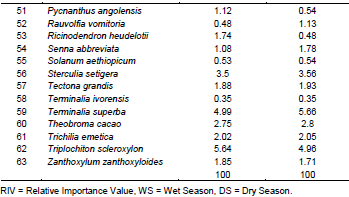
A total of sixty-three trees species were enumerated in the wet and dry seasons (Table 6). The most abundant species was Triplochiton scleroxylon with 5.64% of RIV (the highest in wet season) and 4.96% of RIV in dry season (Table 6). Terminalia superba followed with 5.66% of RIV (the highest in dry season) and 4.99% of RIV in wet season followed by Pterygota macrocarpa which had 3.91 and 3.97% of RIV in wet and dry season while Cedrela odorata had 1.31 and 1.32% of RIV in wet and dry season, respectively. The lowest common RIV (0.24%) in wet and dry season belonged to the four species namely Afzelia africana, Brachyestegia eurycoma, Gambeya albida and Lecaniodiscus cupanioides.
The heavy presence of C. odorata might have influenced the number of herbaceous species composition in the CRIN forest. The high occurrence of C. odorata, a ubiquitous species of disturbed forest in the rainforest supports the assertion of Oke and Isichei (1997) who reported that C. odorata is a common herb of tropical rainforest. The C. odorata like many in Asteraceae family has the peculiar intense competitive ability, fast growth, efficient seed dispersal mechanism and high regeneration rate. The invasion of the study site by C. odorata might have influenced the number of herbaceous species flora due to previous land-use change, especially deforestation for cultivation. This agrees with the findings of Agboola and Muoghalu (2015) that reported a decrease in plant species composition and diversity in sites invaded by Tithonia diversifolia and C. odorata.
The high number of Rubiaceae among shrub species composition in the secondary rainforest of CRIN was supported by Ndah et al. (2013) who mentioned that Rubiaceae was the most prominent family of shrub in species composition, diversity and distribution in a disturbed Takamanda Rainforest, South West, Cameroun. The RIV of L. griffonianus (Fabaceae) remains prominent in both wet and dry season but it was second in its occurrence to L. cyanescens (Fabaceae) in the dry season. L. griffonianus and L. cyanescens (Fabaceae) are pioneer species that have fast regeneration ability. They are usually abundant in disturbed forest or forest in their transition state (Bobo et al., 2006). However, the low number of L. griffonianus in the dry season may be its inability to tolerate vagaries of weather, resource and nutrient competition. This was supported by the findings of Ogwu et al. (2016) who reported that limited access to water resources may affect the survival of young trees (shrubs) in the dry season. Similarly, L. cyanescens which belongs to Fabaceae alongside L. griffonianus may be a better nitrogen pump in the dry season where the decomposition by microorganism would be low due to low humidity. In a tropical rainforest, some Fabaceae like L. cyanescens and L. griffonianus have root nodules that could sequester and fix atmospheric nitrogen into the soil with the aid of Rhizobium. This was corroborated by Chen (2006) who reported seasonal (water and temperature) influence on the performance of nitrogen fixation. Fabaceae had the highest number of tree species (12), followed by Moraceae and Malvaceae, each with six species. The high number of species composition of Fabaceae and Moraceae in Nigeria rainforest has widely been documented (Salami and Lawal, 2018). This aligns with the findings of Adekunle et al. (2010) and Adekunle (2016) who reported that families such as Sterculiaceae, Meliaceae, Moraceae and Ebenaceae dominate the tropical rainforest of southwest Nigeria. The ability of Moraceae to produce a large number of seeds and quickly establish itself may account for their high presence and this was confirmed by Deka et al. (2012). Also, the high species composition of Fabaceae may be attributed to high competitive ability for water and other growth resources (Ogwu et al., 2016). The trees with the highest RIV values in both wet and dry seasons were the most commonly found trees in the study site; T. scleroxylon (Malvaceae) and T. superba (Combretaceae). Therefore, trees with such economic importance to human may have suffered exploitation and responsible for the decline in the number of tree species in a secondary forest. This assertion was supported by Komolafe et al. (2017), who reported that the economic value of trees may influence its selection for exploitation.
Species diversity indices in wet and dry seasons
The diversity indices revealing the plant community structure at CRIN secondary forest in wet season of 2018 and dry season of 2019 are presented in Table 7. The results obtained revealed that the highest numbers of taxa (87) and individuals (26,304) were obtained among herbaceous species in the wet season. However, trees had similar higher number of species (64) in wet and dry season than shrubs that also had similar number of species (49) in the wet and dry season. However, shrubs had higher number of individuals in wet (3481) and dry (2617) seasons than trees in wet (953) and dry (928) seasons. Dominance values 0.16, 0.03 and 0.03 in the wet season and 0.06, 0.03, and 0.03 in the dry season, respectively for herbaceous, shrubs and trees were low. The low values indicates that there was no predominance of a single species, all species were randomly distributed in the area in both seasons. Simpson index values 0.84, 0.97 and 0.97 in the wet season and 0.94, 0.97, 0.97 in dry season respectively for herbaceous, shrubs and trees were very high, which revealed the richness of species in the area in both seasons. The Shannon index (H’) values were 2.94, 3.56 and 3.81 in the wet season and 3.54, 3.63, and 3.80 in dry season, respectively for herbaceous, shrubs and trees were high and close to the highest value in each life form type. The values obtained indicate that there are many species in both seasons, each with few individuals that are randomly distributed and none was dominant. Equitability index was higher in herbaceous in the dry season (0.83) than wet season (0.66) and very high in shrubs in the dry season (0.93) and wet season (0.92) and trees in the wet season (0.92) and dry season (0.91). These values indicate that individuals are randomly distributed among all species in each season. There was highest similarity index (100%) between shrubs enumerated in the wet season (WSS) and shrubs enumerated in the dry season (DSS) on one hand and trees enumerated in the wet season (WST) and trees enumerated in dry season trees (DST) on another hand (Table 8). This was followed by the similarity index (76.40%) observed between WSH and DSH while the lowest similarity (1.03%) was observed between WSS and DSS compared to WST and DST. There were no similarity among herbaceous compared to shrubs and trees, respectively in both seasons.

The CRIN forest is an ecosystem with a high diversity of species in distribution and abundance. This was explained by the high Shannon-Wiener values (2.94 - 3.81) obtained for herbs, shrubs and trees in both wet and dry seasons. There were, however, deviations across seasons for the diversity of each of shrubs, herbs and trees. The trees had the highest diversity in the wet season but the shrubs had the highest diversity in the dry season. There was insignificant diversity in both wet and dry season for herbaceous species. The seasonal similarity in herbs diversity may be attributed to the fallow management in a historically less stressed landscape. This supported the findings that soil conditions, inadequate rainfall pattern and excessive temperature may alter diversity and allows the support of few numbers of herbaceous flora in the forest ecosystem (Thakur, 2018). The forest is diverse with mixed species of herbs, shrubs and trees. This was responsible for the low dominance values in the vegetation structure (herbs, trees and shrubs) of both seasons in the CRIN forest. The seasonal changes did not affect dominance of trees, but shrubs and herbs species dominance increased in the dry season. The few species that can survive the low moisture in the dry season were all well represented. This was also explained by high evenness values obtained for trees, shrubs and herbs in both seasons. The seasonal variation in species diversity of herbs and shrubs agreed with the report of Lu et al. (2010) that suggested high species diversity of herbs with seasonal changes. Also, the increase in diversity of species of herbs and shrubs in the dry season as shown by evenness values may further be attributed to other factors apart from the climate. Moreno and Halffter (2001) and Tuomisto et al. (2003) suggested that landscape pattern and climatic conditions may influence species diversity of an ecosystem. It was similarly reported that ecological legacies including anthropogenic perturbation especially deforestation, intermediate succession and regeneration potential in the ecosystem of a forest which has not reached is a climax. Though herbs had less number of taxa in the dry season than wet season, the diversity was higher in the dry season than wet season. The implication of this result is that the few species that survived the stress of dry season were all randomly distributed with no particular species dominant. This agreed with the findings of Thakur (2018) that the diversity of herbaceous species in the mixed forest was highest in a study conducted in the dry tropical forest of India. The CRIN forest recorded high species richness (taxa) for herbs, shrubs and trees in both seasons. The species richness of herbs was high in the wet season but for the species richness of shrubs and trees, there was no difference across the season. The openness and closure of forest may influence species richness. The secondary forest may have a closed canopy of trees in the wet season, which may favour the shade loving species (sciophytes), annuals and perennials. However, the open canopy in the dry season may result in the loss of sciophytes and annuals. This was further stressed in the reports of Tang et al. (2010) and Awodoyin et al. (2013) that varying herbaceous species that are shade-loving (sciophytes), annuals and perennial plants are favoured in a closed canopy forest. However, high species diversity of trees indicates a resilience forest with mixed tree species population which may, however, enhance vegetation restoration for the continuous growth of the forest to climax (Tang et al., 2010).
CONCLUSION
This study revealed a recovering forest from previous degradation due to unsustainable anthropogenic practices. The flora species abundance and composition indicated heterogeneity among trees, herbs and shrubs enumerated. The presence of C. odorata in both wet and dry season indicated an ubiquitous herbaceous species which must have been introduced through anthropogenic activities since the plant is a known invasive species in Nigeria. Meanwhile, T. scleroxylon and T. superba are commonest trees species enumerated in wet and dry season, respectively. These are endemic tree species of importance in Nigeria, suggesting presence of degraded primary forest remnants. The abundance of shrub species however suggested the potential of the forest for resilience. High species diversity and similarity in species across seasons also indicated wide and random distribution of various herbs, shrubs and tree species in the study site. It is recommended that endangered trees endemic to Nigeria such as T. scleroxylon and T. superba should be adequately protected for sustainable forest resources conservation. Invasive species such as C. odorata should be managed in order to prevent homogeneity of the forest landscape. CRIN secondary forest has a potential to improve biodiversity and other ecosystem goods and services due to its resilience potential with its high diversity. Therefore, it should be protected from illegal exploitation of forest resources in order to ensure an in-situ conservation of genetic resources present.
ACKNOWLEDGEMENTS
The authors are very grateful to the Economic Community of West African States (ECOWAS) and its Department of Education, Science and Culture for the financial support through their scholarship programme.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Adekunle VAJ, Olagoke AO, Ogundare LF (2010). Rate of timber production in a tropical rainforest ecosystem of south-western Nigeria and its implications on sustainable forest management. Journal of Forestry Research 21(2):225-230. |
|
|
|
|
|
Adekunle VAJ (2016). Conservation of tree species diversity in tropical rainforest ecosystem of south-west Nigeria. Journal of Tropical Forest Science 18(2):91-101. |
|
|
|
|
|
Agboola OO, Muoghalu JI (2015). Changes in species diversity, composition, growth and reproductive parameters of native vegetation invaded by Chromolaena Odorata And Tithonia Diversifolia In Osun State, southwest Nigeria. FUTA Journal of Research in Sciences 11(2):217-230. |
|
|
|
|
|
Ahmed DA, Manal F, Saeed NM, Awad MA (2015). Effect of the recent land use on the plant diversity and community structure of Omayed Biosphere Reserve, Egypt. Global Ecology and Conservation 4:26-37. |
|
|
|
|
|
Akinyemi DS, Oke SO (2014). Floristic composition and structural diversity of Shasha Forest Reserve. Notulae Scientia Biologicae 6(4):433-440. |
|
|
|
|
|
Akobundu IO, Ekeleme F, Agyakwa CW, Ogabie CA (2016). A handbook of West African weeds, Third edition. International Institute of Tropical Agriculture (IITA) 3(381). |
|
|
|
|
|
Atanda OA (1977). The third Nigerian cocoa breeding programme: A progress report. 5th International Cocoa Research Conference Ibadan 1975:85-90. |
|
|
|
|
|
Atomsa D, Dibbisa D (2019). Floristic composition and vegetation structure of Ades forest, Oromia regional state, West Hararghe zone, Ethiopia. Tropical Plant Research 6(1):139-147. |
|
|
|
|
|
Awodoyin RO, Egberongbe RK (2010). Potentials of Sesbania pachycarpa DC as a mulch interplant: Seed germination, early growth and competition with Tithonia diversifolia (Hemsl.) A Gray in Ibadan, Nigeria. Nigerian Journal of Science 44:29-39. |
|
|
|
|
|
Awodoyin RO, Akinyemi CY, Bolanle OO, Antiabong IC (2013). Spatial distribution and abundance of Solanecio biafrae (Olive & Heirne) C. Jefrey and Structure of weed communities in some cocoa plots in Ekiti, Oyo and Cross River States, Nigeria. Ife Journal of Science 15(3):661-676. |
|
|
|
|
|
Bobo KS, Waltert M, Sainge NM, Njokagbor J, Fermon H, Mu M (2006). From forest to farmland?: species richness patterns of trees and understorey plants along a gradient of forest conversion in Southwestern Cameroon. Biodiversity and Conservation 15:4097-4117. |
|
|
|
|
|
Chen X (2006). Tree diversity, carbon storage, and soil nutrient in an Old-Growth Forest at Changbai Mountain, Northeast China. Communications in Soil Science and Plant Analysis 37: 363-375. |
|
|
|
|
|
Deka J, Tripathi OP, Khan ML (2012). Urban growth trend analysis using Shannon Entropy approach: a case study in north east India. International Journal of Geomatics and Geosciences 2(4):1-8. |
|
|
|
|
|
Fang Zeh A, Fuashi NA, Maurice ME (2019). Flora composition, structure and diversity in the Kimbi Fungom National Park, North West Region, Cameroon. Journal of Ecology and the Natural Environment 11(1):1-13. |
|
|
|
|
|
Green RE, Cornell SJ, Scharlemann JPW, Balmford A (2005). Farming and the fate of wild nature. Science 307:550-555. |
|
|
|
|
|
Jenkins M, Schaap B (2018). Forest Ecosystem Services. Background Analytical Study 1. Background study prepared for the thirteenth session of United Nations Forum on Forests 41 p. |
|
|
|
|
|
Kent M, Coker P (1992). Vegetation description and analysis. A pratical approach. Bellhaven Press pp. 76-112. |
|
|
|
|
|
Kessler M, Keßler PJA, Gradstein SR, Bach K, Schmull M, Pitopang R (2005). Tree diversity in primary forest and different land use systems in central Sulawesi, Indonesia. Biodiversity and Conservation 14(3):547-560. |
|
|
|
|
|
Komolafe AA, Adegboyega SA, Akinluyi FO (2017). A review of flood risk analysis in Nigeria. American Journal of Environmental Sciences pp. 157-166. |
|
|
|
|
|
Lu ZJ, Li LF, Jiang MX, Huang HD, Bao DC (2010). Can the soil seed bank contribute to revegetation of the drawdown zone in the Three Gorges reservoir region? Plant Ecology 209(1):153-165. |
|
|
|
|
|
Mishra RK, Upadhyay VP, Mohanty RC (2008). Vegetation ecology of the Similipal biosphere reserve, Orissa, India. Applied Ecology and Environmental Research 6(2):89-99. |
|
|
|
|
|
Monarrez-Gonzalez JC, Gonzalez-Elizondo MS, Marquez-Linares MA, Gutierrez-Yurita PJ, Perez -Verdin G (2020). Effect of forest Management on tree diversity in temperate ecosystem forests in Northern Mexico. PLoS ONE 15(5):e0233292. |
|
|
|
|
|
Moreno CE, Halffter G (2001). Spatial and temporal analysis of α, β and γ diversities of bats -in a fragmented landscape. Biodiversity and Conservation 10(3):367-382. |
|
|
|
|
|
Ndah NR, Andrew EE, Bechem E (2013). Species composition, diversity and distribution in a disturbed Takamanda Rainforest, South West, Cameroon. African Journal of Plant Science 7(12):577-585. |
|
|
|
|
|
O'Connor MI, Gonzalez A, Byrnes JEK, Cardinale BJ, Duffy JE, Gamfeldt L, Griffin JN, Hooper D, Hungate BA, Panquette A, Thompson PL, Dee LE, Dolan KL (2017). A general biodiversity-function relationship is mediated by trophic level. Oikos 126(1):18-31. |
|
|
|
|
|
Obatolu BO (2014). Carbon stock estimation of Theobroma cacoa (Cocoa) varieties in Ibadan, Oyo State, Nigeria. Master of Science (Umbuplished paper). University of Ibadan. |
|
|
|
|
|
Ogwu MC, Osawaru ME, Obayuwana OK (2016). Diversity and abundance of tree Species in the University of Benin, Benin City, Nigeria. Applied Tropical Agriculture 21(3):46-54. |
|
|
|
|
|
Oke SO, Isichei AO (1997). Floristics and structure of the fallow vegetation in the Ile-Ife area of Southwestern Nigeria. Nigerian Journal of Botany 10:37-50. |
|
|
|
|
|
Olajuyigbe SO, Jeminiwa MS (2018). Tree Species Diversity and Structure of Eda Forest Reserve, Ekiti State, Nigeria. Asian Journal of Research in Agriculture and Forestry 2(1): 1-12. |
|
|
|
|
|
Oluwatosin B, Jimoh SO (2016). Pattern of plant species diversity in a dry forest ecosystem of Nigeria. Journal of Forestry Research and Management 13:31-47. |
|
|
|
|
|
Salami KD, Lawal AA (2018). Description of economical trees and shrubs species in Northern part of Nigeria and their potentials. Proceedings of 6th Biennial national conference of the forests and forest products society pp. 136-144. |
|
|
|
|
|
Spellerberg IF (1993). Monitoring ecological change. Cambridge University Press P 334. |
|
|
|
|
|
Tang CQ, Li Y, Zhang Z (2010). Species diversity patterns in natural secondary plant communities and man-made forests in a Subtropical mountainous Karst Area, Yunnan, SW China. International Mountain Society 30(3):244-251. |
|
|
|
|
|
Thakur TK (2018). Diversity, composition and structure of understorey vegetation in the tropical forest of Achanakmaar Amarkantak Biosphere Reserve, India. Environmental Sustainability 1(3):279-293. |
|
|
|
|
|
Tuomisto H, Ruokolainen K, Yli-halla M (2003). Dispersal, environment, and floristic variation of Western Amazonian Forests. Science 299(5604):241-244. |
|
|
|
|
|
Van Rooyen M, Van Rooyen N, Orban B, Nsongola G, Miabangana ES, Gaugris J (2016). Floristic composition, diversity and structure of the forest communities in the Kouilou Département, Republic of Congo. Tropical Ecology 57(4):805-824. |
|
|
|
|
|
World Bank (2016). World Bank Group Forest Action Plan FY16-20. World Bank, Washington DC. License: Creative Commons Attribution CC BY 3.0 IGO. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


