Full Length Research Paper

ABSTRACT
Oti-Keran-Mandouri (OKM) is part of the elephant historical range and one of the priority corridors proposed for elephant conservation in West Africa. However, its potentialities to be a functional corridor are yet to be evaluated in a context of increasing anthropogenic pressure. This study aims at assessing habitat biophysical patterns and fragmentation level. A multicriteria evaluation using fuzzy logic was performed to model elephant habitat suitability and vegetation sampling conducted in 123 plots to describe the habitat. In each plot, the physical parameters of woody plants species were recorded. Biodiversity indices, dendrometric parameters, and diameter structure were computed for each habitat type and compared using Jaccard Index and Kruskal-Wallis test. Habitat fragmentation was assessed using the hypsometric method. Apart from a core area located in the south-east, the remnant good habitat is in small patches. Four habitats were distinguished based on their level of degradation. There is a steady increase in habitat diversity from degraded habitat (Habitat 1) to primary habitat (Habitat 4) with the Shannon index increasing from 0.83 to 1.43 bit. In all the habitats, trees are evenly distributed with an evenness higher than 0.7. Dendrometric parameters are significantly different from one habitat to another (P-value < 0.05) apart from the mean diameter and the average regeneration rate. The suitable habitat for elephant constitutes only 31.5% of the area of OKM. The overall habitat fragmentation is 84.74%. Regeneration rates make an eventual restoration possible but further assessment of the socio-ecological system is needed.
Key words: Elephant, habitat suitability, habitat, fragmentation, conservation.
INTRODUCTION
MATERIALS AND METHODS

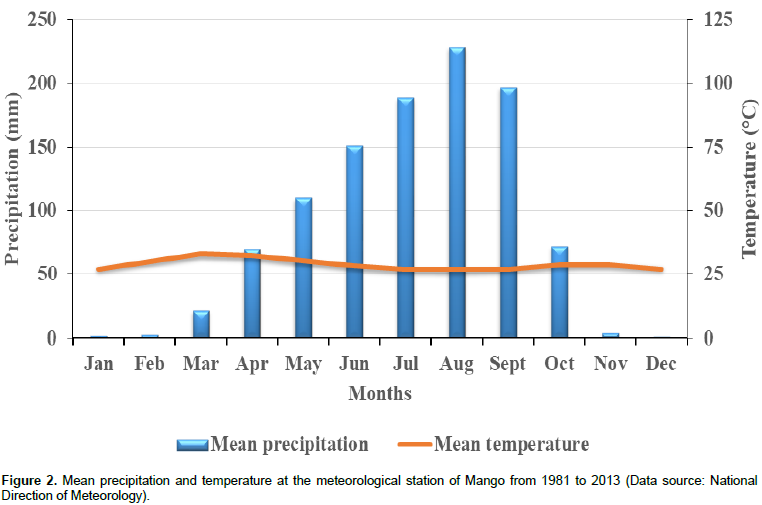
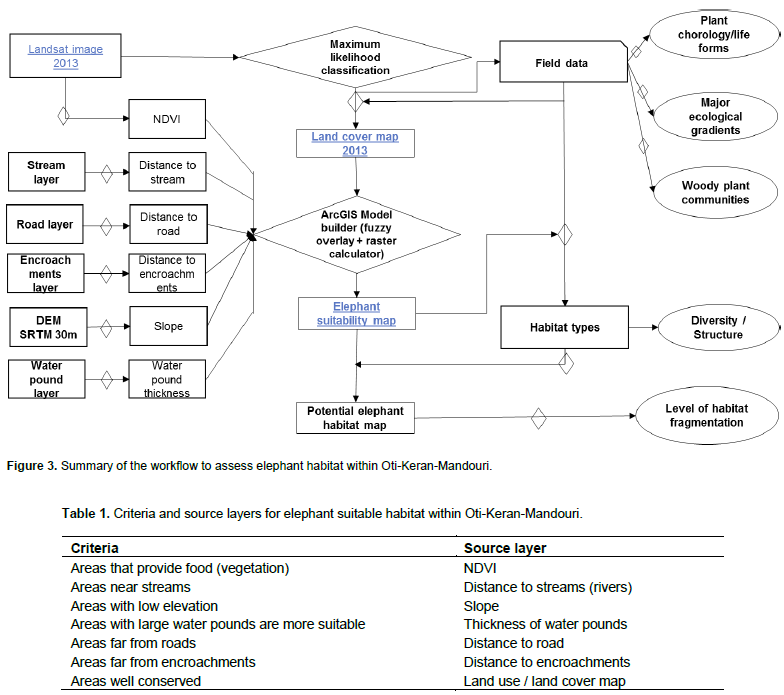

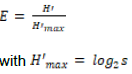
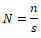
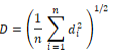


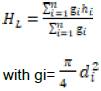


RESULTS






DISCUSSION

Planning for and protecting elephant corridors have been shown to have large benefits for biodiversity conservation by many authors in East Africa especially in Tanzania (Epps et al., 2011; Jones et al., 2012). Furthermore, the African elephant remains a good candidate as a surrogate species for conservation planning. It encompasses all the attributes of a focal species and associated conservation needs as proposed by Brock and Atkinson (2013). An effective conservation of the elephant would require management or mitigation of almost all elements affecting the needs of the total species community and provides ecosystem-level protection. On the other hand, confined population of elephant exert a damaging influence on their habitat. For instance, this effect has already been described in the biosphere reserve of Pendjari where elephants are responsible of inducing bark damage to baobab trees (Adansonia digitata) (Kassa et al., 2013)and to drive spatial isolation in Borassus aethiopum (Salako et al., 2015). A landscape level management of elephant population would alleviate their damaging effect in confined habitat. A corridor between OKM and the nearby block of elephant population in the biosphere reserve of Pendjari could provide a solution to this damaging impact.
CONCLUSION
CONFLICT OF INTERESTS
ACKNOWLEDGEMENTS
REFERENCES
|
Addra TC, Fahem AK, De Jong T, Mank T (1990). Attlas de developpement régional du Togo. Lomé |
|
|
Adjonou K, Bellefontaine R, Kokou K (2009). Les forêts claires du Parc national Oti-Kéran au Nord-Togo : structure, dynamique et impacts des modifications climatiques récentes. Secheresse 20:1-10 doi:10.1684/sec.2009.0217. |
|
|
Akoègninou A, Van der Burg WJ, Van der Maesen LJ (2006). Flore analytique du Bénin. Backhuys Publishers. |
|
|
Aleza K, Wala K, Bayala J, Villamor GB, Dourma M, Atakpama W, Akpagana K (2015). Population structure and regeneration status of Vitellaria Paradoxa (C. F. Gaertner) under different land management regimes in Atacora department, Benin. Agrofor. Syst.:1-13. |
|
|
Atato A, Wala K, Batawila K, Lamien N, Akpagana K (2011). Edible Wild Fruit Highly Consumed during Food Shortage Period in Togo: State of Knowledge and Conservation Status. J. Life. Sci. 5:1046-1057 |
|
|
Barnes RFW (1999). Is there a future for elephants in West Africa? Mamm. Rev 29:175-200 |
|
|
Bartlam-Brooks HLA, Bonyongo MC, Harris S (2011). Will reconnecting ecosystems allow long-distance mammal migrations to resume? A case study of a zebra Equus burchelli migration in Botswana. Oryx 45:210-216 |
|
|
Beier P, Noss RF (1998). Do Habitat Corridors Provide Connectivity? Conserv. Biol. 12:1241-1252 |
|
|
Bierbaum R, Michael S, Henk B, Annette C, Sandra Diaz, Jacob G, Anand P, Ralph S, Guadalupe D, Virginia G, Thomas H., Lev N Christine W (2014). Delivering Global Environ-mental Benefits for Sustainable Development. Report of the Scientific and Technical Advisory Panel (STAP) to the 5th GEF Assembly, México 2014. |
|
|
Boakye EA, Dibi NDH, Barnes RB, Porembski S, Thiel M, Kouamé FN, Kone D (2015). Threat of agricultural production on woody plant diversity in Tankwidi riparian buffer in the Sudanian Savanna of Ghana. Int. J. Biodivers. Conserv. 7:354-363. |
|
|
Boettiger AN, Wittemyer G, Starfield R, Volrath F, Douglas-Hamilton I, Getz WM (2011). Inferring ecological and behavioral drivers of African elephant movement using a linear filtering approach. Ecology 92:1648-1657 |
|
|
Bonou W, Glele Kakai R, Assogbadjo AE, Fonton HN, Sinsin B (2009). Characterisation of Afzelia africana Sm. habitat in the Lama forest reserve of Benin. For. Ecol. Manage 258:1084-1092 |
|
|
Bouché P, Douglas-Hamilton I, Wittemyer G, Nianogo AJ, Doucet J-L, Lejeune P, Vermeulen C (2011). Will Elephants Soon Disappear from West African Savannahs? PLoS One 6:e20619. |
|
|
Brock B, Atkinson EC (2013). Selecting species as targets for conservation planning. In: Craighead FL, Convis CLJ (eds) Conservation Planning: Shaping the Future. First ed. ESRI Press, Redlands, California. pp 123-147 |
|
|
Brunel J, Hiepko P, Scholz H (1984). Flore analytique du Togo: Phanérogames. Eschborn ed. GTZ. doi:10.2307/3776742. |
|
|
Chao A, Chazdon RL, Colwell RK, Shen T-J (2005). A new statistical approach for assessing similarity of species composition with incidence and abundance data. Ecol. Lett. 8:148-159. |
|
|
Cheke RA (2001). Togo. In: Fishpool LDC, Evans MI (eds) Important Bird Areas in Africa and associated islands: Priority sites for conservation. BirdLife Conservation Series vol 11. Pisces Publications and BirdLife International Newbury and Cambridge, UK. pp. 947-952 |
|
|
De Cáceres M (2013). How to use the indicspecies package (ver. 1.7.1). |
|
|
De Cáceres M, Legendre P (2009). Associations between species and groups of sites: indices and statistical inference. Ecology 90:3566-3574. |
|
|
Dimobe K, Wala K, Batawila K, Dourma M, Woegan YA, Akpagana K (2012). Analyse spatiale des différentes formes de pressions anthropiques dans la réserve de faune de l'Oti-Mandouri (Togo). VertigO-la revue électronique en sciences de l'environnement. |
|
|
Dimobe K, Wala K, Dourma M, Kiki M, Woegan Y, Folega F, Batawila K, Akpagana K (2014). Disturbance and Population Structure of Plant Communities in theWildlife Reserve of Oti-Mandouri in Togo (West Africa). Annu. Res. Rev. Biol. 4:2501-2516 |
|
|
Djossa BA, Fahr J, Wiegand T, Ayihouénou BE, Kalko EK, Sinsin BA (2007). Land use impact on Vitellaria paradoxa C.F. Gaerten. stand structure and distribution patterns: a comparison of Biosphere Reserve of Pendjari in Atacora district in Benin. Agrofor. Syst. 72:205-220. |
|
|
Dufrene M, Legendre P (1997). Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 67:345-366 |
|
|
Epps CW, Mutayoba BM, Gwin L, Brashares JS (2011). An empirical evaluation of the African elephant as a focal species for connectivity planning in East Africa. Divers. Distrib. 17:603-612 |
|
|
Ern H (1979). Die Vegetation Togos. Gliederung, Gefährdung, Erhaltung. Willdenowia. pp. 295-312. |
|
|
ESRI (2014) ArcGIS, 10.3 ed., Redlands, CA |
|
|
Field CB, Randerson JT, Malmström CM (1995). Global net primary production: Combining ecology and remote sensing. Remote. Sens. Environ 51:74-88. |
|
|
Folega F, Dourma M, Wala K, Batawila K, Xiuhai Z, Chunyu Z, Akpagana K (2014a). Basic Overview of Riparian Forest in Sudanian Savanna Ecosystem : Case Study of Togo. Revue d'écologie,Terre et Vie 69:24-38 |
|
|
Folega F, Dourma M, Wala K, Batawila K, Zhang CY, Zhao XH, Koffi A (2012). Assessment and impact of anthropogenic disturbances in protected areas of northern Togo. For. Stud. China 14:216-223 |
|
|
Folega F, Samake G, Zhang C-y, Zhao X-h, Wala K, Batawila K, Akpagana K (2011). Evaluation of agroforestry species in potential fallows of areas gazetted as protected areas in North-Togo. Afr. J. Agric. Res. 6:2828-2834 |
|
|
Folega F, Yao A, Woegan D, Marra K, Wala K, Batawila J, Leonardo S, Chun-yu Z, Dao-li P, ZhaoKoffi A (2015). Long term evaluation of green vegetation cover dynamic in the Atacora Mountain chain (Togo) and its relation to carbon sequestration in West Africa. J. Mt. Sci. 12:921-934 |
|
|
Folega F, Zhang C, Woegan YA, Wala K, Dourma M, Batawila K, Seburanga JL, Zhao X, Akpagana K (2014b). Structure and Ecology of Forest Plant Community in Togo. J. Trop. For. Sci. 26:225-239 |
|
|
Folega F, Chun-yu Z, Xiu-hai Z, Kperkouma W, Komlan B, Hua-guo H, Marra D, Koffi A (2014c). Satellite monitoring of land-use and land-cover changes in northern Togo protected areas. J. For. Res. 25:385-392 |
|
|
Foley C, Pettorelli N, Foley L (2008). Severe drought and calf survival in elephants. Biol Lett 4:541-544. |
|
|
Hien M (2001). Etude des déplacements des éléphants, lien avec leur alimentation et la disponibilité alimentaire dans le Ranch de Gibier de Nazinga, Province de Nahouri, Burkina Faso. Thèse de Doctorat, Université de Ouagadougou. |
|
|
Hoekstra JM, Boucher TM, Ricketts TH, Roberts C (2005). Confronting a biome crisis: global disparities of habitat loss and protection. Ecol. Lett. 8:23-29 |
|
|
IUCN (2008). Evaluation de l'efficacité de la gestion des aires protégées :aires protégées du Togo. Programme Afrique Centrale et Occidentale (PACO), Johnson NL, Kotz S (1970). Continuous Univariate Distributions: Distributions in Statistics vol 1. Hougton Mifflin. |
|
|
Jones T, Bamford AJ, Ferrol-Schulte D, Hieronimo P, McWilliam N, Rovero F (2012). Vanishing wildlife corridors and options for restoration: a case study from Tanzania. Trop. Conserv. Sci. 5:463-474 |
|
|
Kassa BD, Fandohan B, Azihou AF, Assogbadjo AE, Oduor AMO, Kidjo FC, Babatoundé S, Liu J, Glèlè Kakaï R (2013). Survey of Loxodonta africana (Elephantidae)-caused bark injury on Adansonia digitata (Malavaceae) within Pendjari Biosphere Reserve, Benin. Afr. J. Ecol. 52:385-394 |
|
|
Kokou K, Nuto Y, Atsri H (2009). Impact of charcoal production on woody plant species in West Africa: A case study in Togo. Sci. Res. Essays 4:881-893 |
|
|
Lovejoy TE, Hannah LJ (2005). Climate change and biodiversity. Yale University Press, Ann Arbor, Michigan |
|
|
Magurran AE (2004). Measuring Biological Diversity. Blackwell Science Ltd, Morrison ML, Marcot BG, Mannan RW (2006). Wildlife-habitat relationships : concepts and applications. Island Press, Washington, DC. |
|
|
McComb K, Moss C, Durant SM, Baker L, Sayialel S (2001). Matriarchs As Repositories of Social Knowledge in African Elephants. Science 292:491-494 |
|
|
Natta AK, Sinsin B, Van Der Maesen LJG (2004). A phytosociological study of riparian forests in Benin (West Africa). Belg. J. Bot. 136:109-128 |
|
|
Padakale E, Atakpama W, Dourma M, Dimobe K, Wala K, Guelly KA, Akpagana K (2015). Woody Species Diversity and Structure of Parkia biglobosa Jacq. Dong Parklands in the Sudanian Zone of Togo (West Africa). Annu. Res. Rev. Biol. 6:103-114. |
|
|
Parren MPE, Sam MK (2003). Elephant corridor creation and local livelihood improvement in West Africa. Paper presented at the International Conference on Rural Livelihoods, Forests and Biodiversity, Bonn, Germany, 19-23 May. |
|
|
Polo-Akpisso A, Wala K, Soulemane O, Folega F, Tano Y (2016). Changes in Land Cover Categories within Oti-Kéran-Mandouri (OKM) Complex in Togo (West Africa) between 1987 and 2013. In: Leal WF, Adamson K, Dunk RM, Azeiteiro UM, Illingworth S, Alves F (eds) |
|
|
Implementing Climate Change Adaptation in Cities and Communities: Integrating Strategies and Educational Approaches. Climate Change Management, 1 ed. Springer International Publishing. pp. 3-21. |
|
|
Prince SD, Goward SN (1995). Global Primary Production: A Remote Sensing Approach. J. Biogeogr. 22:815-835 |
|
|
RGPH (2011) Recensement Général de la Population et de l'Habitat. DGSCN. |
|
|
Rouse JJ, Haas R, Schell J, Deering D (1974). Monitoring vegetation systems in the Great Plains with ERTS. NASA Special Publication 351:309 |
|
|
Salako VK, Azihou AF, Assogbadjo AE, Houéhanou TD, Kassa BD, Glèlè Kakaï RL (2015). Elephant-induced damage drives spatial isolation of the dioecious palm Borassus aethiopum Mart. (Arecaceae) in the Pendjari National Park, Benin. Afr. J. Ecol. 1-11 |
|
|
Sebogo L, Barnes RFW (2003). Action plan for the management of transfrontier elephant conservation corridors in West Africa. IUCN/SSC/AfESG, West Africa Office, Tehou AC, Kossou E, Mensah GA, Houinato M, Sinsin B (2012). Identification et caractérisation des formations végétales exploitées par l'éléphant Loxodonta africana dans la Réserve de Biosphère de la Pendjari au Nord-Ouest de la République du Bénin. Pachyderm 52:36-48 |
|
|
Simberloff D, Cox J (1987). Consequences and Costs of Conservation Corridors. Conserv. Biol. 1:63-71 |
|
|
Tehou AC, Sinsin B (2000). Écologie de la population d'éléphants (Loxodonta africana) de la Zone Cynégétique de Djona (Bénin). Mammalia 64:29-40 |
|
|
UICN (2009) Baseline Study B : Etat actuel de la recherche et de la compréhension des liens entre le changement climatique, les aires protégées et les communautés, Projet : Evolution des systèmes d'aires protégées au regard des conditions climatiques, institutionnelles, sociales, et économiques en Afrique de l'Ouest, rapport final. GEF/UNEP/WCMC/UICN. |
|
|
Van Horne B, Wiens JA (2015). Managing habitats in a changing world. In: Morrison ML, Mathewson HA (eds) Wildlife habitat conservation: Concepts, challenges, and solutions. Wildlife management and conservation. Johns Hopkins University Press, Baltimore, Maryland. pp. 34-43 |
|
|
Vasilijević M, Zunckel K, McKinney M, Erg B, Schoon M, Rosen Michel T (2015). Transboundary Conservation: A systematic and Integrated Approach. IUCN, Gland, Switzerland. doi:10.2305/IUCN.CH.2015.PAG.23.en. |
|
|
Vavrek MJ (2011). Fossil: palaeoecological and palaeogeographical analysis tools. |
|
|
Vlek PLG, Le QB, Temene L (2010). Assessment of land degradation, its possible causes and threat to food security in Sub-Saharan Africa. In: Lal R, Stewart BA (eds) Food security and soil quality. Taylor & Francis Group, Boca Raton, Florida. pp. 57-86 |
|
|
Vogt P (2016) GuidosToolbox (Graphical User Interface for the Description of image Objects and their Shapes): Digital image analysis software collection 2.5 ed. |
|
|
Vogt P, Riitters KH, Estreguil C, Kozak J, Wade TG, Wickham JD (2006). Mapping Spatial Patterns with Morphological Image Processing. Landscape Ecol. 22:171-177 |
|
|
Wiens JA, Rotenberry JT (1985). Response of breeding passerine birds to rangeland alteration in a North American shrubsteppe locality. J. Appl. Ecol.:655-668. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


