Full Length Research Paper

ABSTRACT
Anthropogenic disturbances cause direct and indirect effects on the global decline in biodiversity. For better planning strategies on the conservation of medium and large mammals in Oaxaca, we present an analysis of daily activity patterns of medium and large mammals and their relative abundance in a Communal Natural Protected Area (CNPA) on the central coast of Oaxaca, Mexico. Sampling was carried out between November 2014 and September 2015, during the dry season and the rainy season, using forty camera traps placed within the CNPA El Gavilán and installed along wildlife trails, especially on pathways with clear evidence of use by wildlife. Date records for 10 species of medium and large mammals, obtained with 12,160 day/camera traps. Leopardus pardalis was active during the night but exhibited diurnal and nocturnal tendencies. Herpailurus yagouaroundi, Nasua narica, Dicotyles angulatus and O. virginianus were defined as cathemeral species. The most abundant medium mammals were Dasypus novemcinctus (RAI= 1.23), Didelphis virginiana (RAI= 1.15) and Nasua narica (RAI= 1.05). Our results can provide insights for the conservation of species in the CNPA El Gavilan. We recommend the continuance of studies on the temporal and seasonal variations of the activity patterns in order to maintain mammalian species conservation.
Key words: Conservation, relative abundance, mammals, Oaxaca, patterns.
INTRODUCTION
MATERIALS AND METHODS

RESULTS
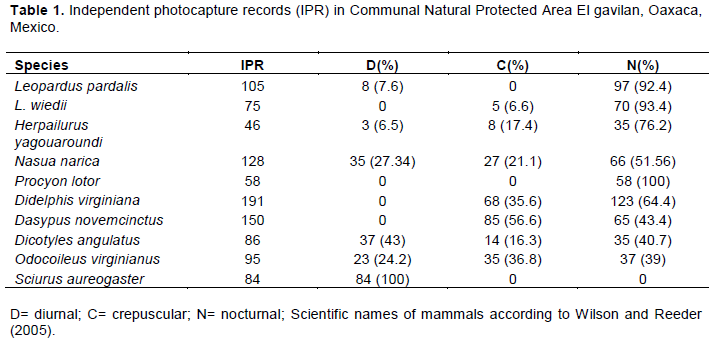
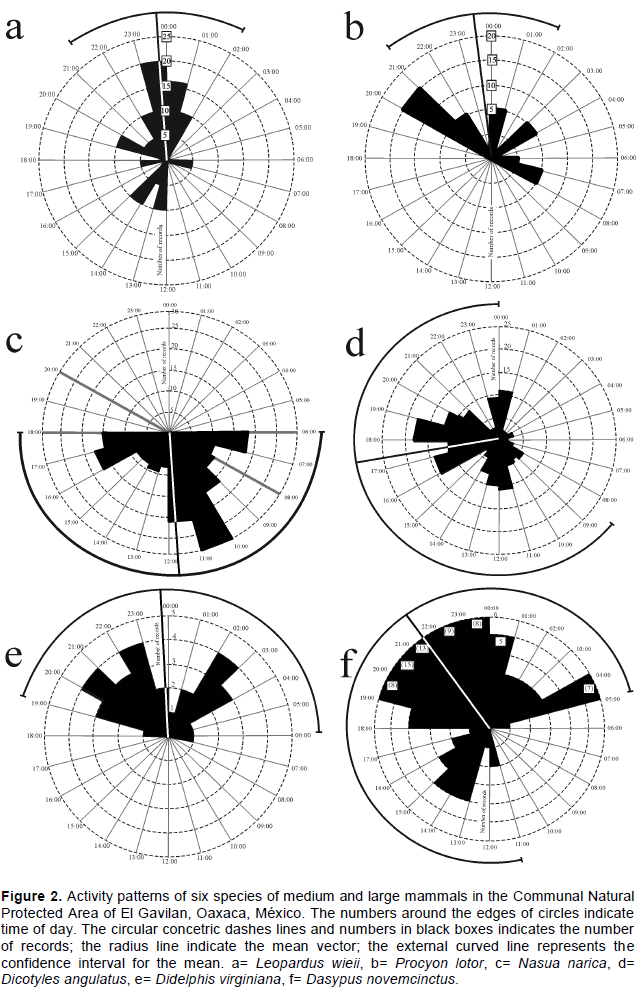



DISCUSSION
CONCLUSIONS
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Aranda M (2005). Leopardus pardalis. In: Ceballos G, Oliva G (eds.), Los mamíferos silvestres de México. México, DF: Fondo de Cultura Económica, Comisión Nacional para el Conocimiento y uso de la Biodiversidad. pp. 359-361. |
|
|
Aranda M (2012). Manual for tracking wild mammals in Mexico. National Commission for the Knowledge and Use of Biodiversity (CONABIO). Mexico DF. |
|
|
Baboo B, Sagar R, Bargali SS, Verma H (2017). Tree species composition, regeneration and diversity within the protected area of Indian dry tropical forest. Tropical Ecology 58(3):409-423. |
|
|
Bargali SS, Singh RP, Singh SP (1992b). Structure and function of an age series of eucalypt plantations in Central Himalaya, II. Nutrient dynamics. Annals of Botany 69:413-421. |
|
|
Bargali SS, Singh SP, Singh RP (1992a). Structure and function of an age series of eucalypt plantations in Central Himalaya, I. Dry matter dynamics. Annals of Botany 69:405-411. |
|
|
Botello F, Sánchez-Cordero V, Ceballos G (2008). Diversidad de carnívoros en Santa Catarina Ixtepeji, Sierra Madre de Oaxaca, México. In. Lorenzo CE, Espinoza J, Ortega J (eds), Avances en el estudio de los mamíferos de México II. México, DF: Asociación Mexicana de Mastozoología, A. C., pp. 335-354. |
|
|
Bradley J, Cardinale J, Emmett D, Gonzalez A, Hooper DU., Perrings C, Venail P, Narwani A, Mace GM, Tilmanx D, Wardle DA, Kinzig AP, Daily G, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012). Biodiversity loss and its impact on humanity. Nature 7401:59-67. |
|
|
Briones-Salas MA, Martín-Regalado N, Lavariega MC (2016). Mammals in tropical dry forest on the central coast of Oaxaca, Mexico. Check List 12(2):1862. |
|
|
Briones-Salas MA, Sánchez-Cordero V (2004). Mamíferos. In. García Mendoza AJ, Ordoñez MJ, Briones-Salas MA (eds.), Biodiversidad de Oaxaca. México, DF: Instituto de Biología, UNAM, Fondo Oaxaqueño para la Conservación de la Naturaleza-World Wildlife Fund, México, pp. 423-447. |
|
|
Buenrostro Silva A, Sigüenza Pérez D, García Grajales J (2015). Mamíferos carnívoros del Parque Nacional de Chacahua, Oaxaca: Riqueza, abundancia y patrones de actividad. Revista Mexicana de Mastozoología Nueva Época 5(2):39-54. |
|
|
Bustamante A (2008). Densidad y uso de hábitat por los felinos en la parte sureste del área de amortiguamiento del Parque Nacional Corcovado, Península de Osa, Costa Rica. Tesis de maestría. Instituto Internacional en Conservación y Manejo de Vida Silvestre, Universidad Nacional, Costa Rica. |
|
|
Cardinale BJ, Duffy EJ, Gonzalez A, Hooper D, Perrings C, Venail P, Narwani A, Mace G, Tilman D, Wardle DA, Kinzig A, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012). Biodiversity loss and its impact on humanity. Nature 486:59-67. |
|
|
Carvajal-Villarreal S, Caso A, Downey P, Moreno A, Tewes ME, Gassman LI (2012). Spatial patterns of the margay (Leopardus wiedii, Falidae, Carnivora) at El Cielo Biosphere Reserve, Tamaulipas, Mexico. Mammalia 76:237-244. |
|
|
Cortés-Marcial M, Briones-Salas MA (2014). Diversidad, abundancia relativa y patrones de actividad de mamíferos medianos y grandes en una selva seca del Istmo de Tehuantepec, Oaxaca, México. Revista de Biología Tropical 62(4):1433-1448. |
|
|
Chiarello AG (2000). Density znd population size of mammals in remants of Brazilian Atlantic Forest. Conservation Biology 14(6):1469-1657. |
|
|
De Oliveira TG (1998). Leopardus wiedii. Mammalian Species 579:1-6. |
|
|
Erkert HG, Bay FA, Kracht S (1976). Zeitgeber induced modulation of activity patterns in nocturnal mammals (Chiroptera). Experientia 32(5):560-562. |
|
|
Frey S, Fisher JT, Burton AC, Volpe JP (2017). Investigating animal activity and temporal niche partitioning using camera-trap data: Challenges and opportunities. Remote Sensing in Ecology and Conservation 3(3):123-132. |
|
|
Gallina S, Bello J (2014). Patrones de actividad del venado cola blanca en el noreste de México. Therya 5(2):423-436. |
|
|
García E (1973). Los climas de México. México, DF: Instituto de Geografía, Universidad Nacional Autónoma de México. |
|
|
Gódinez GA (2014). Patrones de actividad espacio-temporal de ungulados de la Rserva de la Biósfera El Triunfo, Chiapas, México. Tesis de licenciatura, Universidad Michoacana de San Nicolás de Hidalgo, Michoacán, México. |
|
|
Gonthier DJ, Castañeda FE (2013). Large and medium sized mammal survey using camera traps in the Sikre River in the Río Plátano Biosphere Reserve, Honduras. Tropical Conservation Science 6(4):584-591. |
|
|
Gonzalez-Christen A (2010). Los mamíferos de Veracruz: Distribución, endemismo y estado de conservación. México, DF: Comisión Nacional para el Conocimiento y Uso de la Biodioversidad (CONABIO), Gobierno del Estado de Veracruz, Universidad Veracruzana, Instituto de Ecología, A. C. |
|
|
González-Pérez G, Briones Salas MA, Alfaro AM (2004). Integración del conocimiento faunístico de Oaxaca. In: García Mendoza AJ, Ordóñez MJ, Briones Salas MA (eds.), Biodiversidad de Oaxaca. México, DF: Instituto de Biología, Universidad Nacional Autónoma de México, Fondo Oaxaqueño para la Conservación de la Naturaleza, World Wildlife Fund, México, pp. 449-466. |
|
|
Gosain BG, Negi GCS, Dhyani PP, Bargali SS, Saxena R (2015). Ecosystem services of forests: Carbon Stock in vegetation and soil components in a watershed of Kumaun Himalaya, India. International Journal of Ecology and Environmental Science 41(3-4):177-188. |
|
|
Heurich M, Hilger A, Küchenhoff H, Andrén H, Bufka L, Krofel M, Mattison J, Persson J, Rauset GR, Scmidt K, Linell JDC (2014). |
|
|
Activity patterns of Eurasian lynx are modulated by light regime and individual traits over a wide latitudinal range. PLoS ONE 9(12):e114143. |
|
|
Karanth K, Nichols D (1998). Estimation of tiger densities in India using photographic captures and recaptures. Ecology 79:2852-2862. |
|
|
Kovach WL (2011). Oriana-Circular Statistics for Windows. V4. Kovach Computing Services, Pentraeth, Wales, U.K. |
|
|
Kronfeld-Schor N, Dayan T (2003). Partitioning of time as an ecological resource. Annual Review of Ecology, Evolution and Systematics 34:153-181. |
|
|
Lehner PN (1996). Handbook of ethological methods. Second edition, Cambridge, United Kingdon: Cambdridge University Press. |
|
|
Linkie M, Ridout MS (2011). Assessing tiger-prey interactions in Sumatran rainforests. Journal of Zoology 284(3):224-229. |
|
|
Long ES, Nelson TC, Steensma KMM (2013). Conditional daily and seasonal movement strategies of male Columbia black-tailed deer (Odocoileus hemionus columbianus). Canadian Journal of Zoology 91:679-688. |
|
|
Mandujano S, Hernández C (2019). Use of water developments by White-tailed deer in an extensive AHU in the biosphere reserve Tehucan-Cuicatlan, Mexico. Agroproductividad 12(6):37-42. |
|
|
Manfo DA (2013). Agroforestry and dynamics of floristic biodiversity and the agricultural landscape in the forest-savanna transition zone: the case of Obala region, Center-Cameroon. Master thesis in Geography, University of Yaoundé. |
|
|
Marques MD (2004). Rhythms and ecology- do chronobiologists still remember nature? Biological Rhythm Research 35:1-2. |
|
|
Marques RV, Fabián ME (2018). Daily activity patterns of medium and large neotropical mammals in an area of Atlantic rain forest at altitude. Revista Brasileira de Zoociências 19(3):38-64. |
|
|
Mesa-Zavala E, Álvarez-Cárdenas S, Gallina-Tessaro P, Troyo-Diéguez E, Guerrero-Cárdenas I (2012). Vertebrados terrestres registrados medianto fototrampeo en arroyos estacionales y cañadas con agua superficial en un hábitat semiárido de Baja California Sur, México. Revista Mexicana de Biodiversidad 83(1):235-245. |
|
|
Monroy-Vilchis O, Zarco-González M, Rodríguez-Soto L, Urios V (2011). Fototrampeo de mamíferos en la Sierra de Nanchititla, México: abundancia relativa y patrón de actividad. Revista de Biología Tropical 59:373-383. |
|
|
Morgan E (2004). Ecological significance of biological clocks. Biological Rhythm Research 35(1/2):3-12. |
|
|
Mourya NR, Bargali K, Bargali SS (2019). Effect of Coriaria nepalensis Wall. colonization in a mixed conifer forest of Indian Central Himalaya. Journal of Forestry Research 30(1):305-317. |
|
|
O'Connell AF, Nichols JD, Karanth KU (2011). Camera traps in animal ecology: methods and analyses. Tokyo, Japan: Springer Press. |
|
|
Ogurtsov S, Zheltukhin A, Kotlov IP (2018). Daily activity patterns of large and medium mammals based on camera traps data in the Central Forest Nature Reserve, Valdai Upland, Russia. Nature Conservation Research 3(2):68-88. |
|
|
Ortiz-Pérez MA, Hernández JR, Figueroa JM (2004). Reconocimiento fisiográfico y geomorfológico. In: García Mendoza AJ, Ordóñez MJ, Briones Salas MA (eds.), Biodiversidad de Oaxaca. México, DF: Instituto de Biología, Universidad Nacional Autónoma de México, Fondo Oaxaqueño para la Conservación de la Naturaleza. World Wildlife Fund. pp. 43-54. |
|
|
Pérez-Irineo G, Santos Moreno A (2012). Diversidad de mamíferos terrestres de talla grande y media de una selva subcaducifolia del noreste de Oaxaca, México. Revista Mexicana de Biodiversidad 83:164-169. |
|
|
Pérez-Irineo G, Santos-Moreno A (2016). Abundance and activity patterns of médium-sized felids (Felidae, Carnivora) in Southeastern Mexico. The Southwestern Naturalist 61(1):33-39. |
|
|
Pina GPL, Gómez RAC, González CAL (2004). Distribution, habitat association and activity patterns of médium and large sized mammals of Sonora, Mexico. Natural Areas Journal 24:354-357. |
|
|
Rodríguez-Luna E, Gómez Pompa A, López-Acosta JC, Velázquez-Rosas N, Aguilar-Domínguez Y, Vázquez-Torres M (2011). Atlas de los espacios naturales protegidos de Veracruz. Gobierno del estado de Veracruz. Veracruz, México: Universidad Veracruzana. |
|
|
Salas-Morales SH, Saynes A, Schibli L (2003). Flora de la costa de Oaxaca, México: lista florística de la región de Zimatán. Boletín de la Sociedad Botánica de México 72:21-58. |
|
|
Sunquist M, Sunquist F (2002). Wild coast of the world. Chicago, USA: Chicago University Press. |
|
|
Tobler MW, Carrillo-Percastegui S, Leite R, Mares R, Powell G (2008). An evaluation of camera traps for inventorying large- and médium-sized mammals. Animal Conservation Journal 11:169-178. |
|
|
Torres-Colín R (2004). Tipos de vegetación. In. García Mendoza AJ, Ordóñez MJ, Briones Salas MA (eds.), Biodiversidad de Oaxaca. México, DF: Instituto de Biología, Universidad Nacional Autónoma de México, Fondo Oaxaqueño para la Conservación de la Naturaleza, World Wildlife Fund, pp. 105-117. |
|
|
Trejo I (1998). Distribución y diversidad de selvas bajas de México: relación con clima y suelo.Tesis de Doctorado, Universidad Nacional Autónoma de México, México. |
|
|
Trejo I (2010). Las selvas secas del Pacífico mexicano. In: Ceballos G, Martínez L, García A, Espinoza E, Bezaury J, Dirzo R (eds.), Diversidad, amenazas y áreas naturales prioritarias para la conservación de las selvas secas del Pacífico de México, México. México, DF: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad, Fondo de Cultura Económica, México, pp. 4-51. |
|
|
Valdéz M, Téllez G (2005). Sciurus aureogaster F. Cuvier, 1829. In. Ceballos G, Oliva G, (eds.), Los mamíferos de México. México, DF: Fondo de Cultura Económica, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. pp. 547-548. |
|
|
Van Schaik CP, Griffiths M (1996). Activity periods of Indonesian Rain Forest Mammals. Biotropica 28:105-112. |
|
|
Vera MR, Fábian ME (2018). Daily activity patterns of médium and large Neotropical mammals during different seasons in an area of huigh altitude atlantic rain forest in the South of Brazil. Zoociencias 19(3): 38-64. |
|
|
Valenzuela-Galván D, De León-Ibarra A, Lavalle-Sánchez A, Orozco-Lugo L, Chávez C (2013). The margay Leopardus wiedii and bobcat Lynx rufus from the dry forests of Southern Morelos, Morelos. Southwestern Naturalist 58:118-120. |
|
|
Zar JH (2010). Biostatistical analysis. Prentice Hall, Upper Saddle River, New Jersey, USA. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


