ABSTRACT
Three morphotypes A, B and C were isolated after description of Cyclops abyssorum mauritaniae populations obtained from 46 Algerian water bodies. These are described based on their morphometric characters. Morphotype A found in six localities characterized the east of Algeria, morphotype B was collected in the west of Algeria, while morphotype C was collected in High Mountain in the North. After dissection of certain males and females, measurements of the different parts of the body illustrated by drawings were made for each morphotype. The size of morphotype B is intermediate between A and C. The outer side of the second article of the fifth thoracic leg presents a small bump only in morphotype A and C. Morphotype C is characterized by the convexity of its third, fourth and fifth thoracics anterior margin. Other differences were observed on ornamentations of the antenna basis and the coxa of the fourth leg. This subspecies, with common occurrence in the Eastern Region of Algeria and rare occurrence in the northern and western parts of the country indicates that it is dispersed in Algeria from the East to the West and North.
Key words: Algeria, Copepoda, Cyclops abyssorum mauritaniae, distribution, polymorphism.
Cyclops abyssorum mauritania Lindberg 1950 is a Southern element of the Holarctic genus Cyclops O.F.Muller 1776. In North Africa, C. abyssorum mauritaniae was first reported from Morocco by Lindberg (1950) and was met for the first time in Algeria in 1992 (Akli, 1992). In Tunisia it was reported later by Toumi et al. (2013). Three populations from the Middle and High Atlas (Dayat Ifrah, Aguelmane de Sidi Ali, and Lake Ifni) have been compared (Dumont and Decraemer, eutrophication represents a recent dispersion caused by 1977).
In accordance with its eurytopy, the species has been recorded as tolerating a variety of environmental conditions, such as acidic waters (Røen, 1962) and alkaline waters (Morgan, 1972). In Algeria shallow dams, this species was usually observed after eutrophication (Bidi et al., 2014). According to Krajicek et al. (2016), man. Many morphotypes of C. abyssorum have been described by different authors. Einsle (1975) found that the planktonic populations react quickly to the environmental conditions changes with a change of morphometry of the different parts of the body.
Sampling
A total of 46 water bodies (Table 1 and Figure 1) were sampled in different parts of Algeria with one sampling per water body except for three localities (9, 10 and 18). Samples were collected using plankton net of 50µm mesh size. The samples were fixed in 5% formalin solution. Samples were collected by the author.
Dissection
For preparation, the organisms were first placed in small dishes containing a mixture of formalin water and glycerine. The water evaporated in 1-2 days and then specimens were dissected in the concentrated glycerine. The cyclopoids were examined in dorsal view. Total body length and length of antennules in relation to the cephalothorax were measured. Other measurements on the caudal rami, the fourth and fifth thoracic legs were made after dissection and followed by drawings for each morphotype of C. abyssorum mauritaniae. Cyclopoids were identified using the key of Dussart (1969) (Table 2).
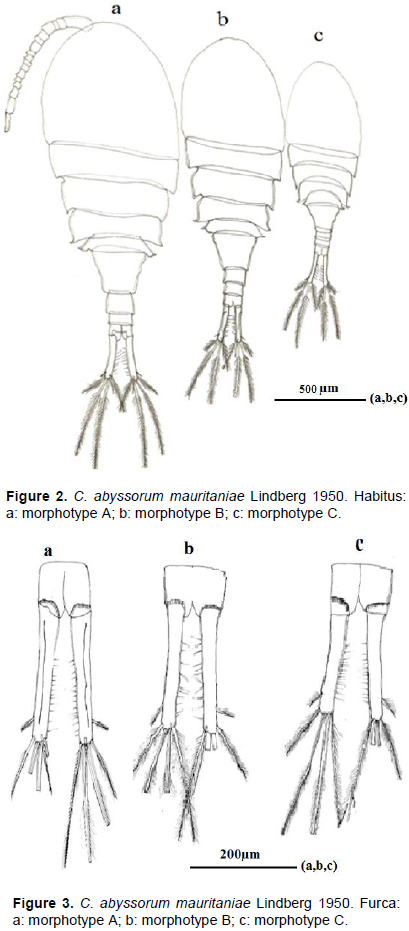
Three Algerian morphotypes A, B and C of C. abyssorum mauritaniae were identified. The morphotype A which characterises the East Algeria water bodies was the most abundant, collected at different altitudes (25, 600 and 1090 m) in lakes and wadis. Only two females of morphotype B was sampled in West Algeria, in wadi at 86 m altitude, while morphotype C was collected in lake in the High Mountain (1200 m altitude), in the North. According to Holynska and Wyngaard (2019), C. abyssorum G. O Sars, 1863 occur in both low and highâ€altitude habitats. Morphotype A (Figure 2a) is the most robust one (2.2-2.3 mm); size of morphotype B (1.9- 2.1 mm) is intermediate between A and C (1.7-2 mm) (Figure 2b and c). Tunisian (1.6-2.2 mm) and Algerian specimens of C. abyssorum mauritaniae have relatively big size compared to Morocco specimens of Aguelmane de Sidi Ali (1.4-1.74 mm) and lake Ifni (1.24-1.29 mm). From Dayat Ifrah one adult female was isolated, suggesting a more robust form than the animals from Aguelmane de Sidi Ali, with shorter furcal rami (Dumont and Decraemer, 1977). Algerian specimens’ first antenna reaches the posterior margin of the cephalothorax; those from Morocco have the first antenna that reaches the half of the second thoracic segment; those from Tunisian exceed it. Morphotype C shows a habitus that is close to the material of Aguelmane Sidi Ali with convexity of its third, fourth and fifth thoracic anterior margin (Figure 2c). Furcal rami in morphotype C (Figure 3c) are slightly shorter (5.6 -6.9 times as long as wide) than of morphotype A and B (6 -7 times as long as wide) (Figure 3a, b). Morphotype A shows furcal rami (Figure 3a) with thickened ridge in the dorsal side which is close to that of Dumont material of Dayat Ifrah (Dumont and Decraemer, 1977).
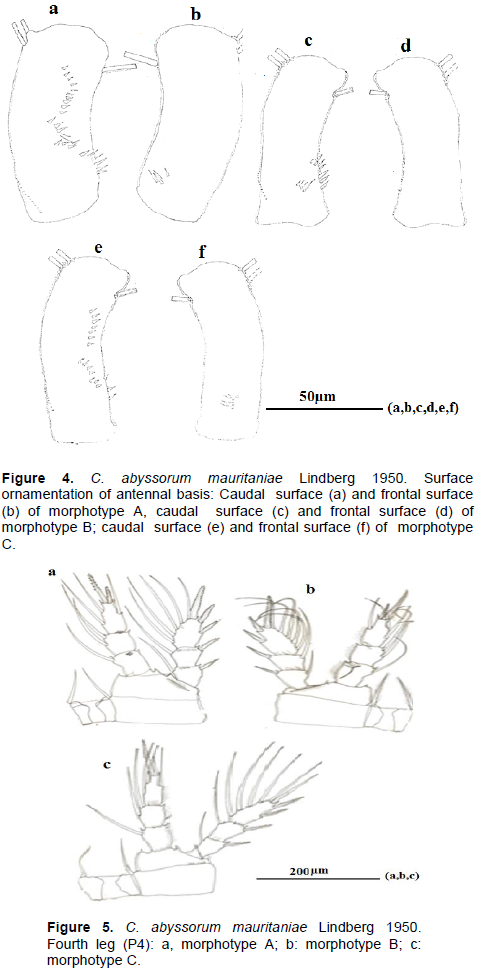
Differences between these three morphotypes were observed in spinule ornamentation of the antennary basipodite. T the antennary basipodite in caudal view (Figure 4a, c and e) is shown in both morphotypes’ proximal spinules; on the lateral rim, it is composed of 6 long spinules slightly incurved in morphotype A, 8 spinules in morphotype B and 3 little spinules in morphotype C. Oblique row of spinules next to proximal row on lateral rim is composed of 7 elongated spinules in morphotype A, 4 spinules in morphotype B and 7 spinules in morphotype C. Longitudinal row along lateral rim is arranged in one curved row of small spinules (10 spinules) of equal size in morphotype A and one continuous row of spinules (7spinules) in morphotype C, in morphotype B, longitudinal row along lateral rim is absent. Frontal spinule pattern of antennary (Figure 4b, f) consists of group of oblique spinules (4) near the base in morphotype A and C. frontal basipodite antenna without spinules (3d).
In all morphotypes, formula of exopodite 3 (P1-P4) is: 3- 4- 3- 3; the endopodite 3 of the fourth leg is 2 to 2.5 times as long as broad. The internal apical spine is 2 times as long as the external apical spine (Figure 5a, b and c). Caudal spinule ornamentation of P4 coxopodite (Figure 6a, b and c) composed of intermittent row of spinules along distal rim is arranged differently in both morphotypes; (4+5) in morphotype A, (11+1) in morphotype B and (4+4) in morphotype C. Along proximal rim of P4 coxopodite, spinules are arranged in two rows with different size, consisting of 17 spinules in morphotype A, 22 spinules in morphotype B and 16 spinules in morphotype C. On lateral rim, a row of numerous fine spicule comb like exists only in morphotype A. The connecting plate of P4 carries two rows of long hair; it is an ornamentation which is identical to all the morphotypes. The outer side of the second article of the fifth thoracic leg presents a small bump in morphotype A and C (Figure 7a and c) with 4 to 5 small spines for morphotype A (Figure 7a) and 3 to 4 small spines for morphotypes B and C (Figure 7b and c).


In the 46 water bodies sampled, C. abyssorum mauritaniae existed only in eight localities with considerable morphological plasticity. Morphological characters of morphotype A are stable in all its localities; for morphotype C, any morphological differences in samples taken at different time were found. These suggest that morphotypes character differences are not local neither temporal, and are due to fragmented dispersion of C. abyssorum mauritaniae. This subspecies is well represented in the Eastern Region of Algeria and rare in the North and West regions, indicating that it originated in the East and began spreading to north and west Algeria taking different forms. C. abyssorum mauritaniae is one of the relatively poorly known representatives of the Holarctic genus Cyclops. A study on the geographic variation of the morphological characters in this North African taxon might have been a very interesting.
The authors have not declared any conflict of interests.
The authors are grateful to Professor H. DUMONT for welcoming them in his laboratory and to Dr. S. MAAS for her availability and helping to confirm the species copepods.
REFERENCES
|
Akli S (1992). Systematic and geographical repartition of fresh water copepods (Crustacean) in the North of Algeria. Thesis. University of Sciences and Technology Houari Boumediene. Faculty of Biological Sciences. Algiers, p. 156.
|
|
|
|
Bidi AS, Arab A, Samraoui B (2014).Variation spatio-temporelle du zooplancton dans le barrage de la réserve de chasse de zéralda (Algérie). Revue d'Écologie (Terre et Vie) 69(3-4):214-224.
|
|
|
|
|
Dumont HJ, Decraemer W (1977). On the continental copepod fauna of Morocco. Hydrobiologia 52(2-3):257-278.
Crossref
|
|
|
|
|
Dussart BH (1969). Les Copépodes des eaux continentales d'Europe Occidentale. 2: Cyclopoïdes et Biologie quantitative. Boubée & Cie, Paris, p. 292.
|
|
|
|
|
Einsle U (1975). Revision der Gattung Cyclops s. str., speziell der abyssorum-Gruppe Memorie dell'Istituto Italiano di Idrobiologia dott. Marco De Marchi 32:57-219.
|
|
|
|
|
Holynska M, Wyngaard GA (2019). Towards a phylogeny of Cyclops (Copepoda): (in) congruences among morphology, molecules and zoogeography. Zoologica Scripta 48(3):376-398.
Crossref
|
|
|
|
|
Krajicek M, Fott J, Miracle MR, Ventura M, Sommaruga R, Kirschner P ÄŒerny M, (2016). The genus Cyclops (Copepoda, Cyclopoida) in Europe. Zoologica Scripta 45(6):671-682.
Crossref
|
|
|
|
|
Lindberg K (1950). Cyclopides (Crustaces, Copepodes) du Maroc. Bulletin de la Société Des Sciences Naturelles 30:23-32.
|
|
|
|
|
Morgan NC (1972). Productivity studies at Loch Leven (a shallow nutrient rich lowland lake). Proc. IBP UNESCO Symposium on Productivity Problems of Freshwaters, Kazimierz Dolny pp. 183-205.
|
|
|
|
|
Toumi H, Bejaoui M, Boumaiza M (2013). Contribution to the ecological study of epigean Cladocera and Copepoda (Cyclopoida) from groundwater in Northern Tunisia. Journal of Nature and Technology. C. Sciences Environnementales 5(1):12-18.
|
|
|
|
|
Røen U (1962). Studies on freshwater Entomostraca in Greenland II: Localities, ecology, and geographical distribution of the species. Greenland Me (2):170dd.
|
|





