ABSTRACT
We investigated how four primate species in Kakum Conservation Area (KCA) coexisted after logging shrunk their feeding resources. We used multivariate discriminant function analysis to determine whether feeding sites and food types could discriminate them. Results showed that lower canopy discriminated Cercopithecus mona and Cercopithecus petaurista; middle canopy, C. petaurista and Procolobus verus; upper canopy P. verus and Colobus polykomos; seed C. polykomos; fruit, P. verus. Variations in seasonal and zonal selections of feeding sites and food types were significant at the 0.05 level. Primates selected upper canopy, flower and fruit more in rainy season (mean abundance ± se = 14.45±1.2, 10.21±0.53, 17.69±0.7) than in dry season (9.32±0.67, 8.11±0.52, 12.58±0.54); middle canopy and seed more in dry season (13.17±0.12 and 16.7±0.84) than rainy season (8.07±0.53 and 8.43±0.6), respectively. Upper and middle canopies and seed were more selected at the park’s centre (mean abundance ± se = 14.3±1.57, 12.8±1.06 and 16.83±1.16, respectively); and lower canopy, periphery (16.98±1.42). C. polykomos selected the park’s centre mostly (16.6±2.4); C. petaurista, inner (16.9±1.2); and C. mona, periphery (14.7±1.3). Selection by C. polykomos reduced from the park’s centre (mean abundance ± se = 16.6±2.4) through inner (10.5±0.2) to periphery (6.6±1.6); but selection by C. mona reduced from periphery (14.7±1.3) through inner (12.2±1.3) to centre (11.2±1.6). C. petaurista and P. verus appeared to be forest generalists. Seasonal and spatial variations, resource variability and forest conditions facilitated resource partitioning to allow co-existence. Strict measures are required at KCA to facilitate forest regeneration to conserve the primates.
Key words: Conservation, monkeys, resource-shrunk, co-existence, reserve, forest, regeneration.
An important phenomenon to ecological separation of sympatric animals is resource partitioning, which allows co-existing species to utilize similar resources. Many factors, including forage quality and quantity, habitat type, patches, feeding sites and the animal’s security influence resource selection by mammals (Bailey et al., 1996; Wallis de Vries et al., 1999). Decisions made by mammals at the levels of one or more of these factors account for their spatial distribution (Turchin, 1991). For their safety, mammals become increasingly confined to protected areas (Dakwa et al., 2016; Dakwa et al., 2014; Barnes, 1999; Newmark, 1996) as there is a growing human population pressure and land use change (Cincotta et al., 2000). Habitat disturbances and hunting have been identified as the main anthropogenic factors causing declines in primate populations (Cowlishaw and Dunbar, 2000; Oates, 1996; Fa et al., 1995). While some primate species, e.g. Cercopithecus petaurista (lesser white-nosed monkey), are unaffected by habitat disturbances such as logging (Dakwa, 2016; Martin and Asibey, 1979), others like the C. diana (Diana monkey) have suffered population declines (Martin and Asibey, 1979). Kakum Conservation Area (KCA) was established in Ghana in 1995 after a sustained logging removed commercial trees from a large area of the then forest reserve. This resulted in shrinking of resources and a disturbed area (Dakwa, 2016; WD, 1996). In general scientific research is scanty at KCA. Research into the behaviour of the animals, particularly, the arboreal species such as the primates, several years after their habitats and feeding resources were destroyed, could be very useful to provide information needed for management planning and action towards their conservation. We tried to find out how the different species of primates are able to co-exist at resource-shrunk KCA, by investigating: 1) the patterns of seasonal and spatial variations of feeding sites and food types of four primate species, Cercopithecus mona (Mona monkey), C. petaurista (lesser white-nosed monkey), Procolobus verus (olive colobus monkey) and Colobus polykomos (black-and-white colobus monkey), which inhabit the tree canopies of KCA, along a habitat disturbance gradient, and; 2) how six feeding resources, three feeding sites (upper, middle and lower canopies) and three food types (flower, fruit and seed) are partitioned to allow co-existence of the primate species. We tested the hypothesis that: 1) variability in feeding resources facilitates differential use of the resources, allowing the primate species to co-exist; and 2) feeding resources partitioned among primates could be influenced by seasonal and spatial variation of feeding sites and food types.
Many authors have documented primates’ use of plant parts as food, e.g. fruit (Chapman, 1989; Duc et al., 2009; Peres, 1994; Johns and Skorupa, 1987), flower (Gautier-Hion, 1970; Johns and Skorupa, 1987), seed (Johns and Skorupa, 1987), leaf (Duc et al., 2009) and gum (Gautier-Hion, 1970); but in this study we focused on fruit, flower and seed, because of the primates involved in the study.
Also C. diana (Diana monkey) was excluded from this investigation because they occur at a very low density and being difficult to sight at KCA; and the remaining two primate species, Periodictus potto (potto) and Galago crassicaudatus (bush baby) reported to occur at KCA (Dakwa, 2016) were excluded for being nocturnal, which is enough reason for ecological separation from others.
Study area
Kakum Conservation Area (Figure 1 ) is located in the Central Region of Ghana and protects about 360 sq.km of rain forest. The area had been selectively logged in the past, but its status changed from forest reserve to wildlife protected area in 1992 (WD, 1996); and attained the present status of Conservation Area in 1995. The average annual rainfall is about 1600 mm and the average relative humidity is about 80% throughout the year while temperature ranges from 18.2 to 32.1°C (FC, 2007). The KCA is surrounded by about 50 local communities with a population of about 40,000 people and farmlands sharing boundaries with the reserve (Monney et al., 2010). About 105 species of vascular plants (WD 1996) and 69 species of mammals including seven primate species (Dakwa, 2016; Yeboah, 1996) have been identified in KCA.
Data collection
We divided the study area into three zones, namely the periphery, the inner part and the centre, which were classified according to canopy coverage and light penetration (Wiafe et al., 2010). The periphery is the reserve’s margins, which suffered severest of disturbances through logging in the past. Trees at the periphery are mostly short and the reserve’s floor is exposed to > 75% sunlight. Canopies are not often the close type, being < 25% close. The centre is the most interior part of the reserve. It has the tallest trees, which form > 75% close canopies mostly, and light penetration to the floor is very low, < 25%. Logging did not affect this part of the reserve very much and so disturbances are minimal. Between the periphery and the centre is the inner part of the reserve. There are more tall trees and canopies are close, >50% but <75 %. Logging was reduced from the periphery to the centre and so disturbances at the inner part of the reserve are intermediate. Light penetration to the reserve floor is <50% but >25%. Thus there are gradients in respect of anthropogenic disturbances, canopy formation, and tree heights from the periphery to the centre, though each of these zones of the reserve has substantial number of upper, middle and lower canopies. Upper canopy refers to canopies occurring at heights above 45 m of a tree; middle canopy refers to canopies occurring from 30-45 m high and lower canopy, below 30 m. Canopy heights were estimated by using a laser range finder (Yardage Pro Compact 800, Bushnell factory, Overland, KA, USA).
The study relied on a field study of sampled plots laid in the three zones of KCA (Figure 2). In each zone, we established eight circular plots, each of 50 m radius, maintaining at least 300 m interval between plots (Figure 2). Thus, a total of 24 plots were established. We conducted the fieldwork over a period of 12 months from March, 2016 to April, 2017 between 6.30 - 9.00 GMT and 16.00 - 18.00 GMT each day, when the primates were feeding. Four groups of workers each comprising three individuals, giving a total of 12 people, made up of the researchers and volunteers, wildlife students of the University of Cape Coast and KCA field staff were involved in this investigation; and all were familiar with the identification of the monkeys. Each group investigated six plots (Figure 2) without overlapping with other groups. All 24 plots were investigated over the same five continuous days every month; thus there were two plots a day per group, one plot in the morning and the other in the evening. We always used the next day to deploy workers to get closer to plots. Each group worker took a portion of the plot and by viewing with Bushnell H2O Proof Prism Binocular 10 x 42-mm (Bushnell Corp, Overland, KA, USA) from hideouts, we observed the primates as they fed on flowers, fruits and seeds on the upper, middle and lower canopies. For each primate observation we recorded the following details; i) species name, ii) canopy type on which it was found, iii) the food type eaten, iv) the number of a particular food type eaten and v) the number of a particular feeding site on which it was found. We repeated this every month, ensuring that plots observed in the morning were observed in the evening the next month and plots observed in the evening were observed in the morning the next month.

Analyses of data
We used linear discriminant function analysis (LDA) (Gail et al., 2007; Quinn and Keough, 2002), a multivariate procedure, which discriminates between two or more naturally occurring groups, to determine whether groups (in this case, four primate species), could be discriminated on the basis of the primate’s feeding resources in this case, number of times at feeding sites, that is, upper canopy, middle canopy, lower canopy, and food types, that is, flower, fruit or seed, which were selected by the primates. First of all, the assumptions that: (1) population covariance matrices did not depend on the population from which the data were obtained; (2) there was no discrimination on any dimension; and (3) there was no overall group effect, were tested. In LDA, the contributions of the functions to the discrimination between groups do not overlap. Rather, LDA allows the first function to provide the most overall discrimination between groups, picking up the most variation; the second function provides the next most overall discrimination and picks up the most of the unexplained variation remaining (Gail et al., 2007; Quinn and Keough, 2002). We used the enter method to assign a predictor with only the unique association it has with the groups. Discriminant function scores as dependent variables, and primates as independent variables, were tested in a one-way ANOVA followed by a Tukey’s HSD that was used to test if groups differed significantly. SPSS (Version 17) software was used in all analyses.
Feeding sites and food types selected by primates
Table 1 displays the mean abundance and standard errors of primates, from all the 384 observations, that selected the various feeding sites and food types at Kakum Conservation Area during the study. Out of the six feeding sites and food types studied, fruits were the most selected by the primates, reaching mean abundance (± se) of 15.14±0.46, followed by the lower canopy (14.14±0.82), and flowers (9.16±0.37) being the least (Table 1). C. mona and C. petaurista selected the lower canopy mostly but C. petaurista (mean abundance ± se = 20.14±1.5) selected a little more than the C. mona (mean abundance ± se =18.79±1.6). C. polykomos and P. verus were more inclined to the upper canopy reaching high means of abundance (± se) of 18.35±2.0 and 15.35±1.2, respectively; but the selection of fruits by P. verus reached the highest mean of abundance ± se = 17.63±0.86 of all the primates studied. C. polykomos (mean of abundance ± se = 15.22±1.4) selected more seeds than other primates, while C. petaurista selected more flowers than other primates (mean of abundance ± se = 12.41±0.84). The middle canopy was selected by P. verus more than other primates reaching the highest mean of abundance ± se of 12.90±0.98, followed by C. mona (mean of abundance ± se = 12.52±0.96).
Primates’ selection of the canopies significantly and positively correlated with their selection of flowers, fruits and seeds except the upper canopy which correlated negatively and not significantly with flowers (Table 2). Primates’ selection of the upper canopy negatively correlated with their selection of the lower canopy. Flower selection correlated significantly and positively with fruit but negatively with seed (Table 2).
Discriminant analysis
Box’s M test was significant (Box’s M=735.32; F(63, 338345) =11.34; p<0.001), therefore the assumption that there were equal population covariance matrices, which did not depend on the population from which the data were obtained was valid and thus justifying the use of linear discriminant function (Gail et al., 2007; Quinn and Keough 2002). All the 384 observations were used in the analysis, with equal prior probability of 25%, thus 96 cases per primate species. The first three canonical discriminant functions were used. The eigenvalues for the first two discriminant functions were 0.5 and 0.07, their canonical correlations were 0.58 and 0.25 respectively and their percentage of variance reached 85.5 and 11.2, respectively. This means that after a cumulative 96.7% between group variance explained by the first two functions, not much of the between group variance remained, to inspect higher dimensions.
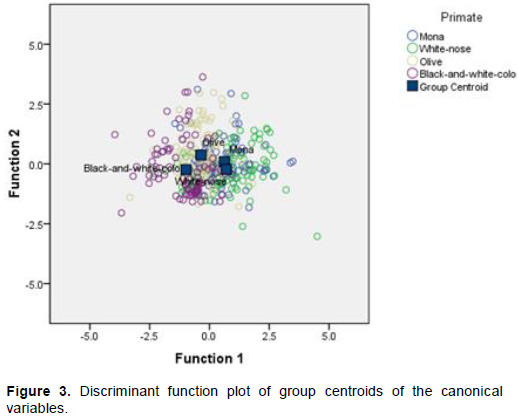
Dimensionality test for group discrimination showed χ2=183.979 with 18 degrees of freedom and the probability that a χ2 with larger value was found was p<<0.001 for the first discriminant function. For the second discriminant function, χ2=31.07 with 10 degrees of freedom and the probability that a χ2 with larger value was found was p = 0.001; and for the third discriminant function, χ2=7.08 with 4 degrees of freedom and the probability that a χ2 with larger value was found was p = 0.132. Therefore, on the account of the first two discriminant functions, the null hypothesis that there was no discrimination on any dimension was rejected. At least two discriminant functions were needed to describe group differences. Thus, the discriminant function is a very useful tool for the discrimination of the primate groups and therefore it was used for classifying observations. Significant Wilks’ Lambda for the first two discriminant functions also implied that the null hypothesis that there was no overall group effect was rejected. There was a significant group effect. The discriminant function scores obtained from standardized canonical coefficients and a discriminant function plot using group centroids of the canonical variables (Figure 3) confirmed that feeding sites and food types selected by the four primate species were separated by two discriminant functions. In general, the two discriminant functions significantly accounted for the between group variability; the first discriminant function separated the four primate species by 85.5% (Wilks’ Lambda = 0.615, p << 0.001) and the second, 11.2% (Wilk’s Lambda = 0.921, p = 0.001). The discriminant functions were:


(Where, LC = Lower canopy, FL = Flower, MC = Middle canopy, S = Seed, UC = Upper canopy and FR = Fruit).
The first discriminant function discriminated feeding sites and food types selected by the primates (Figure 3). C. polykomos and P. verus selected similar sites and food types, on the upper canopy. C. polykomos selected more seeds and P. verus selected more fruits. C. mona and C. petaurista selected similar sites and food types, mostly on the lower canopy and selected flowers mostly. The first function did not separate the C. mona and C. petaurista very clearly (Figure 3). However, along the second discriminant function, C. mona and C. petaurista were separated narrowly, with the C. petaurista selecting more flowers than C. mona, and C. mona selecting the lower canopy more than C. petaurista. C. polykomos and P. verus colobus were further separated along the second function. P. verus preferred the middle canopy to the upper canopy (Figure 3).
Seasonal changes in feeding site selection
Primates’ selection of the upper and middle canopies, flower, fruit and seed was significantly different between rainy and dry seasons (F(1, 382) = 13.9, p << 0.001; F(1, 382) = 27.29, p <<0.001; F(1, 382) = 8.002, p = 0.005; F(1, 382) = 33.74, p<< 0.001; and F(1, 382) = 64.06, p << 0.001, respectively). Seasonal difference in primates’ selection of the lower canopy was not significant (F(1, 382) = 1.097, p = 0.296). Primates used upper canopy more in the rainy season (mean ± se = 14.45±1.2) than in the dry season (mean ± se = 9.32±0.67); middle canopy more in the dry season (mean ± se = 13.17±0.12) than the rainy season (mean ± se = 8.07±0.53); flowers more in the rainy season (mean ± se = 10.21±0.53) than in the dry season (mean ± se = 8.11±0.52); fruit more in the rainy season (mean ± se = 17.69±0.7) than in the dry season (mean ± se = 12.58±0.54); and seed more in the dry season (mean ± se = 16.7±0.84) than in the rainy season (8.43±0.6).
Out of the 192 observations for the rainy season, 64.6% were correctly classified. Feeding site and food type selected by the four primates during the rainy season were separated by the first two discriminant functions significantly (Wilk’s Lambda = 0.231, p = <<0.001 and Wilk’s Lambda = 0.838, p << 0.001 respectively) (Table 3) and a discriminant function plot (Figure 4) confirmed that feeding sites and food types selected by the four primate species were separated by two discriminant functions. Tukey’s post hoc analysis revealed that along the first discriminant function, the differences in feeding site and food type selection by the primates were significant in all cases of comparisons except between the C. mona and C. petaurista (Table 4). Evaluation of discriminant scores (Table 5) and a one-way ANOVA conducted with discriminant scores and primates (Table 4), showed that along the first discriminant function, C. mona and C. petaurista significantly selected similar feeding sites, mainly the lower canopy while upper canopy significantly discriminated C. polykomos and middle canopy discriminated P. verus. Along the second discriminant function, C. petaurista was discriminated by fruit on the middle canopy to separate it from the C. mona while C. polykomos was further discriminated by seed.



There were 192 observations for the dry season and 56.3% of the original grouped cases were correctly classified. During the dry season, feeding site and food type selected by the four primates were separated by the first two discriminant functions significantly (Wilk’s Lambda = 0.49, p << 0.001 and Wilk’s Lambda = 0.767, p<< 0.001, respectively (Table 3) and a discriminant function plot (Figure 4) confirmed that feeding sites and food types selected by the four primate species were separated by two discriminant functions. Tukey’s post hoc analysis revealed that along the first discriminant function, the differences in feeding site and food type selection by the primates were significant only between C. petaurista and P. verus, and between P. verus and C. polykomos (Table 5). Evaluation of discriminant scores (Table 5) and a one-way ANOVA conducted with discriminant scores and primates (Table 4) showed that the most important feeding site in the first function was seed, which discriminated C. polykomos, while in the second function, fruit discriminated the P. verus.
Spatial variation in feeding site and food type selection
The differences in primates’ selection of feeding sites and food types at the three different zones of KCA were significant along the first function (F(2, 383) = 23.315, p << 0.001) and second function (F(2, 383) = 4.574, p = 0.01). The upper canopy, middle canopy and seed were significantly more selected by the primates at the centre of the reserve than other zones, reaching the highest means of abundance (± se) of 14.3±1.57, 12.8±1.06 and 16.83±1.16, respectively (Table 6). The lower canopy was significantly more selected at the periphery than other zones, reaching the highest mean abundance (± se) of 16.98±1.42 (Table 6). Flower and fruit reached the highest means of abundance (± se) of 10.39±0.81 and 15.67±0.65, respectively at the periphery and centre; but these were not significant (Table 6). C. polykomos selected the centre more than the other monkeys, reaching the highest mean abundance (± se) of 16.6 ± 2.4 (Table 7); C. petaurista selected the inner part more than the other monkeys (mean abundance ± se = 16.9 ± 1.2); and C. mona selected the periphery more than the other monkeys (mean abundance ± se = 14.7 ± 1.3) (Table 7). The selection of the different parts of KCA followed a pattern in which the selection by C. polykomos reduced from the centre (mean abundance ± se = 16.6 ± 2.4) through the inner part (mean abundance ± se = 10.5 ± 0.2) to the periphery (mean abundance ± se = 6.6 ± 1.6); and the selection by C. mona reduced from the periphery (mean abundance ± se = 14.7 ± 1.3) through the inner part (mean abundance ± se = 12.2 ± 1.3) to the centre (mean abundance ± se = 11.2 ± 1.6) (Table 7). Selection by C. petaurista and P. verus followed no pattern (Table 7).

Out of the 128 observations made at the centre of the reserve, 53.9% were correctly classified. Feeding sites and food types selected by the four primates at the centre of the reserve were separated by the first two discriminant functions, but only the first function discriminated significantly (Wilk’s Lambda = 0.597, p << 0.001, and Wilk’s Lambda = 0.88, p = 0.113, respectively) (Table 3) and a discriminant function plot (Figure 4) confirmed that feeding sites and food types selected by the four primate species were separated by two discriminant functions. Tukey’s post hoc analysis revealed that along the first discriminant function, the differences in feeding site selection by the primates were significant only between the C. polykomos and the other monkeys (Table 4). The most important feeding site or food type in the first function was seed, which discriminated the C. polykomos (Table 5).
Out of the 128 observations made at the inner part of the reserve, 56.3% were correctly classified. In the inner part of the reserve, between the periphery and the centre, feeding sites and food types were separated by the first two discriminant functions significantly (Wilk’s Lambda = 0.447, p << 0.001 and Wilk’s Lambda = 0.735, p << 0.001, respectively) (Table 3) and a discriminant function plot (Figure 4) confirmed that feeding sites and food types selected by the four primate species were separated by two discriminant functions. Tukey’s post hoc analysis revealed that along the first function, the differences in feeding site and food type selection by the primates were significant between primate species studied, except between C. mona and C. petaurista, and along the second function the differences were significant between C. mona and C. petaurista and between C. petaurista and P. verus (Table 4). Along the first function, seed discriminated C. polykomos, fruit discriminated P. verus, lower canopy discriminated C. mona and middle canopy discriminated C. petaurista (Table 5). Along the second function, fruit and seed discriminated P. verus, flower discriminated C. mona, upper canopy discriminated C. polykomos and lower canopy, C. petaurista (Table 5).
In the peripheral parts of the reserve, 65.6% of the 128 observations were correctly classified. Feeding sites and food types were discriminated by the first two discriminant functions significantly (Wilk’s Lambda = 0.182, p<< 0.001 and Wilk’s Lambda = 0.654, p <<0.001, respectively (Table 3) and a discriminant function plot (Figure 4) confirmed that feeding sites and food types selected by the four primate species were separated by two discriminant functions.. Tukey’s post hoc analysis revealed that along the first function, the differences in feeding sites and food types selection by the primates were significant between primate species studied, except between C. mona and C. petaurista, and P. verus and C. polykomos (Table 4; and along the second function the differences between primates were significant except between C. mona and C. petaurista, and C. petaurista and C. polykomos (Table 4). The first function separated the four primates. Flower and lower canopy discriminated C. petaurista, fruit discriminated C. mona, upper canopy discriminated C. polykomos and middle canopy discriminated P. verus (Table 5). Along the second function, middle canopy discriminated P. verus, fruits discriminated C. mona, upper canopy discriminated C. polykomos and lower canopy discriminated C. petaurista (Table 5).
Feeding sites and food types selected by primates
Research investigations indicate that co-existence within a guild is made possible by resource partitioning, which is the use of the same resource in different ways by different species of animals (Jarman and Sinclair, 1979). Plant forage has been widely documented as an important factor that allows co-existence within guilds (Ben-Shahar and Skinner, 1988; Mysterud, 2000). Indeed, the results revealed that flower, fruit and seed were all abundant in all three canopies and that the primates were ecologically separated. For example, primates which predominantly selected the upper canopy rarely selected the lower canopy and those that predominantly selected the lower canopy rarely selected the upper canopy. Furthermore, where the selection of flowers as food was common, the selection of fruit was common too, but selection of seed was rare; and where selection of seed was common selection of flower was rare. All these were necessary to allow the primates to discriminate among feeding resources. It is clear from the results that C. polykomos and C. petaurista did not overlap, because predominantly, C. polykomos selected upper canopy while C. petaurista selected lower canopy. Again, C. polykomos selected seed but C. petaurista selected flowers. C. polykomos and C. mona did not overlap for the same reasons. Though C. polykomos and P. verus shared the same feeding sites and food types on the upper canopy in many observations, they were separated by their different choices of food, because C. polykomos selected seed and P. verus selected fruit. The two colobus monkeys were separated also, because P. verus selected middle canopy. C. mona and C. petaurista shared the same feeding sites and food types for similar reasons. Thus, this study confirmed the occurrence of resource partitioning among co-existing primate species in tree canopies at KCA, as observed by earlier researchers elsewhere (Jarman and Sinclair, 1979; Schoener, 1986; Owen-Smith, 1989; Bailey et al., 1996; Wallis de Vries et al., 1999; Ritchie, 2009).
At KCA, resource partitioning among the primates occurred at the level of two factors, namely canopy type and food type. As the primates were able to perceive differences presented by the many combinations of the levels of these factors they were discriminated by the different choices they made, which allowed them to co-exist, while reducing competition. The phenology of tropical plants is a little difficult to explain completely (Ewusie, 1992), but this makes flowers, fruits and/or seeds available at any time of the year for evaluation by foraging primates to avoid competition in order to co-exist as suggested by Ritchie (2009).
Seasonal changes in feeding site and food type selection
The study supported the hypothesis that seasonal changes influenced resource partitioning at KCA. For example, in general, flower and fruit were selected by the primates more in the rainy season than in the dry season; while seed was predominantly selected in the dry season. Also, while there was no seasonal variation in the selection of lower canopy, the upper canopy was selected by primates more predominantly in the rainy season; while the middle canopy was selected more predominantly in the dry season. This may explain why C. petaurista shifted between lower canopy and middle canopy and P. verus, between upper canopy and middle canopy. These shifts in feeding site and food type selection were necessary to allow co-existence.
Spatial variation in feeding site selection
Spatial variation in canopy type availability in the various zones was to be expected for a reserve undergoing regeneration (Dakwa, 2016) after heavy logging in the past (WD, 1996). Though each zone was an admixture of all the three canopy types, lower canopy was more abundant and upper canopy uncommon at the periphery of the reserve, where logging was heaviest. At the centre, in which there were only minimal disturbances, upper canopy was more abundant and lower canopy uncommon. This was to be expected since logging removed upper canopy from the periphery mostly and regeneration added more of lower canopy to the periphery. Since the centre depicted a climax community, lower canopy was naturally rare. The study also supported the hypothesis that resource partitioning was influenced by spatial variation of feeding sites. The upper canopy, middle canopy and seed were more predominantly selected by primates at the central parts of KCA while lower canopy was selected more predominantly at the peripheral parts of KCA but there was no clear pattern in the case of primates’ selection of feeding sites and food types at the inner parts of the reserve, between the centre and the periphery. It is also clear from the results that C. polykomos had low tolerance to the disturbances at KCA and therefore were withdrawn to the most interior parts of the reserve where disturbances such as logging were minimal; and this is consistent with previous observations at KCA (Dakwa, 2016), and elsewhere (Fetene et al., 2011). On the other hand, C. mona was more associated with more disturbed parts of the reserve, at the periphery while C. petaurista and P. verus tended to be forest generalists, flexible in relation to both feeding and spatial options and were likely to be ubiquitous. Therefore, the main influential factor behind their resource selection and distribution seemed to be avoiding competition with other primates to allow co-existence. The logging event that hit KCA likely affected the spatial options for feeding site selection by C. polykomos most but appeared to have favoured C. mona. It is therefore more likely that C. polykomos will occur at the lowest density among the primates studied. The study missed the chance of evaluating the feeding site and food type selection options by the Cercopithecus diana (Diana monkey) compared to the other monkeys. However, it is possible that C. diana, which now lives at a very low density was affected, drastically, by competition with other monkeys, resulting from shrunken feeding resources during the logging regime. Considering that the C. diana is listed in IUCN category of threats as vulnerable (Oates et al, 2016), and also the C. polykomos and P. verus as vulnerable and near threatened, respectively (Oates et al., 2008), there is need to flag KCA for conservation priorities to sustain the populations of the monkey species.
In conclusion, the use of linear discriminant function analysis was successful in giving adequate insight to how various feeding sites and food types have contributed to the ecological separation and hence co-existence of the four primate species studied at KCA. The study was consistent with the hypothesis that variability in feeding resources facilitated differential use of the resources, which allowed the primate species to co-exist. Feeding resources partitioned among primates could be influenced not only by seasonal and spatial variations of feeding sites but also the prevailing forest conditions in different zones of the KCA landscape. Therefore, management of KCA should consider strict measures to facilitate forest regeneration, especially at the peripheral parts of the reserve as this is important for the conservation of the primates at KCA.
The authors have not declared any conflict of interests.
REFERENCES
|
Barnes RFW (1999). Is there a future for elephants in West Africa? Mammal Review 29:175-200.
Crossref
|
|
|
|
Bailey DW, Gross JE, Laca EA, Rittenhouse LR, Coughenour MB, Swift DM, Sims PL (1996). Mechanisms that result in large herbivore grazing distribution patterns.Journal of Range Management 49:386–400.
Crossref
|
|
|
|
|
Ben-Shahar R, Skinner JD (1988). Habitat preferences of African ungulates derived by uni- and multivariate analyses. Ecology 53:1479-1485.
Crossref
|
|
|
|
|
Chapman CA (1989). Primate seed dispersal: The fate of dispersed seeds. Biotropica 21:148-154.
Crossref
|
|
|
|
|
Cincotta RP, Wisnewski J, Engelman R (2000). Human population in the biodiversity hotspots. Nature 404(6781):990-992.
Crossref
|
|
|
|
|
Cowlishaw G, Dunbar R (2000). Primate Conservation Biology. London: The University of Chicago Press.
|
|
|
|
|
Dakwa KB (2016). Monitoring and evaluation of vertebrate fauna composition and structure of a high forest zone in Ghana, 20 years after heavy logging. Journal of Biology and Nature 5(4):196-210.
|
|
|
|
|
Dakwa KB, Monney KA, Attuquayefio D (2014). Density and distribution of bongos Tragelaphus eurycerus) in a high forest zone in Ghana. Journal of Ecology and the Natural Environment 6(9):331-341.
Crossref
|
|
|
|
|
Dakwa KB, Monney KA, Attuquayefio D (2016). Raid range selection by elephants around Kakum Conservation Area: Implications for the identification of suitable mitigating measures. International Journal of Biodiversity and Conservation 8(2):21-31.
Crossref
|
|
|
|
|
Duc HM, Baxter GS, Page MJ (2009). Diet of Pygathrix nigripes in Southern Veitnam. International Journal of Primatology 30: 15-28.
Crossref
|
|
|
|
|
Ewusie JY (1992). Phenology in Tropical Ecology. Accra: Ghana Universities Press.
|
|
|
|
|
Fa JE, Juste J, del Val Perez J, Castroviejo J (1995). Impact of market hunting on mammal species in Equatorial Guinea. Conservation Biology 9:1107-1115.
Crossref
|
|
|
|
|
FC-Forestry Commission (2007). Monitoring staff deployment, patrol effort, illegal activity and wildlife trends to facilitate adaptive and performance management in nine protected areas in Ghana. Second Evaluation, Accra.
|
|
|
|
|
Fetene A, Mengesha G, Bekele T (2011). Spatial distribution and habitat references of selected large mammalian species in the Nech SAR National Park(NSNP), Ethiopia. Natural Science 9:80-90.
|
|
|
|
|
Gail M, Krickeberg K, Samet J, Tsiatis A, Wong W (2007). Statistics for biology and health. New York: Springer.
|
|
|
|
|
Gautier-Hion A (1970). L'ecologie du talapion du Gabon. Terreet Vie 25:427-490.
|
|
|
|
|
Jarman PJ, Sinclair ARE (1979). Feeding strategy and the pattern of resource partitioning in ungulates. Serengeti: dynamics of an ecosystem. Chicago:University of Chicago Press.
|
|
|
|
|
Johns AD, Skorupa JP (1987). Responses of rain-forest primates to habitat disturbance: A review. International Journal of Primatology 8:157-191.
Crossref
|
|
|
|
|
Martin C, Asibey EOA (1979). Effects of timber exploitation on primate population distribution in the Bia rain forest area of Ghana. Paper presented at the 7th Congress of the International Primatological Society, Bangalore, India.
|
|
|
|
|
Monney KA, Dakwa KB, Wiafe ED (2010). Assessment of crop raiding situation by elephants (Loxodonta africana cyclotis) in farms around Kakum Conservation Area, Ghana. International Journal of Biodiversity and Conservation 2(9):243-249.
|
|
|
|
|
Mysterud A (2000). Diet overlap among ruminants in Fennoscandia. Oecologia 124(1):130-137.
Crossref
|
|
|
|
|
Newmark WD (1996). Insularization of Tanzanian parks and local extinction of large mammals. Conservation Biology 10:1549-1556
Crossref
|
|
|
|
|
Oates JF (1996). African Primates: Status Survey and Conservation Action Plans. Revised Edition. Gland, Switzerland: International Union of Conservation of Nature and Natural Resources.
Crossref
|
|
|
|
|
Oates JF, Gippoliti S, Groves CP (2008). Procolobus verus and Colobus polykomos. The IUCN Red List of Threatened Species.
|
|
|
|
|
Oates JF, Gippoliti S, Groves CP (2016). Cercopithecus diana. The IUCN Red List of Threatened Species.
|
|
|
|
|
Owen-Smith N (1989). Megafaunal Extinctions: The Conservation Message from 11,000 Years Ago. Conservation Biology 3(4):405-412.
Crossref
|
|
|
|
|
Peres CA (1994). Primate responses to phenological change in an Amazonian terrafirme forest. Biotropica 26:98-112.
Crossref
|
|
|
|
|
Quinn GP, Keough MJ (2002). Experimental design and data analysis for biologists. New York: Cambridge University Press.
Crossref
|
|
|
|
|
Ritchie ME (2009). Scale, heterogeneity, and the structure and diversity of ecological communities. New Jersey: Princeton University Press.
Crossref
|
|
|
|
|
Schoener TW (1986). Overview: kinds of ecological communities-ecology becomes pluralistic. Community ecology. New York: Harper and Row.
|
|
|
|
|
Turchin P (1991). Translating foraging movements in heterogeneous environments into the spatial distribution of foragers. Ecology 72(4):1253-1266.
Crossref
|
|
|
|
|
Wallis de Vries MF, Laca EA, Demment MW (1999). The importance of scale of patchiness for selectivity in grazing herbivores. Oecologia 121:355-363.
Crossref
|
|
|
|
|
Wildlife Department (WD) (1996). Kakum National Park and Assin AttandansoResource Reserve. The Management Plan. Accra: Ghana Wildlife Department.
|
|
|
|
|
Wiafe ED, Dakwa KB, Yeboah S (2010). Assemblages of avian communities in forest elephant (Loxodonta cyclotis) range in Ghana. Pachyderm 48:41-47.
|
|
|
|
|
Yeboah S (1996). Small mammals and the herpetofauna of the Kakum National Park. Facing the Storm. Proceedings of the Kakum Conservation Area Research Colloquium. Washington, DC: Conservation International.
|
|