ABSTRACT
The identification of factors that influence the timing of seedling emergence in topsoil is important to improve restoration strategies in former mining areas. The aim of this study was to determine the time of emergence of seedlings present in the topsoil of high fields in the Atlantic Forest on bauxite mines in the plateau region of Poços de Caldas, Minas Gerais, in both the dry and rainy seasons. The surface layer soil (top 5 cm) and litter were collected in August 2011 and March 2012. We collected 12 samples, consisting of 6 subsamples distributed in 3 sites (4 samples/site), from both the dry and wet periods (n = 24). The collected contents were placed in plastic trays and germination was evaluated in the greenhouse. The seedling emergence curve was adjusted. The values were submitted to analysis of variance associated with the Scott-Knott test (p < 0.05). The number of emerged seedlings and the time required for emergence to stop were lower in the rainy season than the dry season. The results show that the best time for the storage of the topsoil layer is the dry season, since it prolongs the viability of propagules and improves the success of recovery activities on mined areas.
Key words: Atlantic forest, environmental restoration, emergence time, degraded areas, bauxite mining.
Fields Plateaus in southeast Brazil contain rare vegetation and are considered areas of great environmental importance due to their high level of endemism (Caiafa and Silva, 2005). This plant communities occurs in open areas (Vasconcelos, 2011) covered by bunchgrass matrixes and some pteridophytes (Safford, 1999), and is usually found at an altitude of around 1,500 m (Vasconcelos, 2011). The plateaus deserve attention for their scenic beauty, biological importance, and geological particularities, standing out as sources of primary water capture, supplying water to approximately 25% of the Brazilian population (Safford, 1999).
The Poços de Caldas Plateaus are located within the Atlantic Forest ecosystem (Veloso et al., 1991) with a rich biodiversity and high level of endemism, where large deposits of bauxite are being mined in areas of typical regional vegetation. The knowledge and awareness of the impacts generated by mining in the region, as well as its importance for the national and international economies, motivate practices related to the conservation and recovery of these environments (Barros et al., 2012). The topsoil seed bank may be an indicator of the behavior of the species in areas subjected to mining (Martins et al., 2008; Machado et al., 2013), thus studying it provides important information for the development of conservation plans (Adams et al., 2005). Therefore, the evaluation of the emergence time of propagules in certain environments, taking into account spatial and temporal patterns (Ceccon and Hernández, 2009), allows for the identification of periods of higher propagule emergence, as well as periods with greater plant species diversity. This information can improve the expected results of environmental restoration plans that, among other factors, also include indicators of plant performance, physical and chemical aspects of the soil, and the impact on the fauna (Almeida and Sánchez, 2005; Mensah, 2015). Studies making use of this approach are important to gather cumulative data about the timing and plant diversity of propagule emergence, allowing inferences about the optimal periods to implement restoration practices in altered environments. Analysis of the propagule emergence time under controlled conditions is widely used for species of commercial interest to address the needs of seed and grain producers (Caldeira et al., 2015). However, few studies have been developed using this analysis for plant conservation in natural environments (Machado et al., 2013).
The removal, stocking, and returning of topsoil are used to promote environmental recovery after mining activities (Hall et al., 2010; Barros et al., 2012; Jaunatre et al., 2014). The topsoil, the layer that includes the surface horizons of the soil, is rich in organic matter, autochthonous seeds, and microorganisms, and is considered to be of the utmost importance in the restoration of altered environments (Koch, 2007). The knowledge of the wealth of plant species that make up the soil seed bank provides information on the conservation status of the area and will certainly contribute to the development of appropriate management protocols for vegetation condition (Fisher et al., 2009).
The returning of this surface layer is also recommended to maintain the physical characteristics of soil as close as possible to the existing prior to mining (Barros et al., 2013), since it must be considered the influence of storage time on the longevity of the existing propagules and their influence on the quality of soil and seeds bank. Therefore, this study aimed to determine the duration and timing of emergence of propagules collected in two periods (dry and rainy seasons) to optimize techniques for restoring bauxite mining areas in the Poços de Caldas Plateaus region of Minas Gerais, Brazil.
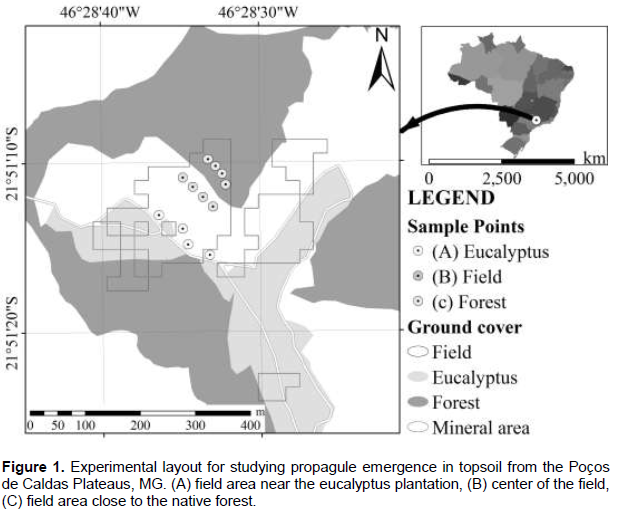
The study area is located in the Poços de Caldas Plateaus, at an altitude of around 1,300 m, where the forest formations are classified as High-mountain Semi-deciduous Seasonal Forest (Oliveira Filho and Fontes, 2000), surrounded by native fields. The climate in this region is mesothermic, of the Cwb type (Alvares et al., 2013) with an annual average precipitation of 1,300 to 1,600 mm for the rainy season (October to March) and 300 mm for the dry season (April to September) (Guimarães et al., 2013). The collection of potential propagative material for restoration activities was carried out in August 2011 (winter dry season) and March 2012 (summer rainy season). The upper 5 cm of the topsoil was collected after removing the vegetation (Araújo et al., 2004). Soils classified as dystrophic Red Nitosol predominate in the study area (Embrapa, 2006), it is characterized as typically porous, high acidity, and in the case of the study area of large amounts of clay, gravel and strongly undulating terrain. The study area was divided into 3 sites: (A) close to a eucalyptus stand, (B) in the center of the field, and (C) in the field close to the native forest (Figure 1). The sampling was comprised of a total of 12 samples that included 6 subsamples (Souza et al., 2006), with 4 samples being collected per site for each period (N = 24). Immediately after collection, the samples were sent to a greenhouse at the forestry nursery of the Department of Forest Sciences (DCF) of the Federal University of Lavras (UFLA) and placed under a shade cloth at 50% luminosity. Each soil sample was spread over a 1.5 cm sterilized sand bed, autoclaved (120°C for 1 h), and placed in 33.00 × 44.00 × 8.00 cm plastic trays according to the methodologies proposed by Araújo et al. (2004) and Zhang et al. (2001). The soil samples collected were approximately 0.012 m³.
The method used to quantify the individuals present in the topsoil was counting the daily emergence of propagules (Gross, 1990). The plants were identified and removed from the trays after reaching the adult stage (flowering, when possible), with proper care to avoid contamination of the propagules. The evaluations were performed for 11 weeks, until the emergence of new individuals ended. The maximum average temperature during the rainy season was 27.1°C and the minimum was 16.66°C, with an average relative humidity of around 75%. On the other hand, in the dry season, the maximum average temperature was 28.3°C and the minimum was 14°C, with an average relative humidity of around 58.7%. The propagules presents in the substrate were Irrigated every 24 hours using 500 mL of water in each tray.Data on the number of propagules that emerged in the samples were used to adjust the emergence curve of plateau plants using the software Germinator (Joosen et al., 2010), as described by El-Kassaby et al. (2008), using the parameters of the Hill function (4PHF):
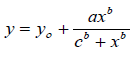
Where: y = percentage of cumulative emergence in x time; y0 = y-axis intercept; b = exponent that controls the form and degree of inclination in the curve; c = time necessary for 50% of viable propagules to emerge (t50); x = assessment time; and a = maximum cumulative emergence percentage.
The adjustment of the cumulative emergence curves indicates the biological behavior before specific environmental conditions (Joosen et al., 2010). In this study, the values found for the adjustment of the emergence time were subjected to variance analysis, having as variables: t50 (necessary time for 50% of the viable propagules to emerge) and u7525 (time interval between the emergence of 25 and 75% of propagules). The design used was completely randomized, in a 3 × 2 factorial scheme (3 areas and 2 periods) with 4 replicates (n = 24). To compare averages, an analysis of variance was performed in association with a Scott-Knott test with p < 0.05 using the programming language medium R (R, 2009). After Shapiro Wilk’s and Levene’s tests, it was found that the data were normally distributed (p > 0.05) and with homogeneous variances (p > 0.05), respectively, thus enabling us to submit them to variance analysis with p < 0.05.
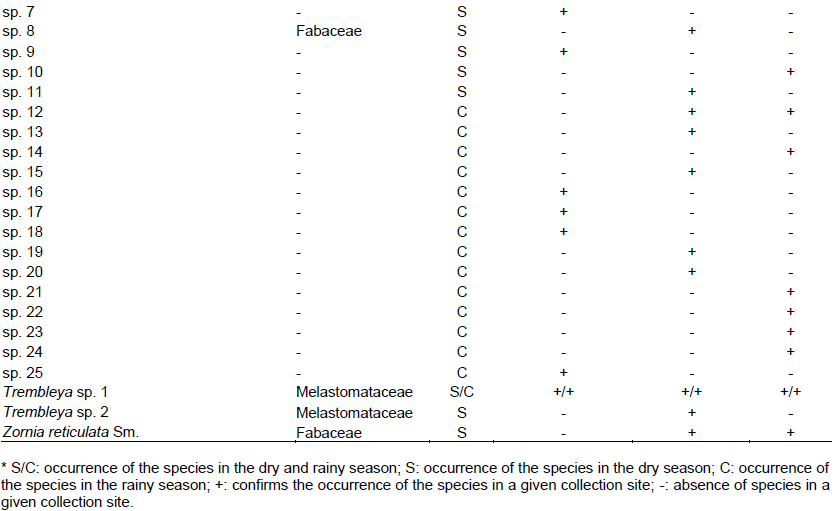
The first propagules emerged in the first week during the rainy season and ended during the fifth week, whereas in the dry season, seedling emergence began in the third week and ended during the tenth week. In total, 585 individuals emerged in the rainy period, and 777 in the dry period. Seven botanical families were identified during the rainy period, comprising 34 species (Table 1). Among these, 5 were identified only at the genus level, and 14 were classified as morphospecies. Among the identified families, Asteraceae showed the greatest species richness in this period, (7 species, 20.59%), followed by Poaceae (6 species, 17.65%), and Rubiaceae and Melastomataceae, both with 2 species (5.88%). In the dry period, 50 species from 14 botanical families were identified. Among the species, 10 were identified only at the genus level and 11 as morphospecies. Among the families identified, Poaceae showed the highest richness (13 species, 26.00%),followed by Asteraceae (7 species, 14.00%) and Melastomataceae with 4 species (8.00%). These plant families predominate in altitude fields, suggesting great ecological importance in studies of restoration areas under the influence of bauxite mining. There was a predominance of plant species classified as herbaceous. The abundance of individuals in dry season, there was a predominance of the family Asteraceae, primarily related to the abundance of Ageratum fastigiatum (163 individuals), which stands out for its representation of the herbaceous habit in grassland vegetation types. Other species of large occurrence in this period were: Achyrocline satureioides (121 individuals), followed by Echinolaena inflexa (90 individuals) Achyrocline alata (87 individuals), Borreria latifolia (43 individuals) and Melinis minutiflora (22 individuals). In rainy season prevailed also Asteraceae family. Again the patterns abundance of A. fastigiatum (225 individuals) in rainy season were higher. Other species of large occurrence in rainy season were: Gamochaeta americana (71 individuals), followed by Borreria latifolia (75 individuals), Paspalum sp. (38 individuals), Phyllanthus niruri (37 individuals) and Echinolaena inflexa (31 individuals). We highlighted the occurrence of two endemic species from Brazil: Panicum campestre and Miconia cinnamomifolia, suggesting greater concern for conservation of open fields. The total species richness was 77 species. Regarding species richness that occurred in each area investigated there was a higher occurrence of species in propagules bank of samples collected in the field strictu sensu and forest environment (40 species), followed by eucalyptus plantation (34 species) and 17 species shared in the three areas covered in this study, equivalent to 23.37% of total species richness (Table 1). Some species occurred mainly in a period or specific collection site. Among these species there are A. fastigiatum, A. satureoides, B. latifolia, E. inflexa and Melinis minutiflora as the most frequent occurrence, the latter being an exotic species of high fields from the Atlantic forest.

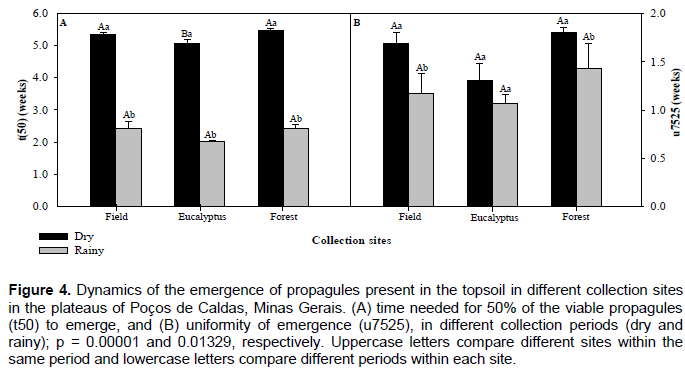
We found a significant difference only for the period (dry and wet) (p < 0.01), which suggests a low contribution of the surrounding areas (field area near the eucalyptus plantation and field area close to the native forest) (p > 0.05) for the restoration of the studied field area. Considering the periods, it was found that dry period showed higher plant species richness when compared to the rainy season (Figure 2). There were distinct differences in the time of emergence between the dry and rainy seasons; propagule emergence ended sooner in the rainy season (Figure 3). In the rainy season, the propagules germinated more quickly, which would cause significant losses if topsoil stocking was carried out during this period. On the other hand, a longer period of propagule emergence was observed in the dry period, in addition to a higher number and richness of species, suggesting a greater viability of propagules, and, possibly, better environmental recovery after mining activities. As expected, the sooner the initial emergence, the shorter the time to reach 50% of the total emergence (t50) was. In this work, the greater propagule emergence speed was during the rainy period. Both t50 and u7525 were lower for this period (Figure 4), indicating that less time was needed for the stabilization of the cumulative emergence curve (p < 0.05). The variable t(50), was not significantly different between the center of the field area and the area close to the native forest. However, both were significantly different from the area close to the eucalyptus stand (p < 0.01) (Figure 4). In addition, there was a shorter interval of time between the emergence of 25 and 75% of propagules for the samples collected in the area close to the eucalyptus stand than for the other areas; however, there was no statistical difference in the u7525 variable among the areas analyzed during the dry and rainy seasons with p > 0.05 (Figure 4).


The emergence performance of propagules in an environment can be characterized through 3 parameters: the time of first emergence and the speed and duration of emergence (El-Kassaby et al., 2008). The early emergence of propagules from the plateau topsoil during the rainy season is related to the higher likelihood of appropriate conditions for germination due to higher humidity. This reflects synchrony between the reproductive strategy, which is concomitant with the dispersion of diaspores at the end of the dry season and emergence right at the beginning of the rainy season (Munhoz and Felfili, 2006). The composition of plant species is also important and should be considered in restoration studies on degraded areas, since restoration success is directly related to the favorableness of a period for a higher diversity of species, along with information of the impact on the soil and fauna (Almeida and Sánchez, 2005). In the case of the field environment under study, we identified more plant families and species during the dry season. In addition, the use of topsoil stock in this season allowed for greater time for the beginning of emergence of propagules, avoiding important losses of plant species and total individuals after restoration of the area. Studies on the speed and uniformity of emergence are useful not only to estimate the conversion of propagules into seedlings (Joosen et al., 2010), but also to direct management practices for minimizing the impact of mining and conserving the diversity of plant species (Kolotelo et al., 2001; Sheoran et al., 2010). From this perspective, the stocking of topsoil for restoring mining areas should be performed during periods of lower physiological activity of the seeds and greater longevity of propagule emergence. The viability of propagules in the long-term has been related to environmental parameters, such as humidity and temperature (Fenner and Thompson, 2005). The early emergence of seedlings during the rainy season in this study indicated a shorter propagule viability time when stocked in the form of static piles, increasing the possibility of losing propagation material (Golos and Dixon, 2014), since, in tropical regions, where up to 90% of rain is concentrated in the 3-month rainy period, storing at the soil level becomes a problematic activity (Sheoran et al., 2010). Seedling emergence and establishment are regulated by diverse factors, including abiotic conditions like humidity and temperature that influence the propagation of species (Eliud et al., 2009; Lebrija-Trejos et al., 2011). The first manifestations of emergence are characterized by the swelling of the propagules and the emergence of the radicle and cotyledon (Rosa et al., 2005). Overall, more propagules from recalcitrant species, which do not show strategies for early emergence, occur in dispersed areas during the rainy period, since their seeds do not dry at the end of maturation but are dispersed during elevated humidity, remaining metabolically active and germinating immediately after dispersion (Hong and Ellis, 1996; Carvalho et al., 2006). Recalcitrant species are more sensitive to drying, and their propagules cannot be stored for long periods (Barbedo and Bilia, 1998; Carvalho et al., 2006), indicating that some of the propagules from the Poços de Caldas Plateaus that emerged during the rainy season share this reproductive strategy. Some species of botany family Asteraceae, Poaceae, Rubiaceae and Melastomataceae are classified as recalcitrant (Farnsworth, 2000), and scatter seeds with high humidity can begin the process of germination quickly (Nery et al., 2014).
During the dry period, more propagules of orthodox species occur (Chazdon et al., 2003), which indicates greater tolerance to dryness when stored (Hong and Ellis, 1996; Barbedo and Bilia, 1998) in the topsoil, which makes them more efficient to use for restoration. On the other hand, seeds from recalcitrant species easily lose viability when subjected to unfavorable circumstances (Nazário et al., 2008), thus storing the upper level of soil for long periods may reduce the number of species restored (Barbedo and Bilia, 1998; Sheoran et al., 2010). According to what was observed in the present work, the rainy season is not the best period for stocking topsoil, based on the speed (t50) and uniformity of emergence (u7525) of the propagules (p < 0.05) (Figure 4). On the other hand, propagules emerged more persistently in the dry period, that is, for a longer period. Seed longevity increases in colder and less rainy conditions, whereas the loss of viability is increased under hot and humid conditions (Pakeman et al., 2012). In addition, excessive humidity accelerates seed germination and induces overcoming dormancy (Golos and Dixon, 2014). When mining work is planned for the rainy season, it is recommended that material removed from the station be quickly used to restore another area, reducing the possibility of propagule loss. Propagule dispersion and germination are important for the establishment and maintenance of plant species, and are important in natural regeneration (Harper, 1977; Deminicis et al., 2009), with the success of the recomposition being related to the type and quantity of seeds produced, size of seeds, maturation period, and environmental conditions (Lebrija-Trejos et al., 2011; Marini et al., 2012). In this study, faster propagule emergence and fewer individuals were observed in the rainy season, which suggest that topsoil stocking should be performed in the dry season to extend the stocking time and, possibly, promote the efficiency of the restoration of mined areas.
The time of emergence of the samples collected in the site near the eucalyptus area was different from the others, reaching its maximum accumulated germination more quickly, possibly because these samples feature the best conditions of humidity for germination. This suggests that areas of the same region with a different land-use history have different propagule emergence dynamics, which should be considered when planning restoration activities in degraded areas. However, richness and dispersion strategies, relief conditions, distance area after forests fragments, surrounding cultures and content moisture of propagules should be considered for activities recovery of high field in the Atlantic forest. The simulation of the propagule emergence time curve under controlled conditions may contribute to understanding the biological responses in the process of plant recolonization (Ikeda et al., 2008), and thus improve the practicality of techniques used in the restoration of mining areas, taking advantage of the best time for the beginning of the restoration process in the local community after the stocking period and topsoil return. After bauxite mining activities in the plateaus in Poços de Caldas, we did not observe significant variations in the main physical characteristics of the soil after returning the upper level of the soil (Barros et al., 2013). This indicates that the stocking period and topsoil return are of great importance and should be taken into account to avoid irreversible modifications in the soil structure that could affect plant establishment. An efficient model of restoration for a degraded system should consider the space (limits of the system), the subsystems (its components), and the time interval to be considered (Aumond and Maçaneiro, 2014).Thus, the planning of restoration activities is fundamental and must include considerations of the favorable periods for the establishment of plant species, especially in regard to the stocking time and the quantity of topsoil to be returned to the upper level during the post-mining restoration process (Salomão et al., 2014). The restoration activities of degraded environments need a systemic, multidisciplinary approach, supported by an ecological model that involves more information on the abiotic and biotic components of the altered ecosystem (Aumond and Maçaneiro, 2014).
Based on the conditions used in this work, it is possible to state that the dry season is better for restoration activities in plateau areas affected by bauxite mining, since the speed and time of emergence, as well as the richness of plant species were greater in this period, indicating a longer propagule viability time. From this work, we suggest that special attention should be given to the areas surrounding the plateau fields, as well as the plant community history, as part of considering the local characteristics, and, consequently increasing the possibility of success during restoration on mining areas.
The authors have not declared any conflict of interests.
REFERENCES
|
Adams VM, Marsh DM, Knox JS (2005). Importance of the seed bank for population viability and population monitoring in a threatened wetland herb. Biol. Conserv. 124:425-436.
Crossref
|
|
|
|
Almeida ROPO, Sánchez LE (2005). Monitoring and evaluating performance of mining revegetation. Revista Árvore. 29(1):47-54.
Crossref
|
|
|
|
|
Alvares CA, Stape JL, Sentelhas PC, Gonçalves JL de M, Sparovek G (2013). Köppen's climate classification map for Brazil. Meteorologische Zeitschrift. 22:711-728.
Crossref
|
|
|
|
|
Araújo MM, Longhi SJ, Barros PLC de, Brena DA (2004). Characterization of the seed rain, soil seed bank and seedling bank in Seasonal Deciduous Riparian Forest, Cachoeira do Sul, RS, Brazil. Scientia Florestalis. 66:128-141.
|
|
|
|
|
Aumond JJ, Maçaneiro JP de (2014). Systemic approach and roughness application to cause emerging properties in the restoration of degraded soils. Ciência Florestal. 24(3):759-764.
|
|
|
|
|
Barbedo CJ, Bilia DAC (1998). Evolution of research on recalcitrant seeds. Scientia Agricola. 55:121-125.
Crossref
|
|
|
|
|
Barros DA de, Guimarães JCC, Pereira JAA, Borges LAC, Silva RA, Pereira AAS (2012). Characterization of the bauxite mining of the Poços de Caldas alkaline massif and its socio-environmental impacts. REM: Revista Escola de Minas. 65:127-133.
Crossref
|
|
|
|
|
Barros DA de, Pereira JAA, Ferreira MM, Silva BM, Ferreira Filho D, Nascimento G de O (2013). Soil physical properties of high mountain fields under bauxite mining. Ciênc. Agrotecnol. 37:419-426.
Crossref
|
|
|
|
|
Brazilian Agricultural Research Corporation – EMBRAPA (2006). National Center for Soil Research. Brazilian system of soil classification. 2nd ed. Rio de Janeiro: Embrapa Soils. 306 pp.
|
|
|
|
|
Caiafa AN, Silva AF da (2005). Floristic composition and biological spectrum of a field altitude in State Park Serra do Brigadeiro, Minas Gerais – Brazil. Rodriguésia. 56:163-173.
|
|
|
|
|
Caldeira, CM, Carvalho MLM de, Oliveira JA, Kataoka VY, Freire AI (2015). Reduced time for evaluation of the germination test for sunflower seeds. J. Seed Sci. 37:70-75.
Crossref
|
|
|
|
|
Carvalho LR de, Silva EAA da, Davide AC (2006). Storage behaviour of forest seeds. Revista Brasileira de Sementes. 28:15-25.
|
|
|
|
|
Ceccon E, Hernandez P (2009). Seed rain dynamics following disturbance exclusion in a secondary tropical dry forest in Morelos, Mexico. Rev. Biol. Trop. 57:257-269.
|
|
|
|
|
Chazdon RL, Careaga S, Webb C, Vargas O (2003). Community and phylogenetic structure of reproductive traits of woody species in wet tropical forests. Ecol. Monogr. 73(3):331-348.
Crossref
|
|
|
|
|
Deminicis BB, Vieira HD, Araújo SAC, Jardim JG, Pádua FT, Chambela Neto A (2009). Natural dispersion of seeds: Importance, classification and dynamics in tropical pastures. Archivos de Zootecnia. 58:35-58.
|
|
|
|
|
Eliud R, Reuben M, Linnet G (2009). Longevity of bean (Phaseolus vulgaris) seeds stored at locations varying in temperature and relative
|
|
|
|
|
El-Kassaby YA, Moss I, Kolotelo D, Stoehr M (2008). Seed germination: mathematical representation and parameters extraction. For. Sci. 54:220-227.
|
|
|
|
|
Farnsworth E (2000). The ecology and physiology of viviparous and recalcitrant seeds. Ann. Rev. Ecol. Syst. 31:107-138.
Crossref
|
|
|
|
|
Fenner M, Thompson K (2005). The ecology of seeds. New York:Cambridge University Press. pp.250.
Crossref
|
|
|
|
|
Fisher JL, Loneragan WA, Dixon K, Veneklaas EJ. (2009). Soil seedbank compositional change constrains biodiversity in an invaded species-rich woodland. Biol. Conserv. 142(2):256-269.
Crossref
|
|
|
|
|
Golos PJ, Dixon KW (2014). Waterproofing topsoil stockpiles minimizes viability decline in the soil seed bank in an arid environment. Restor. Ecol. 22:495-501.
Crossref
|
|
|
|
|
Gross KL (1990). Comparison of methods for estimating seed numbers in the soil. J. Ecol. 78:1079-1093.
Crossref
|
|
|
|
|
Guimarães JCC, Barros DA de, Pereira JAA, Silva RA, Oliveira AD de, Borges LAC (2013). Cost analysis and ecological benefits of environmental recovery methodologies in bauxite mining. Cerne. 19:9-17.
Crossref
|
|
|
|
|
Hall SL, Barton CD, Baskin CC (2010). Topsoil seed bank of an oak- hickory forest in eastern Kentucky as a restoration tool on surface mines. Restor. Ecol. 18:834-842.
Crossref
|
|
|
|
|
Harper JL (1977). Population biology of plans. London: Academic. pp.892.
|
|
|
|
|
Hong TD, Ellis RH (1996). A protocol to determine seed storage behaviour. Reading: The University of Reading. P 62.
|
|
|
|
|
Ikeda FS, Carmona R, Mitja D, Guimarães RM (2008).Light and KNO3 on Ageratum conyzoides L. seed germination at constant and alternating temperature. Rev. Bras. Sementes. 30:193-199.
Crossref
|
|
|
|
|
Jaunatre R, Buisson E, Dutoit T (2014). Topsoil removal improves various restoration treatments of a Mediterranean steppe (La Crau,southeast France). Appl. Veg. Sci. 17:236-245.
Crossref
|
|
|
|
|
Joosen RVL, Kodde J, Willems LAJ, Ligterink W, Van Derplas LHW, Hilhorst HWM (2010). Germinator: a software package for high-throughput scoring and curve fitting of Arabidopsis seed germination. Plant J. 62:148-159.
Crossref
|
|
|
|
|
Koch JM (2007). Alcoa's mining and restoration process in South Western Australia. Restor. Ecol. 15:S11-S16.
Crossref
|
|
|
|
|
Kolotelo D, Van Steenis E, Peterson M, Bennett R, Trotter D, Dennis J (2001). Seed handling guidebook: BC ministry of forests, tree improvement branch. Victoria: USDA. P 106.
|
|
|
|
|
Lebrija-Trejos E, Lobato Maria-Cristina C, Sternberg M (2011). Reproductive traits and seed dynamics at two environmentally contrasting annual plant communities: from fieldwork to theoretical expectations. Israel J. Ecol. Evol. 57:73-90.
Crossref
|
|
|
|
|
Machado VM, Santos JB, Pereira IM, Lara RO, Cabral CM, Amaral CS (2013).Evaluation of the seed bank in a campestre cerrado area under recovery . Planta Daninha. 31:303-312.
Crossref
|
|
|
|
|
Marini L, Bruun HH, Heikkinen RK, Helm A, Honnay O, Krauss J, Kühn I, Lindborg R, Pärtel M, Bommarco R (2012). Traits related to species persistence and dispersal explain changes in plant communities subjected to habitat loss. Divers. Distributions 18:898-908.
Crossref
|
|
|
|
|
Martins SV, Almeida DP de, Fernandes LV, Ribeiro TM (2008).Seed bank as indicator of restoration of a kaolin mining degraded area in Brás Pires, MG . Revista Árvore. 32:1081-1088.
|
|
|
|
|
Mensah AK (2015). Role of revegetation in restoring fertility of degraded mined solis in Ghana: A review. Int. J. Biodivers. Conserv. 7:57-80.
Crossref
|
|
|
|
|
Munhoz CBR, Felfili JM (2006). Phytosociology the herbaceous and subshrub layer of an area of dirty field in Distrito Federal, Brazil. ActaBotanica Brasílica. 20:671-685.
Crossref
|
|
|
|
|
Nazário P, Ferreira SA do N, Rebouças ER (2008). Germination of Cynometra bauhiniifolia Benthan (jutairana) seeds in terms of desiccation and maintenance under humid conditions. Acta Amazônica. 38:439-444.
Crossref
|
|
|
|
|
Nery MC, Davide AC, Silva EAA,Soares GCM, Nery FC (2014). Seed storage behavior of forest tree species seeds. Cerne. 20(3):477-483.
Crossref
|
|
|
|
|
Oliveira-Filho AT, Fontes MAL (2000). Patterns of floristic differentiation among Atlantic Forests in Southeastern Brazil and the Influence of Climate. Biotropica. 32:793-810.
Crossref
|
|
|
|
|
Pakeman RJ, Small JL, Torvell L (2012). Edaphic factors influence the longevity of seeds in the soil. Plant Ecol. 213:57-65.
Crossref
|
|
|
|
|
R Development Core Team (2009). R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing, 2009. Online: 2012.
View
|
|
|
|
|
Rosa LS da, Felippi M, Nogueira AC, Grossi F (2005). Germination assessment on different osmotic potentials and seed and seedling morphologic characterization of the Ateleia glazioviana Baill (Timbo). Cerne. 11:306-314.
|
|
|
|
|
Safford HD (1999). Brazilian Páramos I: an introduction to the physical environment and vegetation of the campos de altitude. J. Biogeogr. 26:693-712.
Crossref
|
|
|
|
|
Salomão RP, Brienza Júnior S, Rosa NA (2014). Reforestation dynamics in areas of restoration after mining in unit of forest conservation in the Amazon. Revista Árvore. 38:1-24.
Crossref
|
|
|
|
|
Sheoran V, Sheoran AS, Poonia P (2010). Soil reclamation of abandoned mine land by revegetation: a review. Int. J. Soil Sediment Water. 3(2). Online: View
|
|
|
|
|
Souza PA de, Venturin N, Griffith JJ, Martins SV (2006). Evaluation of a seed bank contained in the litter of a forest fragment envisaging the recovery of degraded areas. Cerne. 12:56-67.
|
|
|
|
|
Vasconcelos MF de (2011). What are the campos rupestres and campos de altitude in the eastern Brazilian mountaintops? Rev. Bras. De Bot. 34:241-246.
|
|
|
|
|
Veloso HP, Rangel Filho ALR, Lima JCA (1991). Brazilian vegetation classification adapted to a universal system. In: Classificação da vegetação brasileira adaptada a um sistema universal. Rio de Janeiro: IBGE p88. Online: View
|
|
|
|
|
Zhang ZQ, Shu WS, Lan CY, Wong MH (2001). Soil seed bank as an input of seed source in revegetation of lead/zinc mine tailings. Restor. Ecol. 9:378-385.
Crossref
|
|