Review

ABSTRACT
Cancer is a leading cause of death worldwide, resulting in 8.2 million or 14.6% of all human deaths worldwide in 2012. More than 70% of all cancer deaths occurred in low and middle income countries; although the risk of developing/dying from it is still higher in the developed regions of the world, and tobacco use is the cause of about 22% of cancer deaths which is still a major public health problem worldwide that requires new strategies and treatment modalities to optimize patient outcomes. However, the perception of the clinical importance of cancer vaccines has been undermined by a history full of well-meaning but mostly failed attempts to treat patients with ill-defined formulations. Growing knowledge of immunology has influenced approaches to vaccine design over the past centuries, producing different types of vaccines with various associated advantages and challenges. The lack of effective active immunotherapy has led to the development of numerous novel strategies such as the common cancer vaccine strategies and designs. These are: protein subunit and peptide vaccine, dendritic cell vaccine, DNA fusion vaccine, antibody directed vaccine, cell based cancer vaccine, recombinant viral and bacterial cancer vaccine. The majority of cancer vaccines aim to induce a cellular antigen-specific T-cell response. Therefore, effective cancer vaccines must resolve several challenges such as cancer vaccines seek to target an antigen specific to the tumor and distinct from self-proteins. Cancer vaccines should require selection of the appropriate adjuvant and also seek to provide long term memory to prevent tumor recurrence, and both the innate and adaptive immune systems should be activated for total tumor elimination.
Key words: Cancer, tumor cells, vaccine strategies.
INTRODUCTION
Growing knowledge of immunology has influenced approaches to vaccine design over the past centuries, producing different types of vaccines with various associated advantages and challenges. These early attempts employed infected material (as there was no knowledge of microorganisms at that time) either by administration of sub-lethal doses (known as ‘variolation’ in the case of small pox) or by using infectious material from a similar infection in animals that was observed not to be harmful to humans, which was basically the strategy employed by Edward Jenner with cowpox. The first smallpox vaccine was inaugurated by Jenner in 1796 after he recognized the value of the accidental observa-tion that milkmaids infected by cowpox were protected against smallpox disease (Worboys, 2007). By delibe-rately inoculating people with small doses of cowpox pustules, Jenner demonstrated that protection against smallpox could be achieved. It is interesting to note that, centuries before Jenner, the Chinese were already applying the principle of variolation to protect people from infection (Plotkin and Plotkin, 2008). While Jenner had no knowledge of microorganisms and viruses, progress in microbiology and virology from the late 19th century onwards provided the modern concept of communicable diseases.
Hence further advances in vaccinology came from understanding the aetiology of infectious diseases and host-pathogen interactions. One important advance was the demonstration that the administration of pathogens, either attenuated (rendered nonpathogenic) or killed (inactivated by heat or chemical denaturation), yielded protection against the disease caused by that pathogen. Impressively, many of the vaccines employed today are still based on these strategies (Zepp, 2010). This indicates, although there has been significant progress in developing cancer vaccines in the last decade, the concept of stimulating the immune system to combat infection is a century old. Coley first used streptococcal cultures to treat patients with advanced sarcoma (a type of malignant cancer of the connective tissue) between 1900 and 1936 (Coley, 1893; Morse et al., 2004). The result was a clinical immune response against the tumor being the first cancer vaccine to be used.
Today we understand that the components of the bacterial extract stimulated the immune response in a general way, causing Coley's toxin to succeed. Since Coley, Bacillus Calmette-Guerin (BCG), which works in a similar manner, has been successfully used against bladder cancer. In the 1980s, Rosenberg used interleukin-2 (IL-2) to treat advanced cancers. Tumor regression was reported in 15 to 20% of the patients. IL-2 does not directly affect solid tumor growth. The anti-tumor effect seen is believed to be due to its effect on T-cells. All of these vaccines provide a general boost to the immune system, enabling it to respond better against the tumor (Zepp, 2010).
Obviously, the classic concept of the vaccine has been derived from the practice of immunizing against infectious agents to prevent disease by generating a humoral immunity. Individuals are immunized against viral or bacterial antigens before they encounter the pathogenic organisms. This strategy became rapidly successful for viruses because viral genes are relatively simple, possessing a limited number of defined antigens. However, in the case of most tumors, there are an unlimited number of potential antigens that can be the target of an immune response. In addition, it is likely that many of these antigens arise during or as a result of the tumorigenesis process. Therefore, when we talk about tumor vaccines, the most common clinical setting is one in which the induction of a systemic immune response by the vaccine occurs subsequent to, rather than before, the antigen insult. Finally, in contrast to prophylactic vaccines, the majority of cancer vaccines aim to induce a cellular antigen-specific T-cell response (Tim and Elizabeth, 1999).
Since the first reported cancer vaccine was reported by Coley (1893), numerous subsequent clinical trials have been conducted, particularly using irradiated whole tumor cells mixed with bacterial adjuvants such as Calmette-Gue´rin bacillus or Corynebacterium parvum. Studies of this approach for patients with malignant melanoma, renal cell and colorectal carcinomas have demonstrated small but significant clinical effects (Hoover et al., 1993). These early studies established a basis for using immune modulators in a paracrine fashion to generate antitumor immunity (Pardoll, 1995). Treating cancer with vaccines has been a challenging field of investigation since the 1950s.
Over the years, the lack of effective active immuno-therapies has led to the development of numerous novel strategies. However, the use of therapeutic cancer vaccines may be on the verge of becoming an effective modality. Recent phase clinical trials have achieved hopeful results in terms of overall survival (Butts et al., 2005). Yet despite these encouraging successes, in general, very little is known about the basic immuno-logical mechanisms involved in vaccine immunotherapy. Gaining a better understanding of the mechanisms that govern the specific immune responses (that is, cytotoxic T lymphocytes, CD4 T helper cells, T regulatory cells, cells of innate immunity, tumor escape mechanisms) elicited by each of the various vaccine platforms, should be a concern of cancer vaccine clinical trials, along with clinical benefits (Vergati et al., 2010). The objective of this review is, therefore, to overview the different strategies of cancer vaccines.
GENERAL OVERVIEW OF CANCER VACCINE
Tumor immunology
It is obvious that the mammalian immune system consists of two broad arms: innate immunity and adaptive immunity. Innate immunity is constitutive, non-specific and swift. It consists of natural anatomical barriers, such as skin and mucous membranes, and physiological barriers like elevation of temperature and acid in the stomach to digest harmful bacteria, and phagocytic cells and their intracellular and extracellular components. Another feature of the innate immune system is comple-ment, which is a group of inactive proteins in the blood. These are activated in the presence of pathogens and cause cell lysis. Tumor cells have complement regulatory proteins on their cell surface that inhibit the activation of complement and thus escape complement-mediated lysis. Pattern-recognition receptors, present on the cell surface, and antimicrobial proteins present inside cells, are part of the innate immunity. Phagocytic cells, namely, natural killer cells, dendritic cells and macrophages are the components of the innate immune system most directly involved in tumor immunology. These cells also participate in the adaptive response and form a bridge between the two arms of the immune system. Innate immunity evolutionarily precedes adaptive immunity and is therefore present in all vertebrate and many invertebrate species. In addition to the phagocytic function of engulfing bacterial and damaged cells, the cells of the innate branch have specific roles (Mansour et al., 2005).
Natural killer (NK) cells: Natural Killer cells are part of innate immunity that possesses the ability to kill tumor cells without a previous encounter. These cells have killer activating receptors and cause lysis of target cells using specialized enzymes, perforin and granzymes. Killer inhibitory receptors are also present on the NK cell surface, which prevent lysis of cells with MHC molecules. Target cells for NK cells include virally-infected cells and tumor cells. NK cells do not require binding to MHC-antigen complex, so they can kill tumor cells that have low levels of MHC molecules. NK cells play a key role in tumor immunology (Biragyn et al., 2001).
Dendritic cells (DCs): When encountered, the pathogens are phagocytosed, their proteins are processed and inserted into the DC surface to be presented to T-cells. Dendritic cells are professional antigen-presenting cells (APC). They activate helper T cells and cytotoxic T cells and also activate B cells. They are very important in tumor immunology. DCs are CD34+, meaning the CD34 differentiation antigen is present on their cell surface (Biragyn et al., 2002).
Macrophages: Macrophages have granules filled with digestive enzymes. They are important in fighting bacteria and also ingest damaged cells by phagocytosis. In addition to being strongly phagocytic, they present antigens to T cells and thus have a role in adaptive immunity. Macrophages can destroy tumor cells and play a crucial role in the inflammatory response (Janeway and Medzehitov, 2002).
Adaptive immunity: The adaptive or acquired responses of the immune system are very specific and slower than the innate response. The adaptive response follows the innate response and is dependent on specific recognition of antigen by antigen receptors present on the cell surface. There are two types of adaptive immunity: cell-mediated immunity and humoral immunity. T lymphocytes are responsible for cell-mediated immunity and B lym-phocytes for humoral immunity. Immunological memory is a feature of adaptive immunity after the initial immune response, B and T memory cells present in the blood are triggered to mount a stronger and more effective immune response when they encounter the same intruder again (Janeway and Medzhitov, 2002).
B-cells: B-cells are produced and mature in the bone marrow. On the cell surface are antigen receptors or B-cell receptors (BCR), proteins that recognize and bind to soluble antigens in the blood. The antigens are then taken up by the cells and processed. The fragments of the digested antigens are displayed on the surface of the cell bound to MHC class II molecules. This induces a T helper cell to bind and secrete lymphokines. The lymphokines cause the B cell to mature and divide into a plasma cell. The mature B-cell switches into an antibody-producing plasma cell. Antibodies bind with very high specificity to the antigen. In tumor immunity, B cells play a role in destroying tumor cells by two different means. The first is complement-mediated lysis. In addition they facilitate antibody-dependent cell-mediated cytotoxicity, in which antibodies recognize and bind to a tumor cell and then trigger cell lysis of target tumor cell by several different immune cells (Ai et al., 2009).
T-cells (T lymphocytes): During hematopiesis, bone marrow stem cells that are destined to become T cells migrate to the thymus to complete their development and maturation. The thymocytes or developing T cells are protected from the contact with antigens in the blood. CD antigen markers are expressed on the T cell surface as they differentiate that is, develop into the specific type of T cells (Antony et al., 2005).
Cytotoxic T cells: CTL T cells are effector T cells. These cells play a central role in tumor immunology since they destroy tumor cells, which they recognize by virtue of the tumor cell surface antigens. Immature CTLs require activation by antigen-presenting cells. Mature CTLs recognize major histocompatibility complex or MHC- antigen complex on the surface of a target cell and destroy the cell. Tumor cells that do not display MHC-antigen complex are destroyed by NK cells. Together CTL and NK cells are the two cell types that destroy tumor cells (Antony et al., 2005).
Helper T cells (Th cells): Helper cells are so called because they help other immune cells to perform their function primarily by secretion of cytokines. Th cells, CD4+ cells in the thymus, are destined to be helper T cells. These, along with CTL, are effector T cells. Presentation of antigen and the appropriate cytokine lead to differentiation into type 1 (Th1) or type 2 (Th2) cells. Th1 cells participate in cell-mediated immunity in controlling infections and Th2 cells participate in humoral immunity since they cooperate with B cells (Cruise and Lewis, 1999).
Suppressor T cells or regulatory T cells (T supp or T reg cells): Suppressor T cells are subpopulation of T cells that prevent an uncontrolled immune response of effector cells, which could lead to autoimmunity. They are mainly of two types; (1) Natural Treg which are CD4+CD25+ and (2) Adaptive Treg which are CD4+, but acquire CD25 and are induced by inflammation, autoimmunity and cancers. T regs function by secretion of TGF-beta, and IL-10 interference of effector cell T cell receptor (TCR) binding to MHC-antigen complex (von Boehmer, 2005).
Natural Killer T (NKT) cells: NKT cells play a role in searching for tumor cells called immune surveillance and in preventing metastasis. These cells are cytotoxic cells that have characteristics of both NK cells and T cells. NKT cells express TCR (a specific subtype of TCR not found in other T cells) on their cell surface, and in addition express NK receptors, making them unique since they display receptors of both NK cells and T cells. NKT cells rapidly release cytokines like IL-4 or gamma-interferon and are able to activate T helper cells, making NKT cells a link between innate and adaptive immunity (Janeway and Medzhitov, 2002; Franco, 2005).
Antigen presentation and T cell activation: When cells in the body present MHC class I coupled antigens, T cells recognize and destroy the cells. However, T cells do not recognize many tumor antigens directly on the cells. Instead, special antigen-presenting cells (APCs) present the antigen for recognition, which triggers activation of T cells, enabling the T cells to mount a response to destroy the tumor cells. Dendritic cells are proficient at processing and presenting antigens and are thus called professional antigen-presenting cells. DCs have high levels of major histocompatibility complex (MHC), costimulatory molecules and cell adhesion molecules all of which are essential for antigen presentation (Armstrong et al., 2001).
The function of MHC molecules, named for immune rejection of incompatible transplanted tissue, is to play a vital role in cell-mediated immunity. When there are problems within cells, such as an infection or break down of cellular material prior to cell death, the degraded proteins of the cell are displayed on the cell surface bound to the MHC molecule. These protein fragments or antigens are displayed and recognized by circulating immune cells in the blood, allowing the immune system to survey the health status of cells since healthy cells would not have the degraded protein and the fragments displayed on the cell surface. MHC class I molecules are displayed on almost all the cells of the body and are recognized by CTL cells. MHC class II molecules are displayed on immune cells, namely dendritic cells and macrophages, and are recognized by helper T cells (Armstrong et al., 2001).
The MHC molecules are responsible for self-tolerance and mounting an immune response against foreign and potentially harmful agents. Antigen presentation leads to activation of T cells that can recognize and destroy tumors.
MHC class II-antigen complex on the APC cell surface is recognized by the T cell antigen receptor (TCR). The binding of the two is very specific and is called an immunological synapse. Consequently, the costimulatory receptor, CD28, binds DC cell adhesion molecules. The formation of the TCR-MHC-antigen complex sets off a cascade of events within the T cell resulting in gene activation, and the ultimate result is cell proliferation or differentiation or anergy or apoptosis. Cytokines, specifically IL-2 and also other interleukins, play an important role in cell activation. Activation of CTL and Th cells is similar. Differentiation of the T cell into an effector cell (a T helper cell or CTL) is required to produce a cell-mediated immune response. It is noteworthy that T cell activation is a complex event; failure to obtain a sustainable activation signal results in failure to recognize the tumor antigen, leading to failure to attack the tumor (Nel, 2002).
Role of cytokines
Cytokines are essential in any type of immune response. These are proteins secreted by cells of the immune system to signal other cells of the immune system. Typically they are produced locally and affect cells in the near vicinity. The following table lists some of the cytokines involved in generating an immune response (Biragyn et al., 2001).
Immune properties of tumors
Tumors do not induce a strong immune response for two major reasons. First, early in embryonic development, all the lymphocytes that recognize normal cells' antigens, "self antigens", are destroyed to prevent an immune response against one's own tissue called “self tolerance”. Since tumor cells are not dramatically different from normal cells, tumor cells are also immunologically tolerated that is, ignored by the immune system. Second, tumors actively evade the immune system. To examine the immune properties more closely this section is subdivided into three: tumor antigens, the mechanism of immune evasion by tumors and the detection of cancerous growth called “immune surveillance” (Zitvogel et al., 2004).
Tumor antigens
In general, tumors are poorly immunogenic and do not trigger an immune response (Igney and krammer, 2002). There are two types of tumor antigens. Some tumors have unique antigens that they are not found on normal cells, called tumor-specific antigens (TSA) (Robbins, 2004). TSA are present in tumors induced by infectious agents (for example, EBNA-1 antigen from Epstein Barr virus-induced Burkitt's lymphoma) and mutated genes found only in tumor cells for example, mutated caspase-8 enzyme found in head and neck cancer, which is different from the normal caspase-8). However, many tumors have antigens, called tumor-associated antigens (TAA), similar to antigen in normal body cells but, either modified or produced in greater quantities. Some TAA are limited to a specific tumor type for example, melanocyte differentiation antigens, MAGE antigens, are limited to melanomas and some normal tissue, while other TAA are found in several different tumors for example, cancer testis antigen are found in normal testis and a variety of cancers such as prostate carcinoma, breast carcinoma, and lung carcinoma (Bellamy et al., 1995).
Immune evasion of tumors
During the last decade, increased understanding of the molecular mechanisms responsible for immune activation to protect against challenges by tumor cells has revolutionized the field of immunotherapy research. It has been demonstrated that the dysfunction of the host's immune system represents one of the major mechanisms by which tumors evade immune-surveillance. This is due, for example, to T cell anergy, the existence of regulatory T cells, and systemic defects of dendritic cells derived from tumor patients. In addition, escape from immune surveillance can also be linked to tumor-related factors, including secretion of immune-suppressive cytokines, resistance to apoptosis, and deficient expression of immune-modulatory molecules and major histocompatibility complex (MHC) class I antigens possibly due to immune-selection. Both host- and tumor-related mechanisms can lead to a failure to mount a proper anti-tumor-specific immune response, and these are frequently key factors in limiting the success of cancer immunotherapy (Seliger, 2005).
Immune surveillance
Like humans, immune-competent animals are capable of recognizing and eliminating syngeneic cancer cells not only before the development of a tumor but also after tumor formation, the so-called “cancer immune surveillance” (Dunn et al., 2002). T cells (NKT cells and CTLs) are believed to carry out an immune surveillance function. They seek out newly transformed cells by recognizing tumor antigens. The tumor antigens do not need to be on the cell surface, since antigen processing and presentation coupled to MHC I molecules ensures that even intracellular antigens are recognized and the cells are destroyed. Tumor-specific CTL have been observed in many types of cancers. NK cells are also found in tumors and appear to recognize common characteristics of tumor cells. Humoral immune response is also involved in detecting and destroying cancers. As an example, carcinogen-induced and virus-induced leukemia is recognized by antibodies. These antibodies trigger complement-mediated cytotoxicity. Despite the fact that immune cells recognize tumors and are found within them, tumors do occur. Growth of a tumor can therefore be viewed as failure of immune surveillance (Uzzo et al., 2003).
CANCER VACCINE DESIGN STRATEGIES
One reason for the limited performance in some vaccine trials is likely to be the vaccine formulation. For weak immunogens, strategies to induce immunity should take into account immunologic principles established by the use of mouse models have to be carefully sifted because, unlike human cancer, most mouse tumor cell lines carry retroviruses, and retroviral proteins can act as strong tumor-associated antigens (Huang et al., 1996). A clear principle is the requirement for activation of innate immunity, which begins the process of immune activation. This was illustrated in patients with melanoma, in whom dramatic enhancement of the immunogenicity of a peptide incomplete Freund’s adjuvant vaccine against melanoma was observed when a CpG oligonucleotide, capable of triggering dendritic cells by way of Toll-like receptor-9, was included (Peiser et al., 2005). A further principle is the need for CD4+ helper (TH) cells to support both antibody and CTL responses (Janssen et al., 2003). This is particularly important in situations where a natural CD4+TH response is being dampened by regulatory T cells (Antony et al., 2005). It is highly unlikely that vaccination with a tumor-derived protein or peptide alone will induce and maintain the high levels of effector pathways required to suppress cancer. This might account for the low responses observed in some clinical trials. For trials of vaccination of patients with lymphoma using idiotypic immunoglobulin protein antigens, fusion with keyhole limpet hemocyanin (KLH) has generally been used. This ensures activation of TH cells and has led to regular induction of immunity, with promising clinical effects (Weng et al., 2004).
Until recently, the main distinction between vaccination against infectious diseases and against cancer has been in the clinical setting. The requirements for induction of effective immunity against cancer therefore are to present tumor antigens to the immune system in a way that will break tolerance, if it exists, and to activate high levels of effector pathways able to suppress tumor growth on a continuing basis. To achieve this goal, a variety of strategies are currently being investigated both in preclinical models and in clinical trials (Figure 1) (Stevenson et al., 2004).
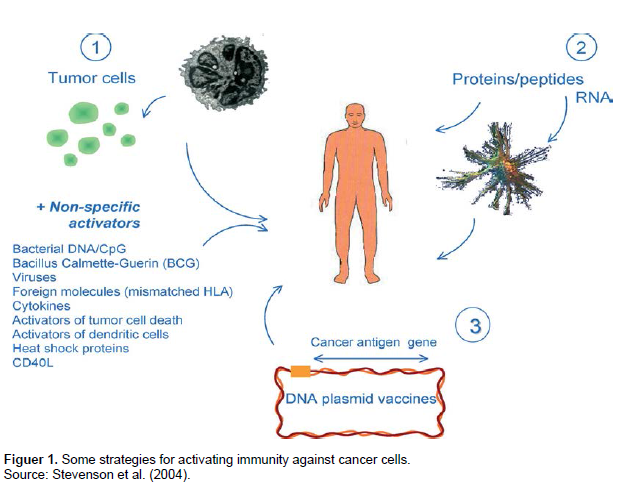
It is now clear that there are two barriers that have to be overcome to activate the immune response. The first is to engage the attention of the patrolling DCs, so that, following antigen uptake, the DC migrates to the draining lymph node and prime naïve CD4 T cells. Non-specific activation alone was used initially as a means of inducing immunity against cancer, most notably with ‘Coley’s toxin’ whereby infection or injection of a bacterial extract could reduce tumor growth (Wiemann and Starnes, 1994). A more refined version of this strategy is to use oligonucleotides containing sequences characteristic of bacterial DNA, the so called CpG motifs (Krieg, 2002).
Bacterial DNA contains immunostimulatory motifs that trigger an innate immune response characterized by the production of predominantly TH1-type cytokines. These motifs consist of an unmethylated CpG dinucleotide flanked by two 5' purines and two 3' pyrimidines. Synthetic oligodeoxynucleotides (oligos) expressing these motifs would act as adjuvants to boost the immune response to DNA- and protein-based immunogens. In vivo experiments demonstrate that CpG-containing oligos augment antigen-specific serum antibody levels by up to tenfold, and IFNgamma production by up to sixfold. These effects were optimized by physically linking the CpG-containing motifs to the immunogen (Klinman et al., 1999).
A wide range of activators of the immune system has been tested against a variety of tumors. For hematological malignancies, stimulation with CpG oligonucleotides and exposure to activating molecules such as CD40 ligand have been used to encourage cancer cells to present intrinsic tumor antigens (Rieger and Kipps, 2003). The attraction is that there is no need to identify the antigen, but this approach is partly countered by the difficulty of measuring specific immune responses. Modeling in animals of non-specific stimulation of immunity is also a problem, because this approach can activate specific immunity against retroviral antigens. While these antigens are known to be present in murine tumors, they are less likely to be as common in human tumors (Kershaw et al., 2001). Efficacy in preclinical models may, therefore, not be relevant for human cancer, and it is highly desirable to move into pilot clinical trials to test this approach (Rieger and Kipps, 2003) or granulocyte–macrophage colony-stimulating factor (GMCSF). An alternative is to directly introduce antigens into DCs by loading antigen in vitro using protein or peptide or RNA (Chang and Dhodapkar, 2003). Adoptive transfer of loaded DCs shows promise, with data accumulating from clinical trials (Timmerman et al., 2002). However, the DC system is complex and heterogeneous, and more refinement will be needed to optimize the approach.
Proteins and peptides based vaccine
The use of proteins or peptides to stimulate a specific immune response against cancer has long been investigated and covers a broad spectrum of possibilities employing single agents or combinations of proteins, heat-shock proteins (HSPs) (Bolhassani and Rafati, 2008), peptides and agonist peptides (Cereda et al., 2010) antiidiotype antibodies (Ai et al., 2009), and fusion proteins (So-Rosillo and Small, 2006).
These protein- or epitope-based vaccines have two main advantages over the use of tumor cells or lysates. First, production, storage, and distribution are faster and more cost-effective, and second, the identification and administration of TSAs is preferable since tumor-cell preparations mostly contain self-proteins with no therapeutic benefit and are potentially capable of generating an autoimmune response. On the other hand, this approach has certain drawbacks. Primarily, it is the weak immunogenicity of a single protein or, especially, a single epitope. Secondly, tumors can easily escape immune recognition through antigen mutation or loss. Thirdly, their use is HLA restricted (mainly for epitope-based vaccines) and limited to a subset of patients (usually HLA-A2+). Fourthly, they have a poor ability to induce balanced activation of CD4 and CD8 subsets, which is thought to be essential for effective, long-lasting antitumor immunity. Protein-based vaccines are capable of generating stronger CD4 responses (MHC class II-restricted), but at the cost of less effective induction of CTLs (Kalinski et al., 2009). Most of the issues described could be easily overcome by the use of longer peptides or the combination of several different epitopes in the same vaccine, while the relatively poor immunogenicity of peptides could necessitate that they are administered with adjuvants or loaded onto DCs (Lesterhuis et al., 2004).
The use of specific proteins or peptides as targets for immunotherapy clearly requires a careful choice of the targeted TAAs and their epitopes, involving knowledge of their structural and functional characteristics. Single-peptide epitopes composed of 8 to 10 amino acids are able to induce a CTL response by binding to MHC class I molecules expressed on APCs. Each epitope is com-posed of conserved anchor residues (mostly at position 2 and the C-terminal position) needed to bind to the cleft of MHC I molecules and residues that are specific for T-cell recognition. Theoretically, changes in the former do not affect the specificity of the latter, and they have been used as a strategy to increase the immunogenicity of several different epitopes (agonist epitopes) (Yokokawa et al., 2007). Furthermore, the ideal TAA should be widely expressed in different tumor types and also play a central role in oncogenic processes or in cancer cell survival, to avoid immune escape by mutations or loss of antigens by tumor cells. Identification of novel TAAs can be achieved through 2 experimental processes. These are direct immunology (starting from patient-derived auto-logous tumor-specific CTL clones specific for an unknown epitope), and reverse immunology (starting from a predicted epitope). The former has been used since the discovery of the first tumor-specific CTL epitope, MAGE-1 (Kessler and Melief, 2007). Direct immunology is further subdivided into genetic or biochemical approaches.
Briefly, in the genetic approach, a patient-derived CTL clone is screened by using target cells transfected with tumor derived cDNA libraries. Subsequently, the increased release of cytokines in the supernatant due to the recognition by the tumor-specific CTL clone allows one to select the cells that contain the antigen-encoding cDNA. These are then subcloned and rescreened to finally identify the cDNA that encodes the specific antigen. The biochemical approach consists of the puri-fication of peptides eluted from MHC class I molecules of antigen-expressing cells by high-performance liquid chromatography fractionation. Antigen-negative target cells expressing the appropriate HLA molecule are used to load these peptides and tested for CTL recognition. Positive fractions are analyzed by mass spectrometry to identify the amino acid sequence of the epitope recognized by CTLs (Kessler and Melief, 2007).
The need for expensive specialized equipment, plus the labor-intensive method, probably accounts for the increasing use of reverse immunology. Over the years, a growing understanding of HLA specific peptide-binding motifs has led to the development of several computer algorithms for amino acid sequences with predicted binding capacity. Reverse immunology consists of two different phases. The first phase is the epitope prediction phase in which proteins are analyzed for the presence of potential epitopes by the use of prediction algorithms. The next is the epitope validation phase in which the candidate peptides are tested by binding and stability assays in vitro. Nevertheless, differences between the processing machinery in normal and tumor cells might be liable for the lack of activity against tumor cells of several CTLs raised against high-affinity binding TAAs (Campoli and Ferrone, 2008).
Nowadays, indeed, the most recent algorithms also take into account the proteasomal processing and transporters associated with antigen processing- (TAP-) translocations are other fundamental processes in the antigen-presentation pathway. Despite many efforts, the use of epitope-based vaccines has not advanced beyond phase I or II clinical trials, probably due to the drawbacks described. To date, the best results have been achieved with the use of fusion protein- or HSP-based vaccines. Provenge (sipuleucel-T, Dendreon Corporation) is in late-stage development for the treatment of mHRPC Sipuleucel-T is an immunotherapy product designed to stimulate T-cell immunity against prostatic acid phosphatase (PAP). It consists of autologous APCs isolated by leukapheresis, cultured with a prostatic acid phosphatase granulocyte-macrophage colony-stimulating factor (PAP-GM-CSF) fusion protein, and reinfused into the patient. The time from apheresis to infusion of final product is approximately 48 h (Small et al., 2006; Higano et al., 2009).
The appropriate tumor antigen is bound to the DC cell surface. The tumor antigens are taken up by dendritic cells, and are processed and presented to the T-cells along with the appropriate costimulatory signal. Once activated by the DCs the cytotoxic T cells recognize and destroy the tumor cells expressing the tumor antigen. DCs are collected from the blood of the patient (a process called leukapheresis) and "loaded" with tumor antigens from the patient's own tumor cells. These DCs are then reintroduced into the patient and stimulate the immune system. DC vaccines have been used in patients with metastatic melanoma, renal carcinoma and prostate cancer (Pardoll, 2008).
Loading of dendritic cells could circumvent the need for initial activation of innate immunity, but the need for T-cell help is likely to remain, however, trials of dendritic cells loaded with protein, peptides, or tumor-derived RNA have shown some clinical effects. Prostate cancer is a useful test bed for vaccine trials because patients tend to be immunocompetent and there are several lineage-specific antigens as potential targets or as monitors of response. In more than 100 dendritic cell-based vaccine trials, a range of immune outcomes and clinical effects has been reported (Ragde et al., 2004). Transfecting dendritic cells with mRNA encoding tumor antigens is another approach being tested for prostate cancer and renal cancer. This allows amplification from small biopsy specimens and is apparently capable of inducing specific T-cell responses, and suggestive clinical responses, in both cancers (Gilboa and Vieweg, 2004).
DNA fusion vaccine
The ease of DNA manipulation has offered an opportunity to develop gene fusion vaccine strategies in which antigens are linked with various immunoenhancing elements. In principle, all pathways of the immune system can be subject to manipulation as outlined in Figure 2. Components to activate innate immunity may be added to enhance the antigen-presenting capacity of DCs. Antigens may be attached to molecules that speci-fically bind ligands on DCs for targeted antigen delivery and cross-presentation. Molecules that are important players of the antigen processing and presentation machinery may be incorporated to direct induction of a specific type of immune response appropriate for the antigen of interest. Finally, activation of cognate CD4+ helper T cells by linking the antigen to highly immuno-genic sequences has proved to be critically important for tumor vaccines to increase antigen immunogenicity and to overcome tolerance (Stevenson et al., 2004).
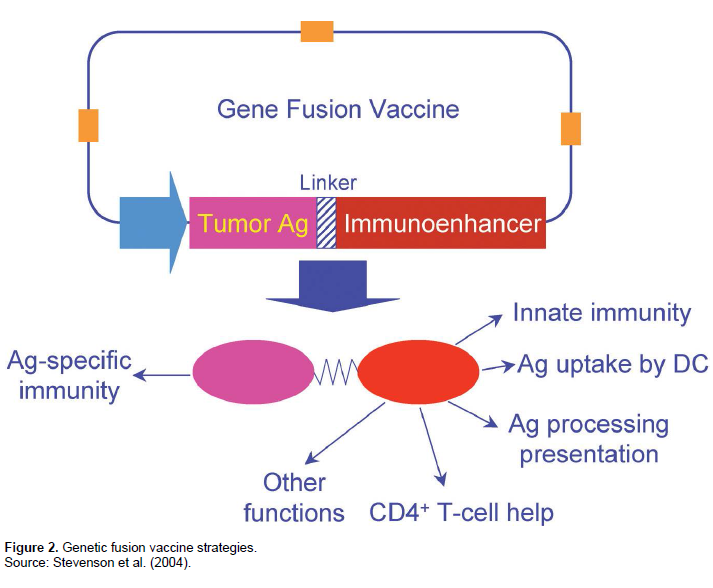
It is becoming increasingly clear that the innate immune system not only provides the first line of defense against invading pathogens but also plays a key regulatory role in the activation of adaptive immunity (Janeway and Medzhitov, 2002). For vaccine purposes, inclusion of components of innate immunity can have an impact on both the magnitude and the nature of the adaptive immune response. Co-delivery of pro-inflammatory cyto-kines and chemokines may improve activation of antigen-specific immunity by recruiting and activating DCs that play a pivotal role in the induction of adaptive immunity. Furthermore, fusion of these molecules directly to antigens may be more effective, because cytokine or chemokine receptor-mediated uptake may additionally result in more efficient antigen presentation by DCs (Biragyn et al., 2001).
One interesting set of molecules associated with the innate immune system is the b-defensins, which are being exploited to increase the potency of DNA vaccines against cancer. Defensins are a family of small cationic peptides abundantly expressed by cells and tissues that are involved in host defense against microbial infection. These peptides can mediate direct killing of bacteria, fungi, and viruses (Ganz, 2003). Moreover, it has recently been shown that human b-defensins are also chemotactic for immature DCs and memory T cells (Yang et al., 1999), while murine b-defensin-2 acts directly on immature DCs via TLR-4, inducing upregulation of costimulatory molecules and DC maturation (Biragyn et al., 2002). Thus, defensins play a role in bridging innate immunity and adaptive immune responses. DNA fusion vaccines consisting of weakly immunogenic B-cell Id antigens and murine b-defensin-2 could induce protective and therapeutic immunity against two different syngeneic mouse lymphomas (Biragyn et al., 2002). Importantly, both chemotactic activities and covalent linkage are required, because neither fusion of inactive prob- defensin nor co-immunization with a mixture of plasmids encoding unlinked Id antigens and b-defensin-2 were effective. These results suggest that targeting the delivery of antigens to immature DCs may be a general approach for increasing the efficacy of DNA vaccines.
Antibody-directed vaccines
Apart from direct treatment of cancer with tumor-specific antibodies that are often used to localize various toxins at the tumor site, different active vaccination strategies are being investigated to induce tumor-specific antibody responses. For example, vaccine strategies for the treatment of melanoma were tested using either GM2-expressing melanoma cells or vaccines containing GM2 conjugated to keyhole limpet hemocyanin. These studies demonstrated that vaccines containing purified GM2 ganglioside result in induction of GM2 antibodies, and high titers of GM2 antibodies were correlated with increased survival (Kitamura et al., 1995). One idiotypic vaccine study, which involved the immunization of non- Hodgkin’s B-cell lymphoma patients with the unique idiotype of the immunoglobulin expressed on the surface of their tumors, demonstrated a clinical benefit for patients with recurrent B-cell lymphomas (Hsu et al., 1997). Furthermore, in a preclinical study, an idiotypic immunization with an antibody specific for mutated p53 prevented the development of a p53 mutation–bearing tumor cell line in mice (Ruiz et al., 1998). These encouraging results strongly support the further clinical development of these approaches.
Cell-based cancer vaccines
Until the tumor antigens expressed by most tumors are identified, the tumor cell itself will continue to be the best source of immunizing antigens. With the development of improved genetic techniques, the concept of presenting immunologically defined ‘‘adjuvants’’ at the same site as tumor antigens in order to augment antitumor immunity has been tested more directly. In animal models, gene transfer of genes encoding MHC molecules, costimulatory molecules, and cytokines has been studied (Townsend and Allison, 1993). Early animal studies evaluated the immune effects of enhanced expression of both autologous and allogeneic MHC class I molecules on the tumor cell surface. Both ex vivo and in vivo gene transfers of these molecules to the tumor have been studied and have met with some success (Plautz et al., 1993) similar vaccine strategies are undergoing clinical testing.
Genes that encode cytokines are the most common types of genes that have been introduced into tumor cells. The cytokine is produced at very high concen-trations at the vicinity of the tumor, thereby altering the local immunologic environment of the tumor cell so as to either enhance presentation of tumor-specific antigens to APCs or to enhance the activation of tumor-specific lymphocytes (Pardoll, 1995). Many cytokine genes have been introduced into tumor cells with varying effects on both tumorigenicity and immunogenicity (Dranoff et al., 1995). The level of cytokine expression, location of immunization, and challenge site are crucial parameters affecting vaccine efficacy for any form of genetically engineered tumor vaccine.
Many of these cytokines have been studied as single cytokines in one or more tumor vaccine models. How-ever, for a human tumor vaccine to be developed, it is critical that these cytokines be compared head to head to determine which cytokine or cytokines are most effective.
Also, given that most mouse tumors show significant immunogenicity when simply irradiated, identification of genes that truly enhance the tumor’s immunogenicity significantly above that of irradiated wild-type tumor cells is important. Only one study has directly compared multiple cytokines and other genes in murine tumor models (Dranoff et al., 1995). This study demonstrated, in a number of poorly and moderately immunogenic tumors, that immunization with the tumors transduced with the cytokine granulocyte-macrophage colony-stimulating factor (GM-CSF) produced the greatest degree of systemic immunity, which was enhanced relative to irradiated non-transduced tumor cells. In vivo depletion of T-cell subsets demonstrated that this immunity was de-pendent on both CD4+ and CD8+ T cells, despite the fact that the tumors did not express MHC class II antigens (Hamilton et al., 2006) importantly; tumor cells genetically altered to express GM-CSF were able to cure mice of pre-established small burdens.
Recombinant viral and bacterial vaccines
With the current knowledge of augmenting antitumor immune responses, the ideal antigen delivery vector should directly infect APCs in vivo and facilitate antigen delivery to both MHC class I and II antigen–processing pathways. Several recombinant viral vectors are currently undergoing rigorous testing for their ability to augment antitumor immune responses against model tumor antigens with particular interest are vaccinia and other pox viruses and Listeria monocytogenes (Greten and Jaffee, 1999). Pox viruses are attractive candidates for the expression of tumor-associated antigens because heterologous proteins are delivered to the cytoplasm, and therefore, are directly targeted to the compartment in which processing of MHC class I antigens is initiated for presentation to CD8+ CTLs. Restifo et al. (1995) have published several studies demonstrating the generation of antigen-specific immunity resulting in the protection against tumor challenges, using vaccinia and fowlpox constructs (Bronte et al., 1995).
Wu et al. (1996) more recently demonstrated the enhanced potency of a vaccinia vector carrying the HPV gene E7 fused with the LAMP1 gene, which targets E7 to the MHC class II antigen–processing pathway for presentation to CD4+ T cells (Wu et al., 1995). In one study, the cure of a significant tumor burden was demonstrated (Lin et al., 1996). A recombinant E6- and E7-expressing vaccinia virus has already been tested in eight patients with late-stage cervical cancer. No significant side effects were described. An HPV-specific CTL response was observed in one of the eight treated patients. However, all eight patients mounted an antivaccinia antibody response (Borysiewicz et al., 1996). These neutralizing antivirus antibodies represent one of the major barriers to the use of viral cancer vaccines, which can be the result of either previous immunizations (as in the case with vaccinia) or exposure to cross-reactive viruses (as in the case with adenoviral vaccines). It is unlikely that recombinant viral vaccines such as vaccinia will reach broad clinical application until methods have been developed to eliminate neutralizing antibodies or new viral vectors are identified that do not induce significant antibody responses. L monocytogenes is a novel tumor antigen delivery vector that is particularly interesting because of its two-phase intracellular life cycle (Falkow et al., 1992) which enables it to deliver antigens to the class I and class II pathways. Recent data have demonstrated the efficacy of using L monocytogenes as a live recombinant vaccine that is sufficiently potent to cause regression of established macrometastases (Pan et al., 1995).
CONCLUSION AND RECOMMENDATION
There are currently no vaccines able to prevent cancer in general. Some types of cancer, such as cervical cancer and some liver cancers, are caused by viruses (known as oncoviruses), and traditional vaccines against those viruses, such as HPV vaccine and hepatitis B vaccine, will prevent those types of cancer. Scientists have also been trying to develop vaccines against existing cancers. Some researchers believe that cancer cells routinely arise and are destroyed by the healthy immune system cancer forms when the immune system fails to destroy them. However, therapeutic cancer vaccines are being developed for the treatment of breast, lung, colon, skin, kidney, prostate, and other cancers. One approach to cancer vaccination is to separate proteins from cancer cells and immunize cancer patients against those proteins, in the hope of stimulating an immune reaction that could kill the cancer cells and another approach to therapeutic anti-cancer vaccination is to generate the immune response in situ in the patient using oncolytic viruses. Therefore, effective cancer vaccines must resolve several challenges. Cancer vaccines seek to target an antigen specific to the tumor and distinct from self-proteins. Selection of the appropriate adjuvant, molecules that activate antigen-presenting cells to stimulate immune responses, is required and also should seek to provide long term memory to prevent tumor recurrence. Both the innate and adaptive immune systems should be activated for total tumor elimination.
CONFLICT OF INTEREST
The authors declare that they have no conflict of interest.
REFERENCES
|
Ai WZ, Tibshirani R, Taidi B, Czerwinski D, Levy R (2009). Anti-idiotype antibody response after vaccination correlates with better overall survival in follicular lymphoma. Blood 113:5743–5746. Crossref |
||||
|
Antony PA, Piccirillo CA, Akpinarli A (2005). CD8+ T cell immunity against a tumor/self-antigen is augmented by CD4+ T helper cells and hindered by naturally occurring T regulatory cells. J. Immunol. 174:2591-2601. Crossref |
||||
|
Armstrong AC, Easton D, Ewing JC (2001). Cellular Immunotherapy for Cancer. Br. Med. J. 323:1289-1293. Crossref |
||||
|
Bellamy CO, Malcomson RD, Harrison DJ, Wyllie AH (1995). Cell death in health and disease: the biology and regulation of apoptosis. Semin. Cancer Biol. 6(1):3–16. Crossref |
||||
|
Biragyn A, Ruffini PA, Leifer CA, Klyushnenkova E, Shakhov A, Chertov O, Shirakawa AK, Farber JM, Segal DM, Oppenheim JJ, Kwak LW (2002). Toll-like receptor 4- dependent activation of dendritic cells by betadefensin 2. Science 298(5595):1025–1029. Crossref |
||||
|
Biragyn A, Tani K, Grimm MC, Weeks S, Kwak LW (2001). Mediators of innate immunity that target immature, but not mature, dendritic cells induce antitumor immunity when genetically fused with nonimmunogenic tumor antigens. J. Immunol. 167:6644–6653. Crossref |
||||
|
Bolhassani A, Rafati S (2008). Heat-shock proteins as powerful weapons in vaccine development. Expert. Rev. Vaccines 7:1185–1199. Crossref |
||||
|
Borysiewicz LK, Fiander A, Nimako M, Man S, Wilkinson GW, Westmoreland D, Evans AS, Adams M, Stacey SN, Boursnell ME, Rutherford E, Hickling JK, Inglis SC (1996). A recombinant vaccinia virus encoding human papillomavirus types 16 and 18, E6 and E7 proteins as immunotherapy for cervical cancer. Lancet 347:1523-1527. Crossref |
||||
|
Butts C Murray N, Maksymiuk A (2005). Randomized phase IIB trial of BLP25 liposome vaccine in stage III B and IV non-small-cell lung cancer. J. Clin. Oncol. 23(27):6674–6681. Crossref |
||||
|
Bronte V, Tsung K, Rao JB (1995). IL-2 enhances the function of recombinant poxvirus-based vaccines in the treatment of established pulmonary metastases. J. Immunol. 154:5282-5292. Pubmed |
||||
|
Campoli M, Ferrone S (2008). HLA antigen changes in malignant cells: epigenetic mechanisms and biologic significance. Oncogene 27:5869–5885. Crossref |
||||
|
Cereda V, Poole DJ, Palena C (2010). New gene expressed in prostate: a potential target for T cell-mediated prostate cancer immunotherapy. Cancer Immunol. Immunother. 59(1):63–71. Crossref |
||||
|
Chang DH, Dhodapkar MV (2003). Dendritic cells and immunotherapy for cancer. Int. J. Hematol. 77:439–443. Crossref |
||||
|
Coley WB (1893). The treatment of malignant tumors by repeated inoculations of erysipelas: With a report of ten original cases. Am. J. Med. Sci. 105:487-511. Crossref |
||||
| Cruise JM, Lewis RE (1999). The Thymus and T Lymphocytes, Atlas of Human Immunology. pp. 161-183. | ||||
|
Dranoff G, Jaffee E, Lazenby A (1995). Vaccination with irradiated tumor cells engineered to secrete murine granulocyte-macrophage colony-stimulating factor stimulates potent, specific, and long-lasting anti-tumor immunity. Proc. Natl. Acad. Sci. 90:3539-3543. Crossref |
||||
|
Dunn GP, Bruce AT, Ikeda H, Old LJ, Schreiber RD (2002). Cancer immuno-editing: from immunosurveillance to tumor escape. Nat. Immunol. 3:991–998. Crossref |
||||
|
Falkow S, Isberg RR, Portnoy D (1992). The interaction of bacteria with mammalian cells. Annu. Rev. Cell Biol. 8: 333-363. Crossref |
||||
|
Franco A (2005). CTL-Based cancer Preventive therapeutic Vaccines for Carcinomas: Role of Tumour-Associated Carbohydrate Antigens. Scand. J. Immunol. 61:391-397. Crossref |
||||
|
Ganz T (2003). Defensins: antimicrobial peptides of innate immunity. Nat. Rev. Immunol. 3:710–720. Crossref |
||||
|
Gilboa E, Vieweg J (2004). Cancer immunotherapy with mRNA-transfected dendritic cells. Immunol. Rev. 199: 251-263. Crossref |
||||
|
Greten TF, Jaffee EM (1999). Cancer Vaccines. American Society of Clinical Oncology. J. Clin. Oncol. 17:1047-1060. Pubmed |
||||
|
Hamilton SE, Wolkers MC, Schoenberger SP, Jameson SC, (2006). The generation of protective memory-like CD8+ T cells during homeostatic proliferation requires CD4+ T cells. Nat. Immunol. 7(5):475–481. Crossref |
||||
|
Higano CS, Schellhammer PF, Small EJ (2009). Integrated data from two randomized, double-blind, placebo controlled, phase 3 trials of active cellular immunotherapy with sipuleucel-T in advanced prostate Cancer. Cancer 115:3670–3679. Crossref |
||||
|
Hoover HJ, Brandhorst JS, Peters LC (1993). Adjuvant active specific immunotherapy for human colorectal cancer: 6.5-Year median follow-up of a phase III prospectively randomized trial. J. Clin. Oncol. 11:390-399. Pubmed |
||||
|
Hsu FJ, Caspar CB, Czerwinski D, Kwak LW, Liles TM, Syrengelas A, Taidi-Laskowski B, Levy R (1997). Tumor-specific idiotype vaccines in the treatment of patients with B-cell lymphoma: Long-term results of a clinical trial. Blood 89:3129-3135. Pubmed |
||||
|
Huang AY, Gulden PH, Woods AS (1996). The immunodominant major histocompatibility complex class I-restricted antigen of a murine colon tumor derives from an endogenous retroviral gene product. Proc. Natl. Acad. Sci. 93:9730-9735. Crossref |
||||
|
Igney FH, Krammer PH (2002). Death and anti-death: tumour resistance to apoptosis. Nat. Rev. Cancer 2:277–288. Crossref |
||||
|
Janeway CA, Medzhitov R (2002). Innate immune recognition. Annu. Rev. Immunol. 20:197–216. Crossref |
||||
|
Janssen EM, Lemmens EE, Wolfe T (2003). CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes. Nature 421:852-856. Crossref |
||||
|
Jemal R, Siegel E, Ward Y, Hao J, Xu, TM (2009). Cancer statistics, 2009. CA Cancer J. Clin. 59:225–249. Crossref |
||||
|
Kalinski P, Urban J, Narang R, Berk E, Wieckowski, Muthuswamy R (2009). Dendritic cell-based therapeutic cancer vaccines: what we have and what we need. Future Oncol. 5:379–390. Crossref |
||||
|
Kershaw MH, Hsu C, Mondesire W, Parker LL, Wang G, Overwijk WW, Lapointe R, Yang JC, Wang RF, Restifo NP, Hwu P (2001). Immunization against endogenous retroviral tumor-associated antigens. Cancer Res. 61(21):7920–7924. Pubmed |
||||
|
Kessler J, Melief CJ (2007). Identification of T-cell epitopes for cancer immunotherapy. Leukemia 21(9):1859–1874. Crossref |
||||
|
Kitamura K, Livingston PO, Fortunato SR (1995). Serological response patterns of melanoma patients immunized with a GM2 ganglioside conjugate vaccine. Proc. Natl. Acad. Sci. 92:2805-2809. Crossref |
||||
|
Krieg AM (2002). CpG motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 20:709–760. Crossref |
||||
|
Klinman DM, Barnhart KM, Conover J (1999). CpG motifs as immune adjuvants. Vaccine 17(1):19-25. Crossref |
||||
| Lesterhuis WJ, de Vries IJ, Adema GJ, Punt C (2004). Dendritic cell-based vaccines in cancer immunotherapy: an update on clinical and immunological results. Ann. Oncol. 15:145–151. | ||||
| Lin KY, Guarnieri FG, Staveley-O'Carroll K (1996). Treatment of established tumors with a novel vaccine that enhances major histocompatibility class II presentation of tumor antigen. Cancer Res. 56:21-26. | ||||
|
Mansour W, Gilham DE, Thistlethwaite FC, Hawkins RE (2005). Engineering T Cells for Cancer Therapy. Br. J. Cancer 93(10):1085-1091. Crossref |
||||
|
Morse MA, Clay TM, Lyerly HK (2004). Biology of the Cancer Vaccine Immune Response. Handbook of Cancer Vaccines. Pp. 3-17. Crossref |
||||
|
Nel AE (2002). T-cell Activation through the Antigen Receptor. Part 1: Signaling components, Signaling Pathways, and Signal Integration at the T-cell Antigen Receptor Synapse. J. Allergy Clin. Immunol. 109:758-770. Crossref |
||||
|
Pan ZK, Ikonomidis G, Lazenby A (1995). A recombinant Listeria monocytogenes vaccine expressing a model tumour antigen protects mice against lethal tumour cell challenge and causes regression of established tumours. Nat. Med. 1:471-477. Crossref |
||||
|
Pardoll DM (1995). Paracrine cytokine adjuvants in cancer immunotherapy. Annu. Rev. Immunol. 13:399-415. Crossref |
||||
|
Pardoll DM (2008). Cancer immunology. In: Abeloff MD, Armitage JO, Niederhuber JE, Kastan MB, McKenna WG, (eds.), Abeloff's Clinical Oncology 4th ed. Philadelphia: Churchill Livingstone. pp. 140-147. Crossref |
||||
|
Peiser DE, Lienard D, Rufer N (2005). Rapid and strong human CD8+ T cell responses to vaccination with peptide, IFA, and CpG oligodeoxynucleotide 7909. J. Clin. Investig. 115:739-746. Crossref |
||||
|
Plautz GE, Yang ZY, Wu BY (1993). Immunotherapy of malignancy by in vivo gene transfer into tumors. Proc. Natl. Acad. Sci. 90:4645-4649. Crossref |
||||
| Plotkin SL, Plotkin SA (2008). A short history of vaccination. In: Plotkin SA, Orenstein WA, Offit PA (eds.), Vaccines 5th ed. New York: Elsevier. pp. 1–16. | ||||
|
Ragde H, Cavanagh WA, Tjoa BA (2004). Dendritic cell based vaccines: progress in immunotherapy studies for prostate cancer. J. Urol. 172:2532-2538. Crossref |
||||
| Restifo LL, Estes PS, Russo CD (1995). Genetics of ecdysteroid-regulated central nervous system metamorphosis in Drosophila, Diptera: Drosophilidae. Eur. J. Entomol. 92(1):169-187. | ||||
|
Rieger R, Kipps TJ (2003). CpG oligodeoxynucleotides enhance the capacity of adenovirus-mediated CD154 gene transfer to generate effective B-cell lymphoma vaccines. Cancer Res. 63:4128–4135. Pubmed |
||||
|
Robbins PF (2004). Tumor Antigen Discovery with T Cells Handbook of Cancer Vaccines. pp. 31-47. Crossref |
||||
|
Ruiz JP, Wolkowicz R,Waisman A (1998). Idiotypic immunization induces immunity to mutated p53 and tumor rejection. Nat. Med. 4:710-712. Crossref |
||||
|
Seliger B (2005). Strategies of tumor immune evasion. Biol. Drugs 19(6):347-54. Crossref |
||||
|
Small EJ, Schellhammer PF, Higano C (2006). Placebo-controlled phase III trial of immunologic therapy with Sipuleucel-T (APC8015) in patients with metastatic, asymptomatic hormone refractory prostate cancer. J. Clin. Oncol. 24:3089–3094. Crossref |
||||
|
So-Rosillo R, Small EJ (2006). Sipuleucel-T (APC8015) for prostate cancer. Expert Rev. Anticancer Ther. 6:1163–1167. Crossref |
||||
|
Stevenson FK, Rice J, Ottensmeier CH, Thirdborough SM, Zhu D (2004). DNA fusion gene vaccines against cancer. Clin. Immunol. Rev. 199:156-180. Crossref |
||||
| Tim FG, Elizabeth MJ (1999). Cancer Vaccines. J. Clin. Oncol. 17:1047-1060. | ||||
|
Timmerman JM, Czerwinski DK, Davis TA, Hsu FJ, Benike C, Hao ZM, Taidi B, Rajapaksa R, Caspar CB, Okada CY, van Beckhoven A, Liles TM, Engleman EG, Levy R (2002). Idiotype-pulsed dendritic cell vaccination for B-cell lymphoma: clinical and immune responses in patients. Blood 99:1517–1526. Crossref |
||||
|
Townsend SE, Allison JP (1993). Tumor rejection after direct costimulation of CD81 T cells by B7- transfected melanoma cells. Science 259:368-370. Crossref |
||||
| Uzzo R, Cairns P, Dulin N, Horwitz E, Pollack A, Kolenko V (2003). Tumor resistance to apoptosis: Mechanisms of evasion and implications for radiation and chemotherapeutic strategies. In Cancer Immunotherapy at the Crossroads: How Tumors Evade Immunity and What Can Be Done. The Humana Press. pp. 215-233. | ||||
|
Vergati M, Intrivici C, Huen NY, Schlom J, Tsang K (2010). Review on Strategies for Cancer Vaccine Development. J. Biomed. Biotechnol. 59: 1-13. Crossref |
||||
|
von Boehmer H (2005). Mechanisms of suppression by suppressor T cells. Nat. Immunol. 6:338-344. Crossref |
||||
|
Weng WK, Czerwinski D, Timmerman J (2004). Clinical outcome of lymphoma patients after idiotype vaccination is correlated with humoral immune response and immunoglobulinGFc receptor genotype. J. Clin. Oncol. 22:4717-4724. Crossref |
||||
| WHO (2014). World Health Organization. GLOBOCAN 2012: Estimated Cancer incidence, mortality and prevalence worldwide in 2012. IARC, 2014 Cancer Surveillance. | ||||
|
Wiemann B, Starnes CO (1994). Coley's toxins, tumor necrosis factor and cancer research: a historical perspective. Pharmacol. Ther. 64:529–564. Crossref |
||||
|
Worboys M (2007). Vaccines: conquering untreatable diseases. B.M.J. 334:1-19. Crossref |
||||
|
Wu TC, Guarnieri FG, Staveley-O'Carroll KF (1995). Engineering an intracellular pathway for major histocompatibility complex class II presentation of antigens. Proc. Natl. Acad. Sci. 92:671-675. Crossref |
||||
|
Yang D, Chertov O, Bykovskaia SN, Chen Q, Buffo MJ, Shogan J, Anderson M, Schröder JM, Wang JM, Howard OM, Oppenheim JJ (1999). Beta-defensins: linking innate and adaptive immunity through dendritic and T cell CCR6. Science 286(5439):525–528. Crossref |
||||
|
Yokokawa J, Bera TK, Palena C (2007). Identification of cytotoxic T-lymphocyte epitope(s) and its agonist epitope(s) of a novel target for vaccine therapy (PAGE4). Int. J. Cancer 121:595–605. Crossref |
||||
|
Zepp F (2010). Principles of vaccine design-Lessons from nature. Vaccine 28 Suppl 3:C14-24. Crossref |
||||
|
Zitvogel L, Casares N, Péquignot M, Chaput N, Albert ML, Kroemer G (2004). The immune response against dying tumor cells. Adv. Immunol. 84:131–179. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


