Full Length Research Paper

ABSTRACT
INTRODUCTION
Scutellonema bradys (Andrassy, 1958) is the economically most important nematodes affecting yam in West Africa (Bridge et al., 2005; Coyne et al., 2005). It is a migratory endo-parasite of roots and tubers, confined usually to the outer 1 to 2 cm of the tuber, feeding intracellular in yam tuber tissues. This results in ruptured cell walls, loss of cell contents, the formation of cavities (Castognone-Sereno and Kermarrec, 1988), tuber dry rot (Bridge et al., 2005), tuber decay and higher rates of desiccation (Nwauzor and Fawole, 1981). Pesticides can be used for nematode control, but they are expensive, unavailable or highly toxic for both the user and the environment, constituting serious health hazards (FAO, 2014; WHO, 2015). Other nematode management practices, such as hot water treatment of tubers (Speijer, 1996; Coyne et al., 2007), use of cover crops (Claudius-Cole et al., 2001; Claudius-Cole, 2005), trap crops (El-Nagdi and Youssef, 2004), chemical fertilizers (Baimey, 2005) or organic fertilizers (Adesiyan and Adeniji, 1976; Mcsorley, 2011) have been explored for yam. Recent progress in biotechnology has also shown that tissue culture of yam will lead to the disease and pest-free planting material, using aseptic in vitro meristem/shoot tip culture techniques (Ng, 1994).
However, the in vitro plants obtained are delicate and fragile, and not only free of pathogens, but also free of all natural beneficial microorganisms, such as arbuscular mycorrhizal fungi (AMF). AMFs are important elements of the soil microflora in agroecosystems, which form a mutualistic symbiosis with most plant species, including almost all plants currently micropropagated (Smith and Read, 2008). AMFs are active in increasing the availability and uptake of soil phosphorus and trace elements, thereby enhancing host plant growth (Ceballos et al., 2013; Cardio et al., 2015), can alleviate biotic and abiotic stresses (Alarcon et al., 2007; Baslam and Goicoechea, 2012; Boyer et al., 2015). Root colonization by AMF, in general, favors plant development by increasing nutrient uptake, hormonal activity, growth rate and consequently yield (Arriagada et al., 2007; Hart et al., 2014), but is also associated with pathogen suppression (Hol and Cook, 2005; Bisadu et al., 2012). Some AMF species have recently become available as commercial products for promoting crop productivity (BIORIZE Company©).
In tuber crops such as potato (Solanum spp.) and sweet potato (Ipomea batata), results following the application of commercial AMF products have shown that individual species of AMF formulation, and even combinations of a number of AMF species in a single formulation, differ in their ability to promote plant growth, which depends on the specific compatibility between plant and fungal species (Duffy et al., 1999; Deliopoulos et al., 2007). The existence of the host plants increases the need for efficient screening of AMF for host-plant species compatibility and especially for broad-spectrum associations. On yam, few studies showed the efficiency of indigenous AMF on yam growth (Lu et al., 2015). However, no studies have yet been reported on the efficiency of AMF isolates or species for promoting vitroplants growth and yield or nematodes management. The present study aimed at evaluating the effect of two commercial AMF products, based separately on F. mosseae and G. dussii, on yam growth parameters and on plant parasitic nematodes using in vitro cultures of four yam cultivars.
MATERIALS AND METHODS
Study site
The experiment was conducted at the International Institute of Tropical Agriculture (IITA), Ibadan (Nigeria), in the Southern Guinea savanna zone of Nigeria (7°30’N, 3°05’E). The experiments were established under controlled conditions in the greenhouse. The mean daily temperatures ranging between 25°C and 32°C and humidity between 65% and 85%. The night temperature ranged between 18°C and 21°C and the humidty 80% and 85%. The soil used for both experiments was collected from a depth of 0 – 15 cm at IITA Ibadan. Soil was passed through a 1 mm sieve to remove roots, sterilized by autoclaving in the oven at 80°C for 3 days and then air-dried. The soil was characterized as a sandy loam soil with a pH 6.0 and total nitrogen and available phosphorus of 0.7 g N kg-1 and 2.96 mg P kg-1, respectively (Oyekanmi et al., 2006).
Source and acclimatization of yam plantlets
For this experiment, in vitro tissue culture plantlets of D. alata (TDa 98-165, TDa 98-01183) and D. rotundata (TDr 745, TDr 87-00551), selected due to their availability, were supplied by the Biotechnology unit of IITA-Ibadan (Nigeria).
AMF inoculum and inoculation procedure
Funneliformis mosseae and G. dussii were obtained from BIORIZE Company© (Dijon, France). These commercial inocula consisted of substrates, spores, hyphae and chopped infested fine roots, 2 g of which (representing approximately 300 spores) were inoculated to each plantlet at transplanting. A hole c. 8 cm deep was made in the substrate at the middle of each 2-l pot using a pencil. The inoculum from 20 g of both AMF products.
Scutellonema bradys inoculum and inoculation procedure
S. bradys were collected from infected peels obtained from heavily infected tubers of D. rotundata cv TDr 131 from IITA-Ibadan. To determine the nematode density for the required inoculum of S. bradys, the infected tubers were manually peeled using a kitchen peeler, and cut into c. 0.5 cm x 0.3 cm pieces (Baimey et al., 2005). Nematodes were extracted from the peels for 48 h using a modified Baermann pie pan method (Coyne et al., 2007). Prior to inoculation, the density of the S. bradys suspension, which had been adjusted to 100 ml with tap water, was estimated from 3 x 10 ml aliquots, after manually shaking without allowing the nematodes to settle down. For nematodes counting, a Leica Wild M3C stereomicroscope was used. Two months after planting, each plant was inoculated with 8 ml suspension of 500 vermiform S. bradys. A water control of 8 ml was added to the control plants. Plants were subsequently watered with 300 ml tap water per plant from the base.
Approach
The experimental treatments were F. mosseae alone, G. dussi alone, F. mosseae + nematode, G. dussii + nematode and overall control (without any inoculum). The nematode inoculation were done at two levels (0 and 500). Sixty pots per yam cultivar were used, totalling 240 pots. The pots were randomly arranged. Plants received no fertilizer during the experiment and were watered regularly as required.
Assessment of AMF spore density and root colonization
Soil core samples were removed one day before harvest of yam tubers, according to Oehl et al. (2003). Roots were extracted by wet sieving and decantation, while the AMF spores were isolated by wet sieving and sucrose density gradient centrifugation (Oehl et al., 2004). The root colonization by AMF was determined according to Brundrett et al. (1996). The gridline-intersect technique (Giovannetti and Mosse, 1980) was used to analyse AMF colonization under a dissecting stereo microscope (Leica Wild M3C) at up to 90x magnification.
Assessment of yam growth parameters at harvest
The plants were harvested six months after transplanting into individual pots. The shoots were cut to soil level while tubers and roots were removed by hand, and soil gently removed away from them. The roots were removed with forceps and collected separately. Shoots, roots and tubers from each pot were rinsed gently under tap water, air dried and separately stored in labelled paper bags. Dry weight of shoots and roots were recorded following oven-drying in a well-ventilated Gallenkamp oven at 80°C for 72 h. Only fresh tuber weight was recorded as they were used for planting in the subsequent season at field sites. Total dry root weights were calculated after taking into account material removed to determine mycorrhizal colonization and nematode density.
Assessment of nematode density and tuber damage symptoms
All tubers harvested were scored for dry rot severity. The tuber dry rot severity was assessed on a scale of 1-5 (Claudius-Cole et al., 2005). All tubers per pot were scored and mean scores calculated per pot when more than one tuber per pot was present.
To assess nematode population density in roots, the complete root system of a plant was washed free of soil before chopping, mixing and removing 2 g subsamples for extraction using a modified Baermann method over a 48 h period (Hooper, 1986). S. bradys densities were assessed for all sampled tubers from 5 g sub-samples of tuber peel. Using a kitchen vegetable peeler, several ‘strips’ of peel (outer cortex) 2 cm wide and 2 to 3 mm deep were removed from different portions of the tuber, chopped finely with a knife and 5 g sub-samples per tuber removed. Nematodes were extracted from each sub-sample using a modified Baermann method over a 48 h period (Hooper, 1986).
Nematodes were also extracted from the soil of each pot by mixing all the contents of each pot, removing 3 x 50 g sub-samples and extracting them using a modified Baermann Pie Pan method (Coyne et al., 2007). Nematodes were counted with a stereomicroscope at 400x magnification.
Data analyses
All data were analysed using STATGRAPHICS, version 9.1 in Windows 2010. Three-way ANOVA was used to compare yam growth parameters (shoot, root and tuber weight) between treatments. Data on nematodes and on mycorrhization were analysed by one-way ANOVA. Prior to analysis, AMF spore density and nematode population density data were log10 (x+1) transformed, while data on mycorrhizal colonization were arcsin (x/100) transformed for homogeneous variances. The differences among treatment means were compared with Fischer’s Least Significant Difference (LSD) Test. Pearson’s correlation coefficient were used to assess the association between root colonization and various growth parameters.
RESULTS
Effect of AMF and S. bradys inoculation on yam root colonization and plant growth parameters
Mycorrhizal root colonization was moderately high (between 17% and 44%) with a significant interaction occurring between AMF inoculation and nematode inoculation (Table 1). Higher colonization by AMF was recorded from all cultivars inoculated with F. mosseae, compared to G. dussii in the treatment without S. bradys, while in the treatments with S. bradys, higher colonization by AMF was recorded from all yam cultivars except for cv. TDa98-165 inoculated with G. dussii (Table 2). Spore production was affected by both yam cultivars and S. bradys inoculation (Table 1). A higher spore density was recorded from cv TDr745 inoculated with G. dussii without S. bradys inoculation compared to cv. TDr745 plantlets inoculated with G. dussii and S. bradys (Table 2).
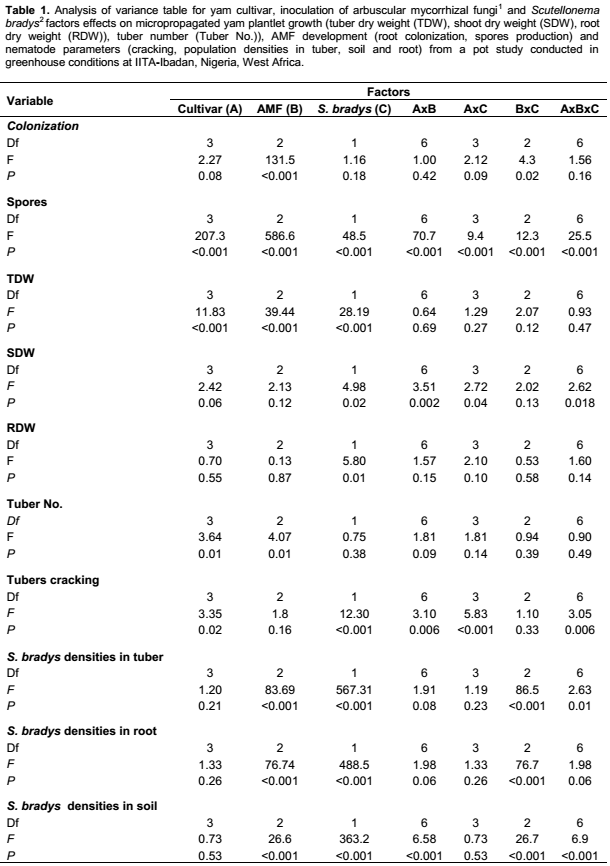
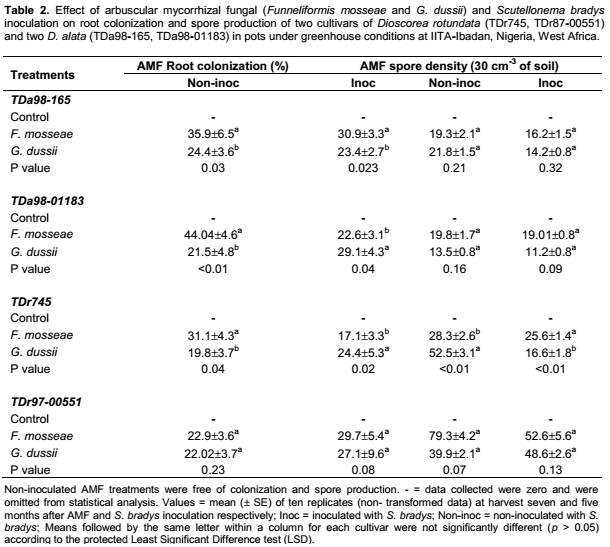
Irrespective of S. bradys inoculation, dry weight and number of tubers were significantly affected by AMF inoculation (and yam cultivar (Table 1). AMF inoculation did not affect shoot or root weight. S. bradys inoculation significantly affected tuber dry weight, shoot dry weight and root dry weight (Table 1). Without S. bradys inoculation, application of each AMF species led to a significantly higher tuber weight and number of tubers than the control for TDa98-01183 (Table 3). In addition, G. dussii application induced higher shoot and root dry weights of TDr97-000551 than the controls (Table 3). With S. bradys inoculation, only plantlets from cvs TDa98-165 and TDa98-01183 inoculated with F. mosseae yielded higher (p <0.01) than controls, while a higher number of tubers was recorded from cv TDr97-00551 when plantlets were inoculated with F. mosseae compared to controls (Table 3). Tuber cracking, and rotting severity were assessed on an arbitrary scale from 1 to 5 described by Claudius-Cole et al. (2005) where 1 = clean tuber; 2 = 1-25% tuber skin showing cracking or galling or dry rot symptoms (low level of damage); 3 = 25-50% of tuber skin showing cracking or galling or dry rot symptoms (low to moderate level of damage); 4 = 51-75% tuber skin showing cracking or galling or dry rot symptoms (moderate to severe level of damage); 5 = 76-100 % tuber skin showing cracking or galling or dry rot symptoms (high level of damage).1 AMF inoculated at rate of 300 spores per pot at yam plantlets transplanting into individual pot.2 S. bradys inoculated at rate of 500 vermiforms two months after AMF inoculation.
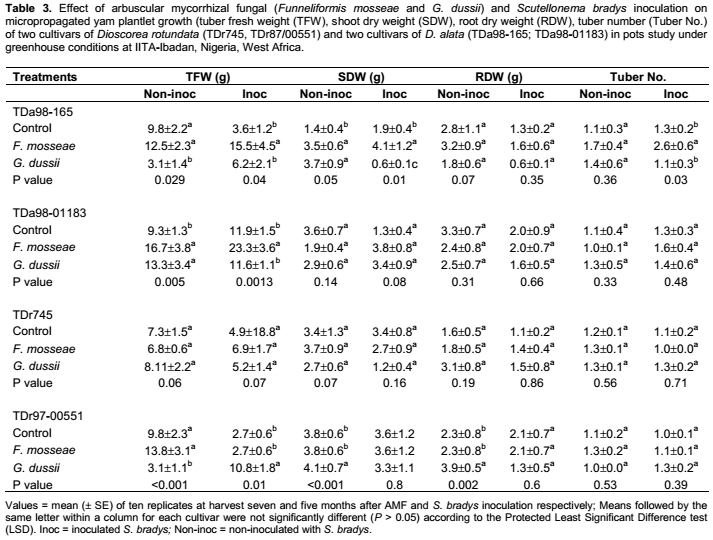
Across all cultivars, a negative correlation was observed between root colonization and tuber fresh weight (p = 0.0002) (Table 4). Within each cultivar, a negative correlation was observed between root colonization and tuber fresh weight for three of the four assessed cultivars: TDa98-165, TDa98-01183 and TDr745 (p = 0.0038, 0.0142 and 0.0147 respectively) (Table 4).
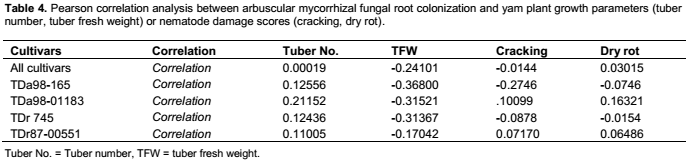
Effect of AMF inoculation on yam tuber quality and S. bradys density
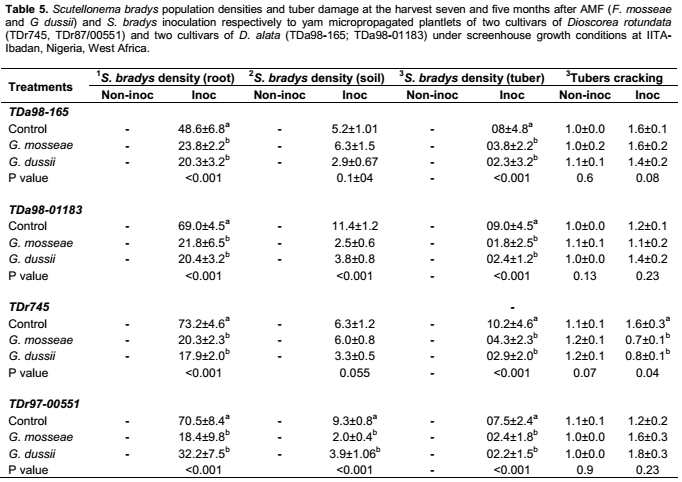
The AMF inoculation apparently did not influence the severity of S. bradys damage recorded as cracking (Table 1). Across the treatments, symptoms were very low in general (Table 5). Tuber cracking was significantly lower though, where AMF species were applied compared with controls for cvs TDa98-165 and TDr745, while tuber dry rot was significantly lower for cv TDr745 (Table 5). Interestingly, AMF inoculation significantly suppressed S. bradys densities in roots soil and in tubers (p<0.05) on plantlets where both AMF + S. bradys were inoculated, compared to S. bradys inoculation only .Tubers, roots and soil from non-inoculated nematode treatments were free of nematode where - = no data were collected. Analysis and means separation of nematode densities were undertaken on log10(x+1) transformed data; 1- Nematode density per 5-g of root; 2- Nematode per 50 g soil. 3- Nematode per 5 g yam peels. Values were mean (±SE) (non- transformed data) of ten replicates. For each yam cultivar treatments means were compared by columns and means followed by different letters were significant difference (P < 0.05) according to the protected least significant different test (LSD). Inoc = inoculated S. bradys and non-inoc = non-inoculated with S. bradys.
DISCUSSION
To date no data exist on the interaction and protective potential of AMF against S. bradys on yam. In vitro plantlets were used as an initial starting point to assess the potential of AMF on yam, with and without challenge from nematodes. The general observation from our results is that, without nematode inoculation, AMF inoculation significantly increased number and weight of tubers, but that the degree of effectiveness depends on yam cultivar and on AMF species. The latter confirms a previous result carried out with 13 AMF species and 41 AMF isolates (Tchabi et al., 2010). AMF colonization was also reported to increase yam yields in Nigeria (Oyetunji and Afolayan, 2007). Tuber Plant growth promotion in relation to AMF colonization is a well-established phenomenon across crops and climatic zones (Chaurasia and Khare, 2005; Caglar and Akgun, 2006; Antunes et al., 2011; Ceballos et al., 2013). In the current study, perhaps the most interesting result was that yam cultivars from D. alata species responded more efficiently to AMF inoculation compared to D. rotundata cultivars. One possible explanation could be related to the morphology and physiology of the two yam species. D. alata cultivars have larger leaves, intercepting more light for photosynthesis than D. rotundata species (Orkwor and Ekanayake, 1998), and probably transfer more carbohydrate to AMF, which in return uptake and transfer nutrients to the plant leading to greater tuber production. Furthermore, a possible difference in the change of phytohormone balance following the association between some yam cultivars and AMF may explain differences in yam cultivar response to AMF inoculation (Allen et al., 1980). Such differences in response to AMF inoculation among plant cultivars are reported (Johnson et al., 1997; Klironomos, 2003), indicating the variable response in root colonization, in phosphate and nitrogen uptake and fungal phosphate metabolism (Smith al., 2003; Mensah et al., 2015).
In contrast to the high colonisation, there was negative correlation between root colonisation and tuber weight. This result may be attributed to AMF functionality specificity. It was proposed by Johnson et al. (1997) that mycorrhizal association could be considered as symbioses, but the functional range along a continuum of parasitism to mutualism may vary according to environmental conditions (climate, temperature, abundance of soil nutrients, presence or absence of pathogens, etc.), the host plants genotype (Klironomos, 2003) and the differences in phosphate and nitrogen uptake and fungal phosphate metabolism (Mensah et al., 2015). The negative correlation should probably indicated that both commercial AMF species were not compatible to promote the four yam cultivars growth used in the present experiment (Klironomos, 2003; Lu et al. 2015) thus, selecting effective AMF species is necessary to facilitate yam growth and improve quantity of yam tubers and the protective effect against yam nematode. Yam in vitro plantlets inoculated with F. mosseae, followed by S. bradys, produced heavier tubers compared to S. bradys alone for TDa98-165, TDa98-01183 and TDr97-00551. Interestingly, a stimulatory effect on tuber weight was observed for plantlets inoculated with both F. mosseae and S. bradys compared to single inoculation of F. mosseae or G. dussii using cv TDa98-01183. These results indicate that AMF can lead to suppression of nematode damage through the phenomenon of compensation (Smith and Read, 2008). Other authors undertaken similar observations and established that a low nematode population in dual combination with endophytic microbes could stimulate host plant growth and yield (Brown and Kerry, 1987; Hao et al., 2005; Zum-Felde et al., 2006). Also, considering that tuber formation in yam is hormonally mediated (Okwor and Ekanayake, 1998), it may be hypothesized that F. mosseae, in combination with S. bradys challenge, affected the hormone balance in yam plantlets (e.g. increased synthesis of growth regulators in response to nematode infection), leading to increased production of yam tubers. McKenry et al. (2001) reported that grapevine (Vitis spp.) (cvs VR 039-16, Schwarzmann, and Freedom rootstocks) had grown larger in the presence of Xiphinema americanum than in its absence.
The present study showed a reduction of nematode density. The mechanism by which AMF reduces nematode damage has not been fully determined. Hypotheses range from depression of nematode development by competition for nutrient and space (Smith et al., 1986; Elsen et al., 2008), microbial changes in the mycorrhizosphere that disturb nematode chemotaxis (Linderman, 1988; Brussaard et al., 2007) to induced resistance through a pre-activation of gene and corresponding proteins responsible for plant defence against pathogen attacks (Slezack et al., 2000; Deliopoulos and Haydock, 2003). For an efficient option for using AMF to protect against nematode damage, AMF essentially need to be established in the roots before nematode attack in order to provide biological control review (Borowicz, 2001; Diedhiou et al., 2003). For this reason, the ‘impregnation’ of AMF at weaning of in vitro plantlets would appear suitable and appropriate, before planting out in the field, where nematode (and other pest and disease) challenge would occur. Tubers were also assessed for symptoms of dry rot and cracking. The fact that the tubers cracking and dry rot symptoms in mycorrhizal yam were not significant different compared to non-mycorrhizal plants indicates probably that the damage of S. bradys occurs less during harvest period (Meerman et al., 2000) but occurs mostly during storage period (Bridge, 1973). The reason for this is that the multiplication of S. bradys in yam tubers is greatest and most severe, during storage (Cadet and Queneherve´, 1994). Damage to yield may occur, however, damage and losses correlate with duration of the tuber storage period (Bridge et al., 2005).
CONCLUSION
Our results have shown for the first time that a commercial AMF can stimulate yam growth parameters. However, AMF species influence on plant growth appears highly dependent on the plant genotype with which they are associated. Taking into consideration tuber weight, number of tubers, AMF attributes, and nematode management, F. mosseae, was a more effective AMF symbiont for association with yam plantlets under the conditions of the study (in pots on In vitro plant) than G. dussii originating from West Africa.
Further studies are needed to verify this hypothesis, which is also likely to vary with cropping environment, crop and cultivar. Alternatively, inoculation with two fungal taxa in dual combination might improve the overall synergistic interaction between plants and fungi and may reflect the possible different roles of AMF within a fungal community (van der Heijden and Kuyper, 2001), towards resolving the selection problem for the most specific fungal partner.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This work was supported by grants from the Swiss Center for International Agriculture (ZIL: http://www.rfpp.ethz.ch), the Indo-Swiss Collaboration in Biotechnology (ISCB: http://iscb. epfl.ch/), and the Swiss National Science Foundation. The publication fees was supported by UEMOA (Union Economique et Monétaire Ouest Africaine).
REFERENCES
|
Alarcon A, Davies Fred T, Davies Jr, Robin L, Davis A, Zuberer DA (2008). Arbuscular mycorrhiza and Petroleum-Degrading microorganisms enhance phytoremediation of petroleum-contaminated soil. Int. J. phytoremediation 10(4): 251-263. |
|
|
Allen MF, Moore Jr. TS, Christensen M (1980). Phytohormones changes in Bouteloua gracilis infected by vascular arbuscular mycorrhizae. I. Cytokinin increases in the host plant. Can. J. Bot. 58: 371-374. |
|
|
Antunes PM, Koch AM, Morton JB, Rillig MC, Klironomos JN (2011). Evidence for functional divergence in arbuscular mycorrhizal fungi from contrasting climatic origins. New Phytol. 189:507-514. |
|
|
Arriagada CA, Herrera, MA, Borie F, Ocampo JA (2007). Contribution of arbuscular mycorrhizal and saprobe fungi to the aluminum resistance of Eucalyptus globulus. Water Air Soil Pollut. 182(1-4): 383-394. |
|
|
Baimey H, Coyne D, Labuschagne N (2005). Effect of fertiliser application on yam nematodes (Scutellonema bradys) multiplication and consequent damage to yam (Dioscorea spp.) under field and storage conditions in Benin. Int. J. Pest Manag. 52: 63-70. |
|
|
Baslam M, Goicoechea N (2012). Water deficit improved the capacity of arbuscular mycorrhizal fungi (AMF) for inducing the accumulation of antioxidant compounds in lettuce leaves. Mycorrhiza 22:347-359. |
|
|
Borowicz VA (2001). Do arbuscular mycorrhizal fungi alter plant-pathogen relations. Ecology 82: 3057-3068. |
|
|
Boyer LR, Brain P, Xu X-M, Jeffries P (2015). Inoculation of drought-stressed strawberry with a mixed inoculum of two arbuscular mycorrhizal fungi: Effects on population dynamics of fungal species in roots and consequential plant tolerance to water deficiency. Mycorrhiza 25: 215-227. |
|
|
Bridge J, Coyne D, Kwoseh CK (2005). Nematode parasites on Root and Tuber Crop. In: Luc M, Sikora RA and Bridge J (eds) Plant Parasitic Nematodes in Subtropical and Tropical Agriculture. 2nd Ed. CAB Int. Wallingford, UK. pp. 221-258. |
|
|
Brown RH, Kerry RH (1987). Principle and practice of nematode control in crops. Academic Press Australia, Australia 447pp. |
|
|
Brundrett M, Bougher N, Dell B, Grove T, Malajczuk N (1996). Working with Mycorrhizas in Forestry and Agriculture. ACIAR Monograph 32. Australian Centre for International Agricultural Research, Canberra. |
|
|
Cadet P, Queneherve´ P (1994). Fluctuations naturelles de Scutellonema bradys (Nematoda: Hoplolaimidae) au cours de la croissance et du stockage de l'igname (Dioscorea alata) à la Martinique.Nematologica 40:587-600. |
|
|
Sereno P, Kermarrec A (1988). Association between Pratylenchus coffeae and Scutellonema bradys in yam tubers under agronomic conditions in the French West Indies. Nematropica 18: 155-157. |
|
|
Ceballos I, Ruiz M, Fernández C, Pe-a R, Rodríguez A, Sanders IR (2013). The in vitro mass-produced model mycorrhizal fungus, Rhizophagus irregularis, significantly increases yields of the globally important food security crop cassava. PLoS ONE 8(8): e70633. |
|
|
Chaurasia B, Khare PK (2005). Hordeum vulgare: a suitable host for mass production of arbuscular mycorrhizal fungi from natural soil. Appl. Ecol. Environ. Res. 4: 45-53. |
|
|
Claudius-Cole AO (2005). Cover crops in the management of Meloidogyne spp. and Scutellonema bradys on edible yam, studies in Nigeria. Ph.D Thesis Faculty of Agriculture University of Ibadan, Ibadan, Nigeria. |
|
|
Claudius-Cole AO, Asiedu R, Fawole B (2001). Cultural control of Scutellonema bradys on Dioscorea spp. In: Akporoda MO, Ekanayake IJ (eds). Proceedings of the International Symposium for Root and Tuber Crops, 12-16 November 2001, Ibadan pp. 522-526. |
|
|
Coyne DL, Tchabi A, Baimey H, Labuschagne N, Rotifa I (2006). Distribution and prevalence of nematodes (Scutellonema bradys and Meloidogyne spp. on marketed yam (Dioscorea spp.) in West Africa. Field Crops Res. 96: 142-150. |
|
|
Coyne D, Nicol J, Claudius-Cole A (2007). Practical Plant Nematology: Field and Laboratoty Guide. IITA, Ibadan, Nigeria. 82pp. |
|
|
Deliopoulos T, Devine KJ, Haydock PJ, Peter WJ (2007). Studies on the effect of mycorrhization of potato roots on the hatching activity of potato root leachate towards the potato cyst nematodes, Globodera pallida and G. rostochiensis. Nematology 9: 719-729. |
|
|
Diedhiou PM, Hallmann J, Oerke EC, Dehne HW (2003). Effects of arbuscular mycorrhizal fungi and a non-pathogenic Fusarium oxysporum on Meloidogyne incognita infestation of tomato. Mycorrhiza 13: 199-204. |
|
|
Duffy EM, Hurley EM, Cassells AC (1999). Weaning performance of potato microplants following bacterization and mycorrhization. Potato Res. 42(3-4): 521-527 |
|
|
El-Nagdi WMA, Youssef MMA (2004). Soaking faba bean seed in some bio-agents as prophylactic treatment for controlling Meloidogyne incognita root-knot nematode infection. J. Pest. Sci. 77: 75-78. |
|
|
Elsen A, Gervacio D, Swennen R, De Waele D (2008). AMF-induced biocontrol against plant parasitic nematodes in Musa sp.: A systemic effect. Mycorrhiza: 18: 251-256. |
|
|
FAO (Food and Agriculture Organization of the United Nation) (2014). FAO Statistical Yearbook: Africa Food and Agriculture. Regional Office for Africa, Accra, 2014. |
|
|
Giovannetti M, Mosse B (1980). An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 84: 489-500. |
|
|
Hao Z, Christie P, Qin L, Wang C, Li X (2005). Control of Fusarium Wilt of Cucumber Seedlings by Inoculation with an Arbuscular Mycorrhizal Fungus. J. Plant Nutr. 28: 1961-1974. |
|
|
Hart M, Ehret DL, Krumbein A, Leung C, Murch S, Turi C, Franken P (2014). Inoculation with arbuscular mycorrhizal fungi improves the nutritional value of tomatoes. Mycorrhiza 25: 359-376. |
|
|
Hol WHG, Cook R (2005). An overview of arbuscular mycorrhizal fungi–nematode interactions. Basic Appl. Ecol. 6: 489-503. |
|
|
Johnson NC, Graham JH, Smith A (1997). Functioning of mycorrhizal association along the mutualism-parasitism continuum. New Phytol. 135: 575-586. |
|
|
Klironomos JN (2003). Variation in plant response to native and exotic arbuscular mycorrhizal fungi. Ecology 84: 2292-2301. |
|
|
Lu F, Lee C, Wang C (2015). The influence of arbuscular mycorrhizal fungi inoculation on yam (Dioscorea spp.) tuber weights and secondary metabolite content. Peer J 3: e1266. |
|
|
McKenry MV, Kretsch JO, Anwar SA (2001). Interactions of selected rootstocks with ectoparasitic nematodes. Am. J. Ecol. Viticult. 52:304-309. |
|
|
Mcsorley R (2011): Overview of Organic Amendments for Management of Plant-Parasitic Nematodes, with Case Studies from Florida. J. Nemat. 43:69-81. |
|
|
Meerman JC, Speijer, PR, Vernier P, Asiedu R (2000). Establishment of the geographic distributiuon of yam nematode pests in the Republic of Benin. In: 1999 Annual Report: Plant Health Management Division. International Institute of Tropical Agriculture (IITA), 125pp. |
|
|
Mensah JA, Koch MA, Antunes PM, Kiers ET, Hart M, Bücking H (2015). High functional diversity within species of arbuscular mycorrhizal fungi is associated with differences in phosphate and nitrogen uptake and fungal phosphate metabolism. Mycorrhiza 25:533-546 |
|
|
Ng SYC (1994). Production and distribution of virus-free yam (Dioscorea rotundata Poir). In: Ofori I F, Hahn S K (eds) Proceedings of the 9th Symposium of the International Society of Tropical Root Crops (20-26 October 1991) Accra, Ghana. pp. 324-328. |
|
|
Oehl F, Sieverding E, Ineichen K, Mäder P, Boller T, Wiemken A (2003). Impact of land use intensity on the species diversity of arbuscular mycorrhizal fungi in agroecosystems of Central Europe. Appl. Environ. Microbiol. 69:2816-2824. |
|
|
Oehl F, Sieverding E, Mäder P, Dubois D, Ineichen K, Boller T, Wiemken A (2004). Impact of long-term conventional and organic farming on the diversity of arbuscular mycorrhizal fungi. Oecologia 138:574-583. |
|
|
Orkwor GC, Ekanayake IJ (1998). Growth and development of yam. In: Orkwor GC, Asiedu R and Ekanayake IJ (eds) Food Yams: International Institute of Tropical Agriculture. Adv. Res. pp. 39-62. |
|
|
Oyekanmi EO, Coyne DL, Fagade OE, Osonubi O (2007). Improving root-knot nematode managment on two soybean genotypes through the application of Bradyrhizobium japonicum, Trichoderma pseudokoningii and Glomus mosseae in full factorial combinations. Crop Prot. 26:1006-1012. |
|
|
Oyetunji OJ, Afolayan ET (2007). The relationship between relative water content, chlorophyll synthesis and yield performance of yam (Dioscorea rotundata) as affected by soil amendments and mycorrhizal inoculation. Arch. Agron. Soil Sci. 53:335-344. |
|
|
Smith SE, Read DJ (2008). Mycorrhizal Symbiosis, 3rd edn. Academic Press, London, UK. |
|
|
Speijer PR (1996). Hot water treatment: a possible strategy for farmer group and NGOs. In: Green KR and Florini DA (eds) Pests and pathogens of yams in storage: A workshop report. Afr. J. Root Tuber Crops 1: 38-42. |
|
|
StatPoint Inc. (2007). The Multilingual Statgraphics Centurion User's Guide. Statistics. |
|
|
Tchabi A, Coyne D, Hountondji F, Lawouin L, Wiemken A, Oehl F (2010). Efficacy of indigenous arbuscular mycorrhizal fungi for promoting white yam (Dioscorea rotundata) growth in West Africa. Appl. Soil Ecol. 45: 92-100. |
|
|
WHO - World health Organization (2014). |
|
|
Zum-Felde A, Pocasangre CA, Monteros C, Sikora RA, Rosales Feriveros RAS (2006). Effect of combined inoculations of endophytic fungi on the biocontrol of Radopholus similis. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


