ABSTRACT
The use of biochar has been shown to suppress populations of soil-borne pathogens. Hence it has been promoted as one of the eco-friendly alternatives to synthetic pesticides. In the current study, the effects of different sources of indigenous biochar on Meloidogyne incognita and overall growth and yield of tomato were evaluated under screenhouse condition in 2016 and 2017. A total of 10 treatments were evaluated in the study including tomato plants infected with 5000 infective juveniles of M. incognita treated with biochar woodchips of gum arabic (Acacia nilotica), bush mango (Irvingia gabonensis), neem (Azadirachta indica), goat head (Acanthospermun hispidum), gmelina (Gmelina arborea), locust bean (Parkia biglobosa), lagos mahogany (Khaya ivorensis), fig (Ficus sur), eucalyptus (Eucalyptus camaldulensis) and an untreated control. Treatments were laid-out in completely randomized design (CRD) and replicated four times. Data collected on nematode damage indices, plant height, number of leaves, and fruit yield/plant were subjected to analysis of variance. Means were separated using Duncan’s new multiple range test at 5% level of probability. Among all the biochar treatments evaluated, bush mango (BM) and neem (N) biochars significantly (p≤0.05) reduced root galling index (RGI), infective juveniles and final populations of M. incognita. In 2016 and 2017, the use of BM and N resulted in 64.05 and 66.52% reduction in galling, respectively while neem biochar reduced RGI by 66.49 and 70.76%, respectively and increased yield of tomato by 57.5%. Findings from this study have provided evidence that biochar formulated from indigenous woodchips in Nigeria, especially bush mango (Irvingia gabonensis) and neem (A. indica) are a promising tool in the control of M. incognita.
Key words: Biochar, tomato, Meloidogyne incognita, indigenous tree, Nigeria.
Tomato (Solanum lycopersicum L.) fruit constitutes one of the major condiments of diets used in most cuisines in Nigeria (Babalola et al., 2010). The fruits are popular and frequently used for a number of delicious dishes such as stew, salad and sauce and are also economical, shelf stable, and easy to use (Oyinlola and Jinadu, 2012). It has been reported that consumption of tomato improves human health due to the high vitamin content of its fruit
(SCANNERS, 2009). The cultivation of tomato also serves as a source of income to farmers and other actors along the tomato value chain. In a study, Zalkuwi et al. (2012) showed that tomato production is profitable in both short and medium term owing to the positive gross margin and net income per hectare of N90,113.78 (≈ $237) and N68,296.26 (≈ $180), respectively. Despite the promising benefits of the crop, its production has been reported to be limited by plant-parasitic nematodes, fungal and bacterial diseases, hence keeping production at sub-optimal levels (ŽnidarÄiÄ et al., 2003; Olaoye et al., 2011; Etebu et al., 2013).
Plant-parasitic nematodes represent a formidable pest problem in the field, greenhouses and polyhouses (Luc et al., 2002). In Ghana, Osei et al. (2011) observed larger populations of plant-parasitic nematodes on tomatoes than on Mucuna pruriens L. and Tithonia diversifolia Hemsl. Et Gray. Losses in the range 20-94% due to nematode infections were recorded in Nigeria (Olowe, 1978). Osei et al. (2011) reported that eight genera of plant-parasitic nematodes are commonly associated with tomato. These nematode genera included Meloidogyne, Helicotylenchus, Hoplolaimus (Sher), Pratylenchus, Rotylenchulus, Scutellonema, Tylenchulus and Xiphinema. Of all these nematodes, Meloidogyne spp. was reported to be the most damaging. Meloidogyne incognita is the most-damaging to vegetables in tropical and subtropical countries causing losses up to 80% in heavily infested fields (Adesiyan et al., 1990; Kaskavalci, 2007; Onkendi et al., 2014). Typically, M. incognita causes formation of galls on the roots. After penetrating the root elongation zone and migrating intercellularly towards the root tip, it enters the vascular cylinder, where it further punctures the cell wall with its stylet and injects secretions from the pharyngeal glands into the plant cell inducing a permanent feeding site known as giant cells (Davis et al., 2000; Gheysen and Mitchum, 2009).
There are several approaches such as use of resistant varieties (Williamson and Kumar, 2006), cultural practices (Okada and Harada, 2007), biological control (Hossain et al., 1989; Khan et al., 2007) and the use of nematicides (Hossain et al., 1989; Radwan et al., 2012). Although the use of synthetic nematicides has been mostly deployed to manage RKN populations in the soil, there is rising concern on their hazardous effects on the environment and non-target organisms and most recently, the promotion of climate change mitigating practices among farming communities including the use of biochar. The International Biochar Initiative defined biochar as “a solid material obtained from the thermo-chemical conversion of biomass in an oxygen limited environment” (IBI, 2012).
Kolton et al. (2011) described biochar as a heterogeneous material generated through pyrolysis, a thermal process carried out at temperatures ranging from 200 to 900°C and under limited oxygen availability. A wide range of organic materials including crop residues (Yuan and Xu, 2011), wood (Spokas and Reicosky, 2009), municipal waste (Mitchell et al., 2013), sewage sludge (Méndez et al., 2012), manure (Uzoma et al., 2011) and also animal bones (Vassilev et al., 2013) have been used to formulate biochar.
Biochar has been used to control certain soil-borne pathogens (Elad et al., 2010; Elmer and Pignatello, 2011; Graber et al., 2014; Jaiswal et al., 2014). Zhang et al. (2013) reported that although the paucity of research on how biochar affects nematode community structure exist, addition to the soil significantly increased the abundance of fungivorous nematodes, and decreased plant-parasitic nematodes in China. In spite of the considerable research and application of biochar in soil amendment regimes, its possibility as a tool for effective control of plant diseases has been mainly ignored (Bonanomi et al., 2015). In fact Huang et al. (2015) reported that despite the control potential of biochar, data were limited on the effect of biochar on plant parasitism by nematodes. It is against this background that this research was conducted to evaluate the effect of some indigenous woodchip-biochars on Meloidogyne incognita infection on tomato. The study further examined the concurrent effect the application of these biochars may have on tomato growth and yield.
Description of experimental site
The experiment was conducted in a screenhouse located at the Teaching and Research Farms of Federal University of Agriculture, Makurdi (Latitude 07°45’ to 07°50’N, Longitude 08°45’E to 08°50’E, 98 m above sea level) during the cropping seasons of 2016 and 2017. The location falls within the Southern Guinea Savannah agro-ecological zone of Nigeria. It experiences a typical tropical climate with two distinct seasons, the wet or rainy season and the dry season, annual rainfall of 1500 mm and an average temperature which ranges between 21 to 35°C.
Soil sterilization and establishment of tomato nursery
Top sandy-loam soil (20 cm deep) was collected from a site at the Teaching and Research Farms of Federal University of Agriculture, Makurdi (FUAM), Benue State, Nigeria. The soil was sterilized by heating for 90 min at 65°C with the aid of a soil sterilizer according to Atungwu et al. (2008). Sterilized soil was distributed into a nursery bed (180 cm × 50 cm×10 cm). The soil in the nursery bed was preconditioned by watering adequately to field capacity and left for three weeks. Tomato cv. Roma VF was obtained from National Horticultural Research Institute (NIHORT), Ibadan, Nigeria. Prior to planting, seeds were surface sterilized by immersion in 70% ethanol for 1-2 min and 0.2% (v/v) sodium hypochlorite for 5-10 min, respectively under aseptic conditions. Twenty grams (20g) of seeds were broadcast on the nursery bed under shade and maintained for two weeks after which they were transplanted to experimental pots.
Formulation of biochar
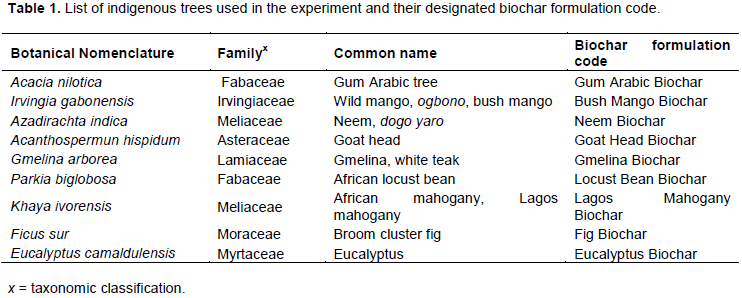
Dried woods obtained from nine mature indigenous trees of similar ages located in the metropolitan city of Makurdi in Benue State, Nigeria were used to formulate nine different biochars as shown in Table 1 based on the principle of pyrolysis using a 208 L traditional kiln (reactor) fabricated at FUAM. A total of 2.5 kg dried wood of Acacia nilotica, Irvingia gabonensis, Azadirachta indica, Acanthospermun hispidum, Gmelina arborea, Parkia biglobosa, Khaya ivorensis, Ficus sur and Eucalyptus camaldulensis were burnt under low oxygen (pyrolysis) using the fabricated reactor. The kiln was properly cleaned using distilled water after each pyrolysis process, and allowed to dry. After cooling, prepared biochars were ground into powder of 0.1 to 1.0 mm particles (Elad et al., 2010). The biochar was mixed with heat-sterilized soil at 30 g/kg soil into individually assigned experimental pots.
Inoculation of tomato plants with M. incognita eggs
Motile second-stage infective juveniles of Meloidogyne incognita were extracted from 5 g of symptomatic roots and soil samples obtained from pure cultures raised on three-month’s old galled Celosia argentea plants at the Teaching and Research Farms of FUAM using modified Baermann tray technique (Whitehead and Hemming, 1965). The nematode suspension was thoroughly mixed using a magnetic stirrer. The number of infective juveniles in 10 ml of the suspension was counted in a Doncaster counting dish under the dissecting microscope. An average of three counts was taken to estimate the infective juvenile population per ml of nematode suspension. Infective nematode population density was adjusted to 5000 juveniles by inoculating tomato plants with 100 ml of concentrated nematode suspension. One hundred milliliter of distilled water was used in the control. Each plant stand was inoculated with 5000 infective juveniles of M. incognita one week after transplanting.
Experimental design
A total of 10 treatments were evaluated in the study and included biochar from gum arabic, bush mango, neem, goat head, gmelina, locust bean, lagos mahogany, fig, eucalyptus and an untreated control. Treatments were laid-out in a Completely Randomized Design (CRD) and replicated four times. Weeding was done on a weekly basis and NPK (15:15:15) was applied at the rate of 120 kg/ha 2WAT (4.3 g/pot).
Nematode extraction and identification
A total of 250 g of soil rhizosphere samples from each pot was collected using a soil auger, to a depth of 20 cm. Soil samples collected from same treatment replicates were thoroughly mixed and bulked into 250g sample, properly labelled and taken to the laboratory for nematode extraction. Root of each plant/treatment was also labelled and assayed for M. incognita. Nematodes were extracted from the bulked soil samples using the modified Baermann tray technique (Whitehead and Hemming, 1965). Extraction of nematodes from the roots was carried out using the technique described by Hussey and Barker (1973). Nematode specimens were identified based on morphological characteristics using pictorial keys (Mai and Lyon, 1975).
Data collection and statistical analysis
Agronomic and nematode damage data were collected during the span of the experiments in 2016 and 2017. The agronomic data included plant height at 50% flowering (height of plant from the base of the stem above soil to the flag leaf before the lowest flower in centimeters), number of branches at 50% flowering (number of fresh and dry branches for each plant stand/pot), number of leaves at 50% flowering (the total number of both leaves found on each plant/pot), number of fruits (number of tomato fruit counted per pot).
Damage data included root gall index, number of infective juveniles/250 g of soil and final nematode population were used to assess nematode damage on tomato. Root gall index was assessed on a scale of 0-5, where, 0 = no gall; 1 = 1 to 2 galls; 2 = 3 to 10 galls; 3 = 11 to 30 galls; 4 = 31 to 100 galls; 5 = more than 100 galls according to Taylor and Sasser (1978). Data were subjected to Analysis of Variance (ANOVA) using general linear model (GLM) procedure of GenStat 17th Edition (Lawes Agricultural Trust, VSN International, 2014) Statistical Software Package. Means were compared using Duncan’s New Multiple Range Test at 5% level of probability. Nematode counts were square-root [(√(x+1)] transformed to improve homogeneity of variance before analysis.
Effect of biochar on M. incognita
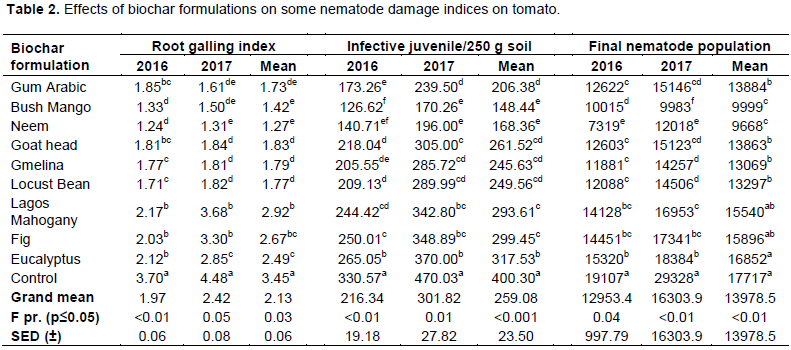
Findings obtained with respect to root galling index (RGI), number of infective juveniles/250g of soil (IJ2) and final nematode population (Pf) generally followed a similar trend as presented in Table 2. Among all biochar formulations, Bush Mango and Neem biochars significantly (p≤0.05) reduced RGI, IJ2 and Pf of M. incognita the most. In 2016 and 2017 use of bush mango biochar resulted in percentage reductions of RGI on tomato by 64.05 and 66.52%, respectively while neem biochar reduced RGI by 66.49 and 70.76%, respectively. Pots amended with goat head biochar, gmelina biochar and locust bean biochar ranked second among all the evaluated biochar formulations with average RGI values of 1.83, 1.79 and 1.77 representing percentage galling reductions of 46.96, 48.12 and 48.7%, respectively. Although use of lagos mahogany biochar, fig biochar and eucalyptus biochar significantly reduced RGI, they were the least effective since they only resulted in 15.36, 22.61 and 27.83% reduction of RGI against the untreated control.
The number of infective juveniles was significantly (p≤0.05) affected by application of biochar formulations (Table 2). Pots treated with bush mango biochar and neem biochar also consistently and significantly reduced IJ2. Average mean IJ2 values showed that of all biochar formulations evaluated, only bush mango biochar (circa. 148 J2s), Neem biochar (circa. 168 J2s), gum arabic biochar (circa. 206 J2s), gmelina biochar (circa. 246 J2s) and locust bean biochar (circa. 250 J2s) were below the grand mean value ((circa. 259 J2s) and resulted in percentage reduction in the number of infective juveniles by 42.7, 35.02, 20.34, 5.19 and 3.67%.
The population of M. incognita significantly decreased in 2016 and 2017 when pots were treated with bush mango biochar (9999 M. incognita) and neem biochar (9668 M. incognita). Although the highest population of M. incognita was encountered in the control pots, separation of means using Duncan’s New Multiple Range Test consistently showed that there were no statistically significant differences in the populations of the control pots, Lagos Mahogany biochar-treated pots, Fig biochar-treated pots and Eucalyptus biochar-treated pots (p>0.05) (Table 2).
Effect of biochar on growth and yield of tomato
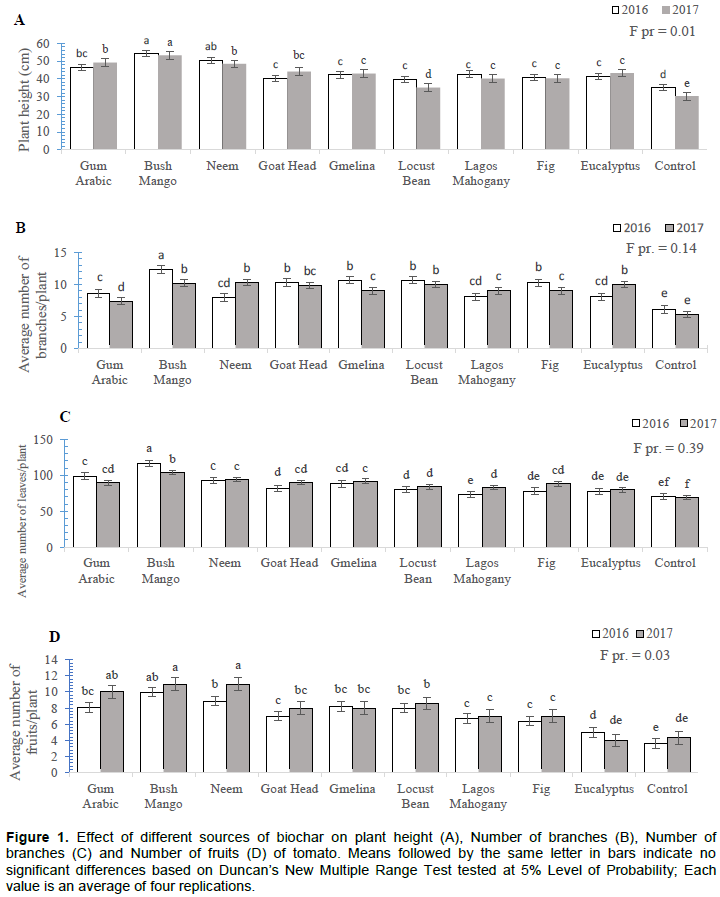
The effects of biochar formulation on tomato growth and yield were also determined and presented in Figure 1. The four parameters were significantly affected by the type of biochar applied on tomato (Figures 1A, B, C and D). Plants grown in pots amended with bush mango and neem biochars were the tallest (Figure 1A) and had the highest number of fruits (Figure 1D), closely followed by gum arabic biochar. The lowest number of fruits was observed in the control pots and was not significantly different from fruit yield recorded in Eucalyptus biochar-treated pots (p>0.05). Bush mango biochar also had the most profound effect on number of branches/plant and number of leaves/plant. The performance in terms of the four measured parameters was observed in the untreated control.
The use of nematicides has serious adverse consequences and several useful chemicals are being phased out of the market and beyond the reach of farmers, thus creating the need for alternative measures for effective management of plant-parasitic nematodes (Daramola et al., 2015). There is a worldwide drive to shift from a petroeconomy fueled by fossil carbon to an economy fueled by renewable energy resources as a result of growing concern over global climate change caused by manmade, anthropogenic greenhouse gas emissions (Mathews, 2008; Elad et al., 2010). In recent years, in order to develop a more eco-sustainable agriculture, research has been carried out on strategies for plant pathogen control characterized by high efficiency and limited environmental impact (Bonanomi et al., 2015). One of these control strategies developed to manage soil-borne pathogens is the use of biochar Elad et al., 2010; Bonanomi et al., 2015). These biochars have been reported to stimulate beneficial microbes, in bulk soil as well as in the rhizosphere (Lehmann et al., 2011; Thies et al., 2015).
The current research investigated the effects of indigenous biochar on M. incognita which constitutes a major threat to tomato production. Our findings clearly showed that the use of biochars, especially gum arabic biochar, bush mango, neem biochar, goat head biochar, gmelina biochar and locust bean biochar significantly repressed populations of the nematode by varying proportions. Biochar had also been reported to induce resistance against different pathogens. For example, tomato plants treated with biochar pyrolyzed from citrus wood suppressed grey mold and powdery mildew caused by Botrytis cinerea and Leveillula taurica (Elad et al., 2010). Biochar prepared from pepper plant waste suppressed three foliar diseases (B. cinerea, C. acutatum, P. aphanis) with different infection strategies in strawberry plants (Harel et al., 2012). Jaiwal et al. (2014) found that biochars pyrolyzed from eucalyptus wood chips and pepper plant wastes were found to be effective at decreasing the severity of R. solani infection in beans.
Eucalyptus-based biochar in the current study did not result in a significant control of the nematode. Nevertheless, there was a marked difference in the populations of M. incognita and overall tomato growth of treated pots compared to the untreated pots. These findings suggest that the effect of these biochars could be the result of a balance between the availability of labile organic carbon sources and the presence of recalcitrant and/or nematotoxic compounds that provide little support or even inhibit reproduction of M. incognita. At least five different mechanisms have been proposed in an attempt to underscore how biochars suppress plant diseases. These include: (i) induction of systemic resistance in the host plants; (ii) enhanced abundance and/or activities of beneficial microbes; (iii) modification of soil quality in terms of nutrient availability and abiotic conditions; (iv) direct toxic effect of biochar; (v) sorption of allelopathic, phytotoxic compounds (Bonanomi et al., 2015). Several studies reported an increase of microbial biomass47, mycorrhizal fungi (Warnock et al., 2007), and plant-growth-promoting microbes (Liang et al., 2010; Kolton et al., 2011) as a result of biochar applications, with related changes in microbial community functionality.
CONCLUSION AND RECOMMENDATIONS
In conclusion, findings from this study have provided evidence that biochar formulated from indigenous wood chips in Nigeria, especially bush mango (I. gabonensis) and neem (A. indica) are a promising tool in the control of M. incognita. These plants are readily available and processing of their biochar is relatively cheap and indicative of low cost alternative to synthetic nematicides. However further research is needed towards a better understanding of the precise mode of action of these two biochars under field conditions, and to develop stable and low cost nematostatic formulations that can be applied on other plant-pathogen systems as well.
The authors have not declared any conflict of interests.
REFERENCES
|
Adesiyan SO, Caveness FE, Adeniji MO, Fawole B (1990). Nematode pest of tropical crops. Heinemann Educational Books (Nig.) Plc. 114p.
|
|
|
|
Babalola DA, Makinde YO, Omonona BT, Oyekanmi MO (2010). Determinants of post-harvest losses to tomato production: A case study of Imeko-Afon local government area of Ogun State. Journal of Life and Physical Science 2:14-18.
|
|
|
|
|
Bonanomi G, Ippolito F, Scala F (2015). A "black" future for plant pathology? Biochar as a new soil amendment for controlling plant diseases. Journal of Plant Pathology 97(2):223-234.
|
|
|
|
|
Daramola F, Afolami S, Enikuomehin O, Omonhinmin C, Adebayo A (2015). Nematicidal effects of carbofuran and GC-MS analysis of its residue in pineapple fruits. International Journal of Agriculture and Biology 17:357-362.
|
|
|
|
|
Davis EL, Hussey RS, Baum TJ, Bakker J, Schots A, Rosso MN (2000). Nematode parasitism genes. Annual Review Phytopathology 38:365-396.
Crossref
|
|
|
|
|
Elad Y, David DR, Harel YM, Borenshtein M, Kalifa HB, Silber A, Graber ER (2010). Induction of systemic resistance in plants by biochar, a soil-applied carbon sequestering agent. Phytopathology 100:913-921.
Crossref
|
|
|
|
|
Elmer WH, Pignatello JJ (2011). Effect of biochar amendments on mycorrhizal associations and fusarium crown and root rot of asparagus in replant soils. Plant Disease 95:960-966.
Crossref
|
|
|
|
|
Etebu E, Nwauzoma AB, Bawo DDS (2013). Postharvest spoilage of tomato (Lycopersicon esculentum Mill.) and control strategies in Nigeria. Journal of Biology, Agriculture and Healthcare 3(10):51-61.
|
|
|
|
|
Gheysen G, Mitchum MG (2009). Molecular insights in the susceptible plant response to nematode infection. Plant Cell Monogram 15:45-81.
Crossref
|
|
|
|
|
Graber ER, Frenkel O, Jaiswal AK, Elad Y (2014). How may biochar influence severity of diseases caused by soilborne pathogens? Carbon Management 5:169-183.
Crossref
|
|
|
|
|
Harel Y, Elad Y, Rav-David D, Borenstein M, Shulchani R, Lew B (2012). Biochar mediates systemic response of strawberry to foliar fungal pathogens. Plant Soil 357:245-257.
Crossref
|
|
|
|
|
Hossain S, Mian IH, Tsuno K (1989). Efficacy of three nematicides and two oilcakes for control of root knot nematode (Meloidogyne incognita) in potato seedlings. Journal of the Faculty of Agriculture, Kyushu University 34(1&2):15-121.
|
|
|
|
|
Huang W, Ji H, Gheysen G, Debode J, Kyndt T (2015). Biochar-amended potting medium reduces the susceptibility of rice to root-knot nematode infections. BMC Plant Biology 15:267.
Crossref
|
|
|
|
|
Hussey RS, Barker KR (1973). A Comparison of Methods of Collecting Inocula for Meloidogyne spp. Including a New Technique. Plant Disease reporter 57:1025-1028.
|
|
|
|
|
International Biochar Initiative (IBI) (2012). Standardized product definition and product testing guidelines for biochar that is used in soil. IBI biochar standards.
|
|
|
|
|
Jaiswal AK, Frenkel O, Elad Y, Lew B, Graber ER (2014). Non-monotonic influence of biochar dose on bean seedling growth and susceptibility to Rhizoctonia solani: the "shifted Rmax-effect". Plant and Soil 395(1-2):125-140.
Crossref
|
|
|
|
|
Kaskavalci G (2007). Effect of soil solarization and organic amendment treatments for controlling Meloidogyne incognita in tomato cultivars in western Anatolia. Turkish Journal of Agriculture and Forestry 31:159-167.
|
|
|
|
|
Khan Z, Kim YH, Kim SG, Kim HW (2007). Observations on the suppression of root-knot nematode (Meloidogyne arenaria) on tomato by incorporation of cyanobacterial powder (Oscillatoria chlorina) into potting field soil. Bioresource Technology 98(1):69-73.
Crossref
|
|
|
|
|
Kolton M, Harel YM, Pasternak Z, Graber ER, Elad Y, Cytryn E (2011). Impact of biochar application to soil on the root-associated bacterial community structure of fully developed greenhouse pepper plants. Applied Environmental Microbiology 77(14):4924-4930.
Crossref
|
|
|
|
|
Lehmann J, Rillig MC, Thies J, Masiello CA, Hockaday WC, Crowley D (2011). Biochar effects on soil biota - a review. Soil Biology and Biochemistry 43:1812-1836.
Crossref
|
|
|
|
|
Liang B, Lehmann J, Sohi SP, Thies JE, O'Neill B, Trujillo L, Gaunt J, Solomon D, Grossman J, Neves EG, Luizão FJ (2010). Black carbon affects the cycling of non-black carbon in soil. Organic Geochemistry 41:206-213.
Crossref
|
|
|
|
|
Luc M, Sikora RA, Bridge J (2002). Plant-parasitic Nematodes in subtropical and tropical agriculture. CAB International, Wallingford, UK. 629p.
|
|
|
|
|
Mai WF, Lyon HH (1975). Pictorial key to genera of plant-parasitic nematodes. No.4th Ed. (revised) P 221.
|
|
|
|
|
Mathews JA (2008). Carbon-negative biofuels. Energy Policy 36:940-945.
Crossref
|
|
|
|
|
Méndez A, Gómez A, Paz-Ferreiro J, Gascó G (2012). Effects of sewage sludge biochar on plant metal availability after application to a Mediterranean soil. Chemosphere 89:1354-1359.
Crossref
|
|
|
|
|
Mitchell PJ, Dalley TS, Helleur RJ (2013). Preliminary laboratory production and characterization of biochars from lignocellulosic municipal waste. Journal of Analytical and Applied Pyrolysis 99:71-78.
Crossref
|
|
|
|
|
Okada H, Harada H (2007). Effects of tillage and fertilizer on nematode communities in a Japanese soybean field. Applied Soil Ecology 35(3):582-598.
Crossref
|
|
|
|
|
Olaoye G, Takim FO, Aduloju MO (2011). Impact of tillage operation on the fruit yield of six exotic tomato varieties on an alfisol in the southern guinea savanna of Nigeria. In: Ogunlela AO (ed.), Proceedings of Nigerian Branch of International Soil Tillage Research Organization. 21st-24th February, Ilorin, Nigeria. pp. 396-403.
|
|
|
|
|
Olowe T (1978). Importance of root-knot nematodes on cowpea Vigna unguiculata (L.) Walp. in Nigeria. In: Proceedings of the International Meloidogyne Project, 3rd Planning Conference on Root-knot Nematodes, 16-20 Nov. 1991, IITA, Ibadan. Nigeria. pp. 85-109.
|
|
|
|
|
Onkendi EM, Kariuki GM, Marais M, Moleleki LN (2014). The Threat of root-knot nematodes (Meloidogyne spp.) in Africa: A review. Plant Pathology 63:727-737.
Crossref
|
|
|
|
|
Osei K, Moss R, Nafeo A, Addico R, Agyemang A, Danso Y and Asante JS (2011). Management of plant parasitic nematodes with antagonistic plants in the forest-savanna transitional zone of Ghana. Journal of Applied Biosciences 37:2491-2495.
|
|
|
|
|
Oyinlola EY, Jinadu SA (2012). Growth, yield and nutrient concentrations of tomato as affected by soil textures and nitrogen. Asian Journal of Agricultural Research 6(1):39-45.
Crossref
|
|
|
|
|
Radwan MA, Farrag SA, Abu-Elamayem MM, Ahmed NS (2012). Efficacy of some granular nematicides against root-knot nematode, Meloidogyne incognita associated with tomato. Pakistan Journal Nematology 30(1):41-47.
|
|
|
|
|
SCANNERS (2009). Understanding the health benefits of tomato products. Sports, Cardiovascular and Wellness Nutrition 2(2):1-4.
|
|
|
|
|
Spokas KA, Reicosky DC (2009). Impacts of sixteen different biochars on soil greenhouse gas production. Annals of Environmental Science 3:179-193.
|
|
|
|
|
Taylor AL, Sasser JN (1978). Biology, Identification and control of root-knot nematodes (Meloidogyne spp.). A Cooperative Publication of the Department of Plant Pathology, North Carolina State University and USAID, Raleigh NC. 111p.
|
|
|
|
|
Thies J, Rilling M, Graber ER (2015). Biochar effects on the abundance, activity and diversity of the soil biota. Biochar for Environmental Management: Science, Technology and Implementation 2:327-389.
|
|
|
|
|
Uzoma KC, Inoue M, Andry H, Fujimaki H, Zahoor A, Nishihara E (2011). Effect of cow manure biochar on maize productivity under sandy soil condition. Soil Use and Management 27:205-212.
Crossref
|
|
|
|
|
Vassilev N, Martos E, Mendes G, Martos V, Vassileva M (2013). Biochar of animal origin: a sustainable solution to the global problem of highâ€grade rock phosphate scarcity? Journal of the Science of Food and Agriculture 93:1799-1804.
Crossref
|
|
|
|
|
Warnock DD, Lehmann J, Kuyper TW, Rillig MC (2007). Mycorrhizal responses to biochar in soil and concepts and mechanisms. Plant and Soil 300:9-20.
Crossref
|
|
|
|
|
Whitehead AG, Hemming JR (1965). A Comparison of Some Quantitative Methods of Extracting Small Vermiform Nematodes from Soil. Annals of Applied Biology 55:25-38.
Crossref
|
|
|
|
|
Williamson VM, Kumar A (2006). Nematode resistance in plants: the battle underground. Trends in Genetics 22(7):396-403.
Crossref
|
|
|
|
|
Yuan JH, Xu RK (2011). The amelioration effects of low temperature biochar generated from nine crop residues on an acidic Ultisol. Soil Use and Management 27:110-115.
Crossref
|
|
|
|
|
Zalkuwi J, Maurice DC, Garba A, Usman J (2012). Profitability of small-scale dry season tomato production in Guyuk local government area of Adamawa State, Nigeria. Journal of Technology and Education Research 5(1):78-82.
Crossref
|
|
|
|
|
Zhang XK, Li Q, Liang WJ, Zhang M, Bao XL, Xi ZB (2013). Soil nematode response to biochar addition in a Chinese wheat field. Pedosphere 23(1):98-103.
Crossref
|
|
|
|
|
ŽnidarÄiÄ D, Trdan S, ZlatiÄ E (2003). Impact of growing methods on tomato (Lycopersicon esculentum Mill) yield and sensory quality. Zb. Biotech. Fak. Univ. Ljublj. Knet 81(2):341-348.
|
|