Full Length Research Paper

ABSTRACT
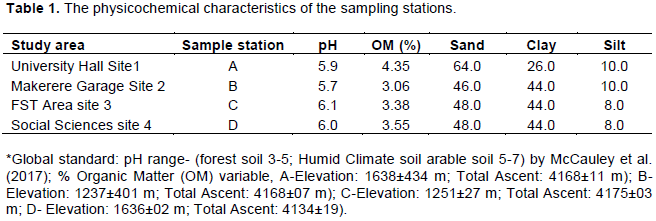
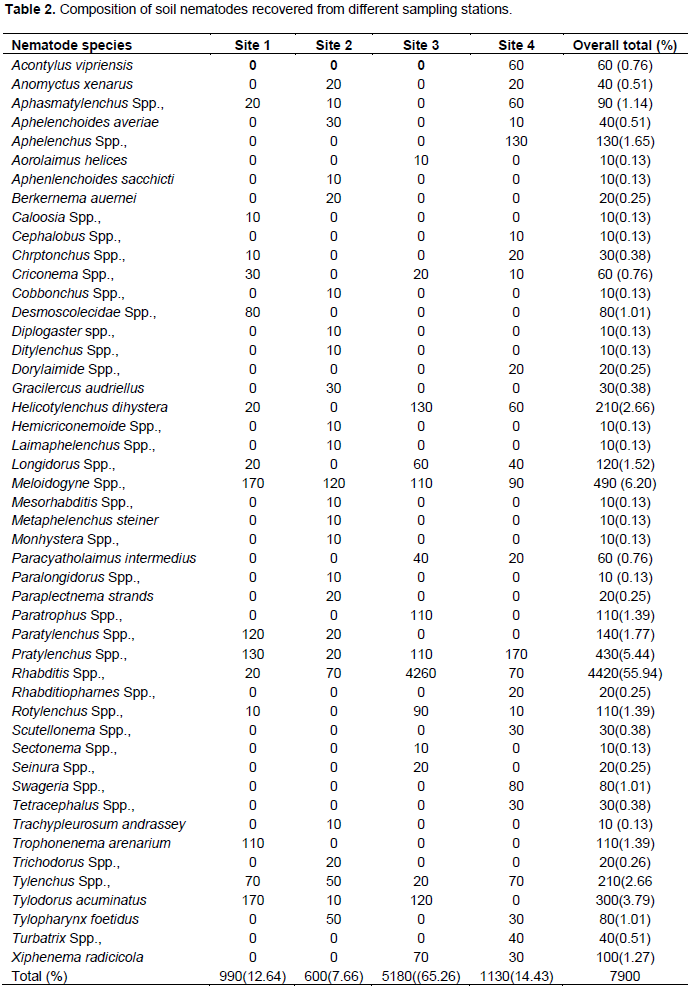
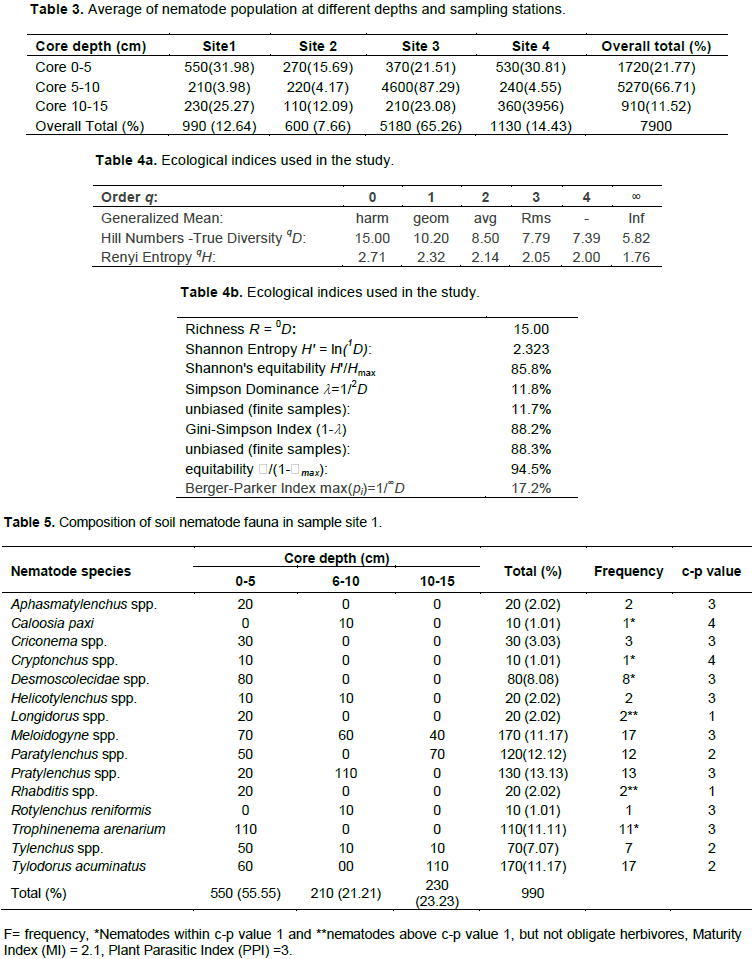
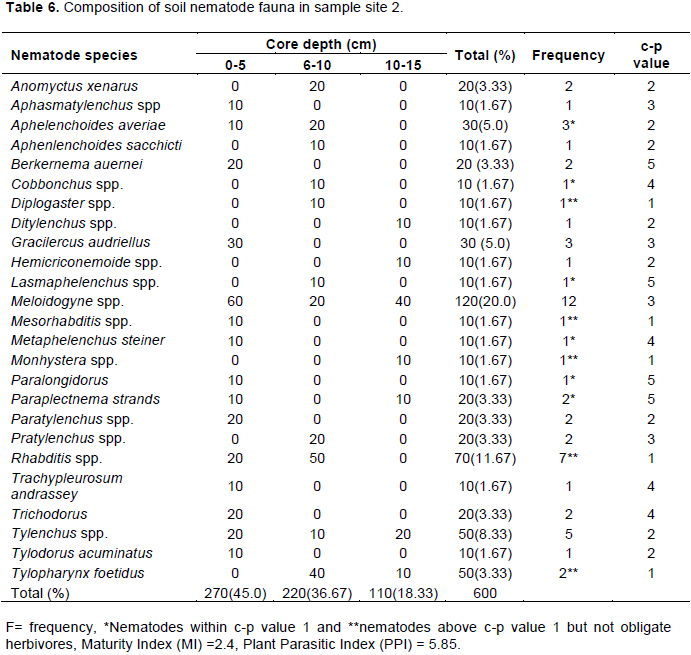
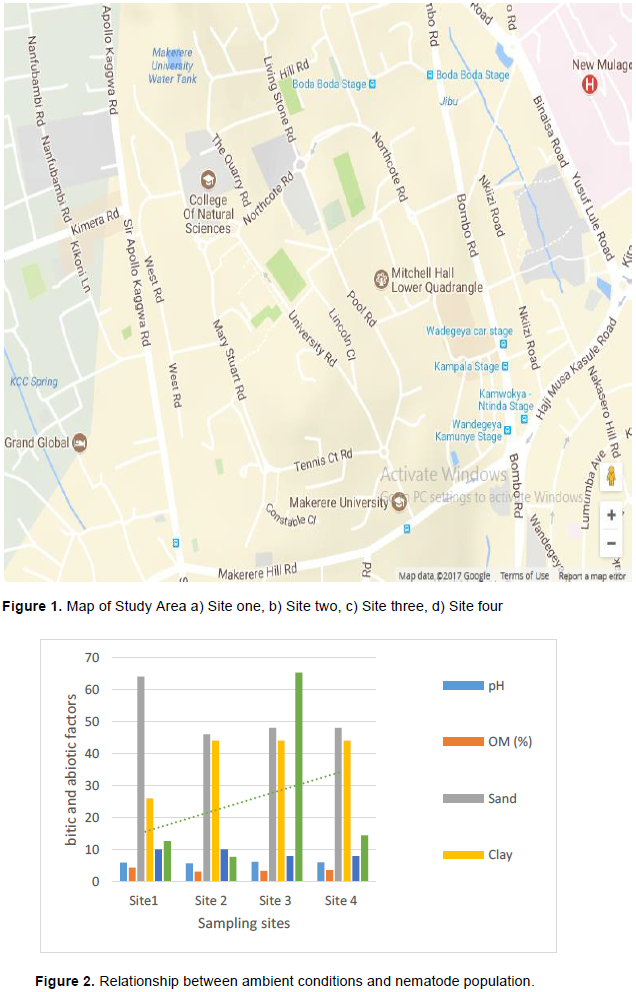
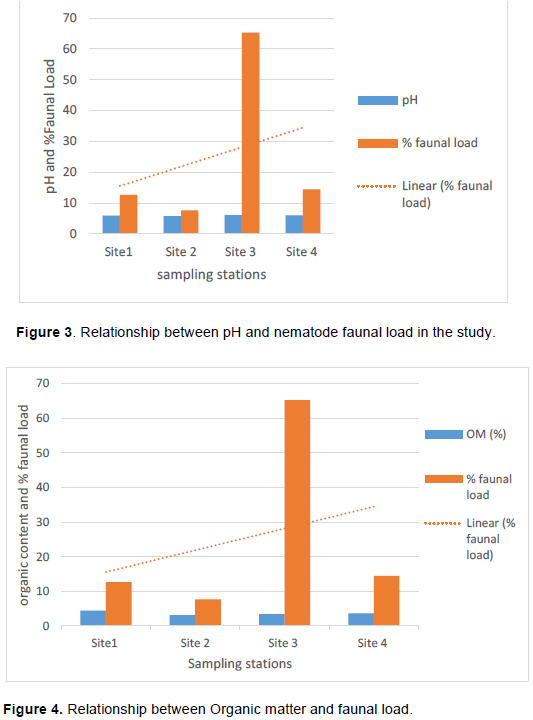
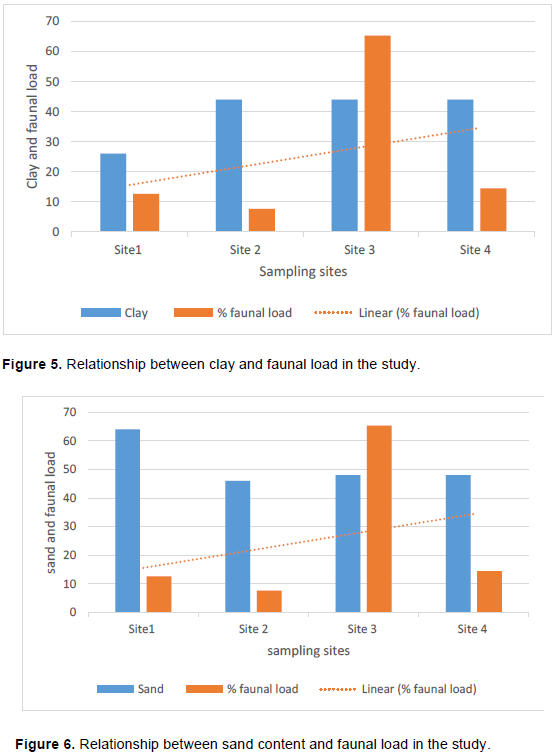
Soil nematode faunal analysis is necessary to ascertain the health status of the soil ecosystem. Composite soil samples were taken at designated sites; A, B, C and D from the Makerere Hill area, Kampala and analyzed to characterize the nematode fauna status. Soil samples were collected vertically at 0-5 cm, 5-10 cm and 10-15 cm core depths with a 5 cm wide soil auger. A total of 7,900 nematodes were collected from the study out of which 1,720 (21.8%) nematodes came from 0-5 cm core depth, 5,270 (66.7) from 5-10 cm core depth and 910 (11.52) from the 10-15 cm core depth. Species diversity showed nine orders of nematodes comprising twenty four families and forty nine species. The Order; Tylenchida had eight families and twenty five species. The Dorylaimida had six families and eleven species, The Rhabditida had families and seven species. The orders; Enoplida, Desmoscolida, Monhysteriida, Chromadorida, Araeolaimida and Tetracephalida had only one family and species each. Nematode species richness and abundance were more in the sites located at the lower fringes of the hill, induced by inherent environmental characteristics that promoted organic enrichment of the soil. The top soil (0-5 cm core depth) comprised the bacterivores c-p 1 (Rhabditis spp.) and c-p 2 nematodes (Desmoscolecidae Spp.), the 5-10 cm core depth had a composite population of all the trophic guilds but dominated by specialist obligates (plant parasitic) while the wide host range obligates (Meloidogyne spp., Pratylenchus spp. and Tylenchus spp.) occurred at 10-15 cm core depth. There was a large assortment of specialist parasites; Aphelenchus spp., Aphelenchoides spp., Aphastimatylenchus nigeriennsis and Trichodorus spp., occasioned by vegetation characteristics of the study area. The study area is a compendium of divergent habitats with peculiar ecomorphological characteristics that can serve as a reference in future environmental impact evaluation studies in relation to soil nematode faunal integrity in Uganda.
Key words: Soil nematodes, species diversity, abundance, bacteriovores, specialist obligates, ecomorpological characteristics.
INTRODUCTION
MATERIALS AND METHODS
|
b
|
|
d
|
RESULTS
DISCUSSION
CONCLUSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
REFERENCES
|
Aakansha G (2014). Kampal; National Capital, Uganda, Encyclopaedia Britanica. Available at: https: |
|
|
Angaye GP, Imafidor HO, Nzeako SO, Angaye TCN (2015a). Bionematicidal Potentials of Azadirachta indica (A. juss), Vernonia amygdalina (DEL), Manihot esculenta, Carica papaya. L. and Citrus sinensis on Meloidogyne incognita of Capsicum annuum, Var. Bell. Journal of Advanced Biological and Basic Research 1(5):9-104. |
|
|
Angaye GP, Imafidor HO, Nzeako SO, Angaye TCN (2015b). Performance Evaluation of Some Potential Bionematicides on Leaf and Fruit Production of Capsicumannuum, Bulletin of Advanced Scientific Research 1(7):151-156. |
|
|
Barker KR, Nusbaum CJ, Nelson LJ (1969). Effects of storage temperature and extraction procedure on recovery of plant-parasitic nematodes from field soils. Journal of Nematology 1:240-247. |
|
|
Barker KR, Carter CC, Sasser JN (1985). An Advanced Treatise on Meloidogyne, volume II Methodology, North Carolina State University Graphics pp. 2-35. |
|
|
Berkelmans R, Ferris H, Tenuta M, van Bruggen AHC (2003). Effects of long-term crop management on nematode trophic levels other than plant feeders disappear after 1 year of disruptive soil management. Applied Soil Ecology 23:223-235. |
|
|
Briar SS, Barker C, Tenuta, M, Entz MH (2012). Soil nematode responses to crop management and conversion to native grasses. Journal of Nematology 44(3):245-254. |
|
|
Bongers T (1990). The maturity index: an ecological measure of environmental disturbance based on nematode species composition. Oecologica 83:14-19. |
|
|
Bongers T, Bongers M (1998). Functional diversity of nematodes. Applied Soil Ecology 10:239-251. |
|
|
Bongers T, de Goede RGM, Korthals G, Yeates GW (1995). Proposed changes of c-p classification for nematodes. Russian Journal of Nematology 3:61-62. |
|
|
Bongers T, Rob AE, Yeates GW (1999). Interpretation of disturbance-induced maturity decrease in marine nematode assemblages by means of the maturity index. Marine Ecology Progress Series 176:135-142. |
|
|
Bongers T, van der Meulen H, Korthals G (1997). Inverse relationship between the nematode maturity index and plant parasite index under enriched nutrient conditions. Applied Soil Ecology 6:195-199. |
|
|
Bongers T, Alkemade R, Yeates GW (1991). Interpretation of disturbance-induced maturity decrease in marine nematode assemblages by means of the Maturity Index. Marine Ecology Progress Series 76:135-142. |
|
|
Shabeg BS, Barker C, Tenuta M, Martin HE (2012). Soil Nematode Responses to Crop Management and Conversion to Native Grasses. Journal of Nematology 44(3):245-254. |
|
|
Cesarz S, Reich PB, Scheu S, Ruess L, Schaefer M, Eisenhauer N (2015). Nematode functional guilds, not trophic groups, reflect shifts in soil food webs and processes in response to interacting global change factors. Journal of Soil Ecology Pedobiologia 58:23-32. |
|
|
Coyne DL, Nicol JM, Claudius-Cole B (2007). Practical plant nematology: A field and laboratory guide, P-IPM Secretariat, International Institute of Tropical Agriculture (IITA), Cotonou, Benin. |
|
|
Elele K, Nzeako SO, Asor JE (2017). Nematode Occurrence and Distribution in an Organically Managed Solanum Melongena L Field. Scientia Agriculturae 20(3):71-80. |
|
|
Ferris H, Bongers T (2006). Nematode indicators of organic enrichment. Journal of Nematology 38:3-12. |
|
|
Ferris H, Venette RC, Scow KM (2004). Soil management to enhance bacterivore and fungivore nematode populations and their nitrogen mineralization function. Applied Soil Ecology 24:19-35. |
|
|
Ferris H (2010). Contribution of nematodes to the structure and function of the soil food web. Journal of Nematology 42(1):63-67. |
|
|
Ferris H, Benkelman K (2003). Changes in estuarine Nematode communities following a decrease of organic pollution. Aquatic Ecology 32(3):195-202. |
|
|
Ferris H, Bongers T (2009). Indices for analysis of nematode assemblages, In: Wilson M, Kakouli-Duarte T (Eds.) Nematodes as Environmental Bioindicators. CABI Wallingford, U.K. pp. 124-145. |
|
|
Ferris H, Bongers T, de Geode RGM (2001). A framework for soil food web diagnostics: extension of the nematode faunal analysis concept. Applied Soil Ecology 18:13-29. |
|
|
Ferris H, Griffiths BS, Porazinska DL, Powers TO, Wang KH, Tenuta M (2012). Reflections on Plant and Soil Nematode Ecology: Past, Present and Future. Journal of Nematology 44(2):115-126. |
|
|
Fiscus DA, Neher DA (2002). Distinguishing Sensitivity of free-living soil nematode genera to physical and chemical disturbances. Ecology Applied 12:565-575. |
|
|
Goodey T, Goodey JB (1963). Soil and freshwater Nematodes. Butler and Tanner Ltd, London, Great Britain. |
|
|
Haegeman A, Mantelin S, Jones JT, Gheysen G (2012). Functional roles of effectors of plant-parasitic nematodes. Gene 492:19-31. |
|
|
Hagerbaumer A, Höss S, Heininger P, Traunspurger W (2015). Experimental Studies with Nematodes in Ecotoxicology: An Overview. Journal of Nematology 47(1):11-27. |
|
|
Heip C, Vincx M, Aschner M (2000). The ecology of marine nematodes. Ocean Marine Biology Annual Reviews 23:399-489. |
|
|
Heip C, Vincx M, Vranken G (1985). The ecology of marine nematodes. Oceanography and Marine Biology Annual Review 23:399-489. |
|
|
Herris H, Bongers T (2009). Indices developed specifically for analysis of nematode assemblages, In: Nematodes as Environmental Indicators (eds.), Wilson MJ, Kakouli DT CAB International, Wallingford pp. 124-145. |
|
|
Imafidor HO, Angaye GP, Nzeako SO (2016). The Effects of some bio-nematicides on the Productivity of Capsicum annum. Journal of Environmental Treatment Techniques 4(4):111-117. Available at: |
|
|
Kimenju J, Sibanda Z, Talwana H, Wanjohi W (2007). Nematology Training Manual, NIESA Training Manual, Funded By NIESA and University of Nairobi, Crop Protection Department. |
|
|
Korner C (2007). The use of 'altitude' in ecological research. Trends in Ecology and Evolution 22:569-574. |
|
|
Manzanilla-Lopez RH, Hunt JD (2004). Tropical Plants and Nematode Diversity and Interaction. Tropical Biology and Conservation Management Vol. 5. |
|
|
McCauley A, Jones C, Olson-Rutz K (2017). Soil pH and Organic Matter. Nutrient Management Module 8:1-12. |
|
|
Mulder C, Schoutena AJ, Hund-Rinkeb K, Breure AM (2005). The use of nematodes in ecological soil classification and assessment concepts. Ecotoxicology and Environmental Safety 62(2):278-289. |
|
|
Mwamba S (2016). Root knot nematodes (Meloidogyne incognita) interaction with selected Asteraceae plants and their potential use for nematode management. A dissertation submitted in partial fulfilment for the Degree of a Master of Science in Research methods in the department of Horticulture of Jomo Kenyatta University of Agriculture and Technology pp. 1-87. |
|
|
Nzeako SO, Imafidor HO, Ogwumba E, Ezenwaka CO (2016). Vertical Distribution of the Lesion nematode: Pratylenchus species in selected turf fields in Rivers State, Nigeria. 9(7):53-58. |
|
|
Nzeako SO, Imafidor HO, Uche AO, Ogufere MO (2014). Agro-related anthropogenic activities on soil nematodes in the Niger Delta, Nigeria. International Journal of Applied Science Research Review 1(3):161-169. |
|
|
Nzeako SO, Imafidor HO, Ihenacho PC (2011). Effect of Crude Oil spillage on soil Nematodes community composition. Bioscience Research Journal 23(3):141-145. |
|
|
Nzeako S, Imafidor H, Uche A, Udofia I (2015). An Assessment of the Nematode Population in the Benthic Zone of a Tidal Freshwater Body using the Lower Reaches of the New Calabar River as a Case Study. Journal of Biology, Agricure and Healthcare 5(17):109-112. |
|
|
Okalebo JR, Gathua KW, Woomer PI (2002). Laboratory methods of soil and plant analysis: a working manual (2nd ed.) Sacred African Publishers, Nairobi, Kenya. |
|
|
Pen-Mouratov S, Shukurov N, Steinberger Y (2010). Soil free living nematodes as indicators of both industrial pollution and livestock activity in Central Asia. Ecological Indicators 10:955-967. |
|
|
Popovici I, Ciobanu M (2000). Diversity and distribution of nematode communities in grasslands from Romania in relation to vegetation and soil characteristics. Applied Soil Ecology 14:27-36. |
|
|
Raquel LS (2012). Nematodes as soil quality indicators in coffee systems, M.Sc. Thesis report, farming systems ecology, Wageningen University, The Netherlands Droevendaalsesteeg 1-6708, Pb Wageningen. |
|
|
Renco M, Kovacik P (2012). Response of Plant Parasitic and Free Living Soil Nematodes to Composted Animal Manure Soil Amendments, Journal of Nematology 44(4):329-336. |
|
|
Seiyaboh EI, Ogamba EN, Utibe DI (2010). Impact of Dredging on the Water Quality of Igbedi Creek, Upper Nun River, Niger Delta, Nigeria. Journal of Environmental Science, Toxicology and Food Technology 7(5):51-56. |
|
|
Talwana H, Sibanda Z, Wanjohi W, Kimenju W, Luambano-Nyoni N, Massawe C, Manzanilla-Lopez R, Davies KG, Hunt DJ, Sikora RA, Coyne LD, Gowen SR, Kerry BR (2015). Agricultural nematology in East and Southern Africa: problems, management strategies and stakeholder linkages. Pest Management Science 72:2. |
|
|
Thorne G (1961). Principles of nematology. McGraw- Hill, New York P 553. |
|
|
Traunspunger W (2002). Nematoda. pp. 63-104, In: Rundle SD, Robertson A. and Schmid-Araya J, eds. Freshwater meiofuana: Biology and Ecology. Leiden, The Netherlands; Blachuys Publishers. pp. 63-104. |
|
|
Traunspurger W, Bergtold M, Ettermeyer A, Goedkoop W (2006). Effects of copepods and chironomids on the abundance and vertical distribution of nematodes in a freshwater sediment. Journal of Freshwater Ecology 21(1):81-90. |
|
|
Urkmez D, Sezgin M, Bat L (2014). Use of nematode maturity index for the determination of ecological quality status: a case study from the Black Sea. Mediterranean Environment 20(2):96Ë—107. |
|
|
Von der Ohe PC, Goedkoop W (2013). Distinguishing the effects of habitat degradation and pesticides stress on benthic invertebrates using stressor- specific metrics. Science of the Total Environment 444:480-490. |
|
|
Von der Ohe PC, Prub A, Schafer RB, Liess M, de Decker E, Brack W (2007). Water quality indices across Europe- a comparison of the good ecological status of five river basins. Journal of Environmental Monitoring 9:970-978. |
|
|
Wang KH, McSorley R, Gallaher RN (2009). Can nematode community indices elucidate plant health conditions? Journal of Nematology 41:392. |
|
|
Warwick RM (1986). A new method for detecting pollution effects on marine macrobenthic communities. Marine Biology 92:557-562. |
|
|
Xiaoming S, Zhang X, Zhang S, Dai G, Liang SHW (2013). Soil Nematode Responses to Increases in Nitrogen Deposition and Precipitation in a Temperate Forest. PLOS ONE 8(12):1-8. |
|
|
Yeates GW (2007). Abundance, diversity, and resilience of nematode assemblage in forest soils. Canadian Journal for Research 37:216-225. |
|
|
Yeates GW, Wardle DA, Watson RN (1999). Response of soil nematode populations, community structure, diversity and temporal variability to agricultural intensification over a seven-year period. Soil Biology and Biochemistry 31:1721-1733. |
|
|
Zhang M, Liang WJ, Zhang XK (2012). Soil Nematode Abundance and Diversity in Different Forest Types at Changbai Mountain, China. Zoological Studies 51(5):619-626. |
|
|
Zhu B, Wang XP, Fang JY, Piao SL, Shen HH, Zhao SQ, Peng CH (2010). Altitudinal changes in carbon storage of temperate forests on Mt. Changbai, Northeast China. Journal of Plant Research 123:439-452. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


