Full Length Research Paper

ABSTRACT
Large-scale coal mining practices, particularly surface coal extraction and associated valley fills as well as residential wastewater discharge, are of ecological concern for aquatic systems in central Appalachia. Identifying and quantifying alterations to ecosystems along a gradient of spatial scales is a necessary first-step to aid in mitigation of negative consequences to aquatic biota. In central Appalachian headwater streams, apart from fish, salamanders are the most abundant vertebrate predator that provide a significant intermediate trophic role linking aquatic and terrestrial food webs. Stream salamander species are considered to be sensitive to aquatic stressors and environmental alterations, as past research has shown linkages among microhabitat parameters, large-scale land use such as urbanization and logging, and salamander abundances. However, there is little information examining these relationships between environmental conditions and salamander occupancy in the coalfields of central Appalachia. In the summer of 2013, 70 sites (sampled two to three times each) in the southwest Virginia coalfields were visited to collect salamanders and quantify stream and riparian microhabitat parameters. Using an information-theoretic framework, effects of microhabitat and large-scale land use on stream salamander occupancy were compared. The findings indicate that Desmognathus spp. occupancy rates are more correlated to microhabitat parameters such as canopy cover than to large-scale land uses. However, Eurycea spp. occupancy rates had a strong association with large-scale land uses, particularly recent mining and forest cover within the watershed. These findings suggest that protection of riparian habitats is an important consideration for maintaining aquatic systems in central Appalachia. If this is not possible, restoration riparian areas should follow guidelines using quick-growing tree species that are native to Appalachian riparian areas. These types of trees would rapidly establish a canopy cover, stabilize the soil, and impede invasive plant species which would, in turn, provide high-quality refuges for stream salamanders.
Key words: Canopy cover, coal mining, Desmognathus, Eurycea, headwater streams, surface mining.
INTRODUCTION
Amphibians, particularly salamanders, are an important component to both terrestrial and aquatic ecosystems (Davic and Welsh 2004). Long-lived stream salamander species are a keystone faunal group, can reach high densities in undisturbed areas, and their life history traits make them sensitive to both stream and upland watershed alterations (Welsh and Ollivier, 1998; Welsh and Droege, 2001; Southerland et al., 2004). In headwater streams, apart from fish, salamanders are the most abundant vertebrate predators. Stream salamanders provide a significant intermediate role in stream food webs, feeding on small prey such as benthic macroinvertebrates and are an important prey item for larger vertebrates (Petranka et al., 1993; Davic and Welsh, 2004; OHEPA, 2012). Undisturbed riparian areas of central Appalachia have relatively stable, intra- and inter-annual salamander populations (as compared to macroinvertebrates), with densities of up to 1.4/m2 (Hairston and Wiley, 1993). Movement of many Appalachian stream salamander species has been shown to be less than 100 m (Pauley et al., 2000), and although adult stream salamanders are able to cross barriers, such as dry streambed sections and waterfalls, mobility may be limited by riparian and upper watershed habitat fragmentation (Grant et al., 2010; Resetarits 1997; Willson and Dorcas, 2003; OHEPA, 2012). Their low mobility reduces the likelihood of abundance being a reflection of immigration, which helps confirm linkages between local, long-term stream and watershed conditions and salamander survival and reproduction (Welsh and Ollivier, 1998; Welsh et al., 2005). Life history and behavior vary greatly among aquatic salamander species with some larval forms having aquatic phases of up to 48 months, whereas other species have very short- or even no aquatic larval phase (OHEPA, 2012). This biphasic life history trait shown by some salamander species can provide a gradient of responses to aquatic and upland environmental conditions giving better insight to both terrestrial and stream conditions (Fisher et al., 1998; Petranka, 1998). For example, some species of stream salamander abundances may be reduced, even to the point of local extirpation, by decreases in physiochemical stream and riparian conditions from watershed land uses such as mining, urbanization and timber harvesting that cause changes in pH, streambed sedimentation, suspended sediment, and water and soil temperature (Willson and Dorcas, 2003; Welsh et al., 2005; Moseley et al., 2008). Accordingly, other species of stream salamanders may be very tolerant of habitat degradation. Therefore, variance in species composition may be indicative of stream condition and environmental health.
Research has tried to better quantify the effects of anthropogenic land use on salamander abundance, density and most recently, occupancy. In the eastern United States, majority of this work has focused on the effects of forestry practices in relation to salamander abundances. In the central and southern Appalachians, data suggest forestry practices at the stand and landscape scale, at least in the short-term, lead to declines in terrestrial salamander abundance until sufficient forest maturation occurs (Ash, 1988; Petranka et al., 1993; Harper and Guynn, 1999; Ford et al., 2002). At smaller spatial scales, logging-related alterations to riparian quality (reduced canopy cover, leaf litter depth and decreased soil moisture) along with the alteration of physical stream conditions (change in stream substrate class, substrate embeddedness, riffle/run/pool composition) negatively influence stream salamander populations (Crawford and Semlitsch, 2008; Moseley et al., 2008; Peterman and Semlitsch, 2009). Stream salamanders may be significantly affected by fine spatial scales (microhabitat) more than landscape-level land use; however, microhabitat conditions are often directly related to cumulative historical and current watershed land use (Bury and Corn, 1989; Russell et al., 2004; Surasinghe and Baldwin, 2014). These studies illustrate the need for better understanding the dynamics among salamander occupancy, large-scale land use, and microhabitat, especially for Appalachian salamanders given the limited data available.
There has been minimal effort to examine the effects of coal mining on stream salamanders relative to forestry research. Most of the amphibian research on mined lands has focused on constructed settling ponds and use and recolonization by pond salamanders such as those in the family Ambystomidae (Fowler et al., 1985; Lacki et al., 1992; Jansen et al., 2004). Other work has centered on acid mine drainage because of its direct effects on stream biota, including stream salamanders (Freda, 1986; Middlekoop et al., 1998; Schorr et al 2013). However, acid mine drainage is often a localized issue rather than pervasive throughout all the Appalachian coalfields (Herricks and Cairns, 1974; Minear and Tschantz, 1976).
Salamander community richness and abundance is generally lower in valley fill streams (aquatic systems buried by surface overburden during the mining process) than reference streams in central Appalachia (Hamilton, 2002; Wood and Williams, 2013a; Muncy et al., 2014). However, Hamilton (2002) observed no difference in 18 year old valley fill streams versus unmined reference streams. Nonetheless, complete examination linking microhabitat to landscape-level conditions and stream salamanders has rarely occurred. The authors’ main objective was to investigate the relationship between salamander communities and habitat parameters at both a microhabitat and a landscape-level along a gradient of conditions in the coalfields of central Appalachia. These foundational correlations with an information-theoretic occupancy/detection modeling approach were examined using microhabitat data collected in the field along with a geographic information system (GIS) analysis of landscape-level land use.
MATERIALS AND METHODS
In southwest Virginia, five 12-digit Hydrologic Unit Codes (HUC-12) watersheds of similar area and located in the Cumberland Plateau and Mountains Range (Ecoregion 69d) of the central Appalachians were selected. Callahan Creek (area = 54.7 ha), Pigeon Creek (area = 58.9 ha), Roaring Fork (area = 66.0 ha) and Rock Fork (area = 91.0 ha) watersheds are located in Wise County, Virginia. Dumps Creek (area = 82.3 ha) is located in Dickenson and Russell Counties, Virginia. These watersheds have been substantially altered from past coal mining and residential areas as well as being actively mined for coal. The forested areas of the watersheds are characterized by a diverse mix of hardwood and conifers (Woodward and Hoffman, 1991). First- or second-order stream segments were randomly selected for salamander sampling sites; however, in some cases, best professional judgment was used to select sites that had landowner access and that considered safe to sample (Sweeten 2015). In total, 70 sites were sampled within the five HUC-12 watersheds. Because of the difficulty in accessing sites, the number of sites within each HUC-12 was not equal: 20 sites were sampled within Callahan Creek, 15 sites in Roaring Fork, 15 sites in Rocky Fork, 10 sites in Pigeon Creek, and 10 sites in Dumps Creek.
The authors attempted to sample each site three times between May 15 and August 15, 2013. However, because of access issues, three sites were only sampled twice in 2013. Two quadrats (25 m long by 5 m wide) were placed parallel to the stream at every sampling location with the edge of each quadrat placed along the center of the stream (Figure 1). At each sampling event, salamanders were captured from either the left or right quadrat (facing upstream) which was determined randomly. All rocks, logs and detritus within the quadrat were overturned in order to capture adult salamanders (all transformed salamanders, sexually mature and immature, were considered adults). Dip nets were used to capture larval salamanders within the stream. adult salamanders were identified to the species level, and larval salamanders to the genus-level. All salamanders were then released within a meter of their capture location.
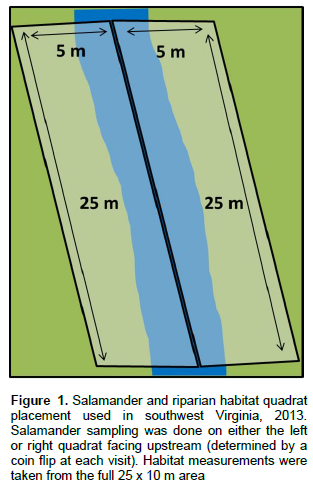
Microhabitat parameters
The two 25 m by 5 m quadrats were combined for measuring riparian conditions as well as physical and chemical stream attributes. Past research was reviewed to identify microhabitat parameters thought to influence salamander occupancy and/or abundance (Bury and Corn, 1989; Miller et al., 2007; Moseley et al., 2008; Kroll et al., 2010). Temporally variable parameters such as water temperature, conductivity, soil temperature, air temperature and stream flow condition were sampled at each of the three sampling events. Parameters thought to likely remain constant over the field season such as percent canopy cover, tree species richness and large woody debris (LWD) counts, were measured once during the summer. More details on collection methods for all microhabitat parameters are shown in Appendix 1.
Spatial analysis
Using multiple GIS layers such as the 2011 National Agriculture Imagery Program (NAIP), mining GIS data from the Virginia Department of Mines Mineral and Energy (DMME), the 2011 National Land Cover Dataset (NLCD), the USGS National Elevation Dataset (NED), the USGS National Hydrological Dataset (NHD), historical topographical maps and land use cover data from Maxwell et al. (2014), the authors measured landscape-level parameters at the subwatershed (the watershed from above the sampling location) scale and HUC-12 watershed scale. Parameters included watershed area, length of stream segment, kilometers of roads, area of valley fill, and number of structures and constructed ponds within the subwatershed. Land use was classified as percent of each watershed with recent mining (active mining permits and areas still barren), past mining and forested. All GIS analysis were performed using ArcMap2 (v10.1, ESRI, Redlands, California). More details on the methods for the GIS analysis of landscape-level parameters are shown in Appendix 2.
Statistical analysis
The Program PRESENCE software (Any use of trade, firm or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government) (v7.3, MacKenzie et al., 2006) was used to assess salamander occupancy and detection probabilities in an information-theoretic framework. Program PRESENCE fits multiple models and Akaike’s Information Criterion (AIC) values and maximum likelihood were used to rank all detection (p) and occupancy (psi, Ψ) parameters relative to environmental condition (Kroll et al., 2010). If detection is unaccounted for, the true presence of a species may be misclassified as absent if a species was present but not detected. Consequently, occupancy estimates can be skewed (MacKenzie, 2006).
Model selection
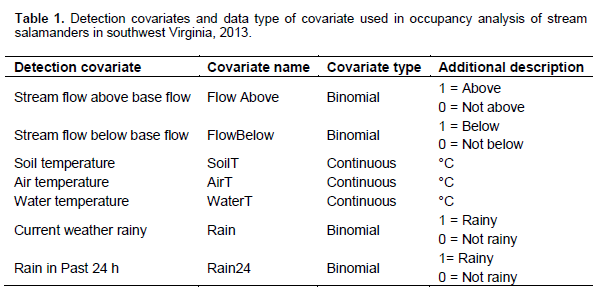
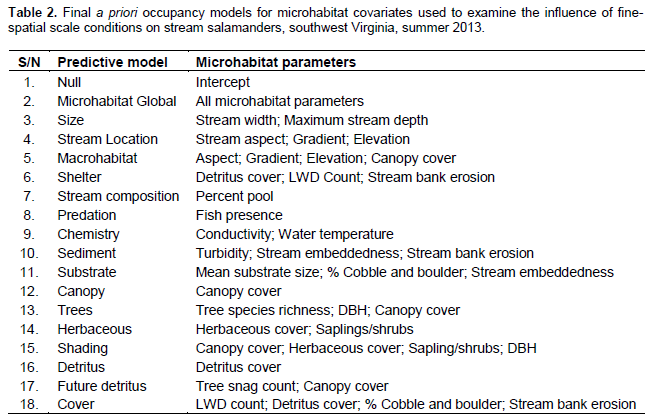
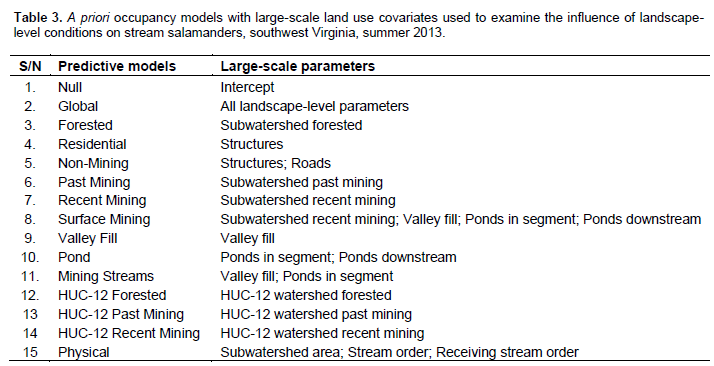
A two-step method was used to determine which detection covariates to include for each species in the occupancy analysis (Burnham and Anderson, 2002). A priori detection covariates was ran for each species against the null (intercept) model. All detection covariates that had an AIC smaller than the null were then run for all combinations to determine the best detection covariate for each species. This detection covariate was then used for the occupancy analysis. Utilizing this two-step method helps to reduce the total number of models in the final analysis. For this study, seven detection covariates were analyzed (Table 1). Prior to analysis, continuous detection covariates was normalized. 18 a priori models (containing 21 parameters) were developed for microhabitat occupancy analysis (Table 2). 15 landscape-level a priori models were developed using 14 GIS-derived covariates (Table 3). For both sets of models, covariates were grouped based on biologically relevant information or recommendations from available literature (Petranka et al., 1993; Ford et al., 2002; Willson and Dorcas, 2003; Moseley et al., 2008). All combinations of the parameters were not considered as this would have created unreliable outputs (Burnham and Anderson, 2002). Prior to analysis, all continuous occupancy covariates were normalized (Sweeten, 2015).
RESULTS
Species selection
Overall, nine species of aquatic salamanders were detected during the 207 surveys (Table 4). Because of the large number of Eurycea spp. larval salamanders (442 individuals) and the small number of adult Eurycea spp. (39 individuals), larval and adult Eurycea longicauda (Long-tailed Salamander) and Eurycea bislineata cirrigera (Southern Two-lined Salamander) salamanders were combined to the genus-level for the Eurycea spp. group. All the larval and adult Desmognathus were also combined to the genus-level to make the Desmognathus spp. group. Based on estimated occupancy and detection probabilities, five salamander groups had sufficient data for occupancy analysis: Desmognathus fuscus (Northern Dusky Salamander), Desmognathus monticola (Seal Salamander), Desmognathus ochrophaeus (Mountain Dusky Salamander), Desmognathus spp. and Eurycea spp. (Table 5). However, the model fit (c-hat) for the Desmognathus spp. group was over-dispersed, and therefore, excluded from analysis. Species or genus groups that were detected too infrequently to be included in the occupancy analysis were Desmognathus welteri (Black Mountain Salamander), Desmognathus spp. larval, Eurycea spp. adults, Eurycea bislineata cirrigera, Eurycea longicauda, Gyrinophilus porphyriticus (Spring Salamander), Pseudotriton montanus (Mud Salamander) and Pseudotriton ruber (Red Salamander) (Table 4).
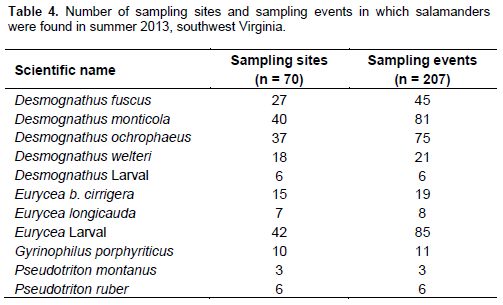
Occupancy results
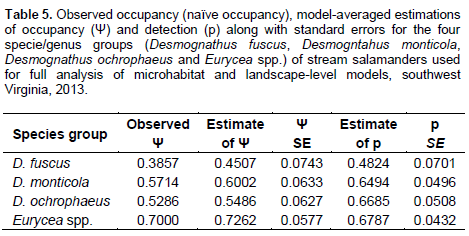
In comparison of microhabitat and landscape-level models, the models with either strong empirical support (within Δ AIC of < 2) or moderate empirical support (Δ AIC of 2 – 4) for D. fuscus were all microhabitat models (Table 6). The canopy model showed the most empirical support with an AIC weight of 0.2994 (Table 6). The percent canopy cover covariate was in five of the six top models for D. fuscus (Table 7). With the exception of the Non-Mining Model (a landscape-level model) with a Δ AIC of 2.78, all models with empirical support (within Δ AIC of 4) for D. monticola were models with microhabitat or small-scale components (Table 6). The Shelter Model was the top ranked model with an AIC weight of 0.2895 (Table 6). The covariates percent detritus cover and the number of large woody debris pieces (LWD) were present in two of the top three models, and were positively associated with D. monticola occupancy (Table 7). D. ochrophaeus results had empirical support for microhabitat models (Table 6). The best-ranked model was the Macrohabitat Model with an AIC weight of 0.4202
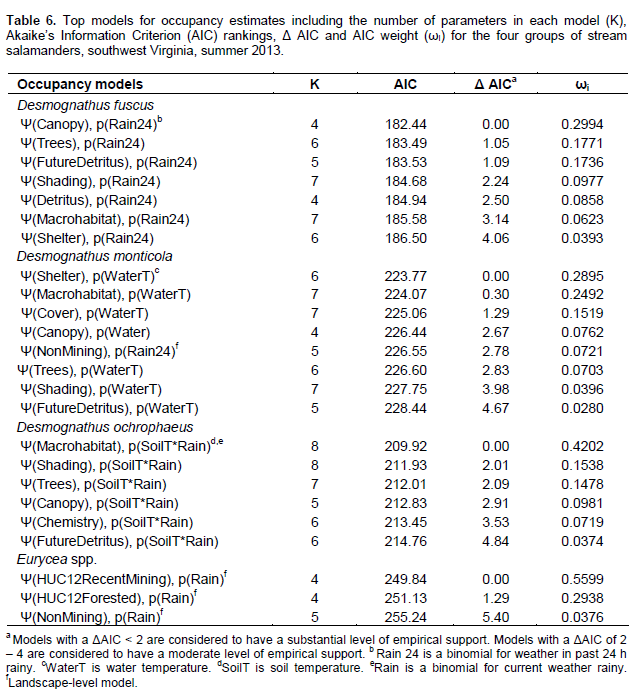
(Table 6), and percent canopy cover covariate had a positive relationship with D. ochrophaeus occupancy (Table 7). Landscape-level models had empirical support for Eurycea spp. (Table 6). The HUC-12 Recent Mining Model is the top model with an AIC weight of 0.5599 (Table 6). Within this model, the percent of recent mining within the HUC-12 watershed showed a negative correlation with Eurycea spp. occupancy probabilities (Table 8).

DISCUSSION
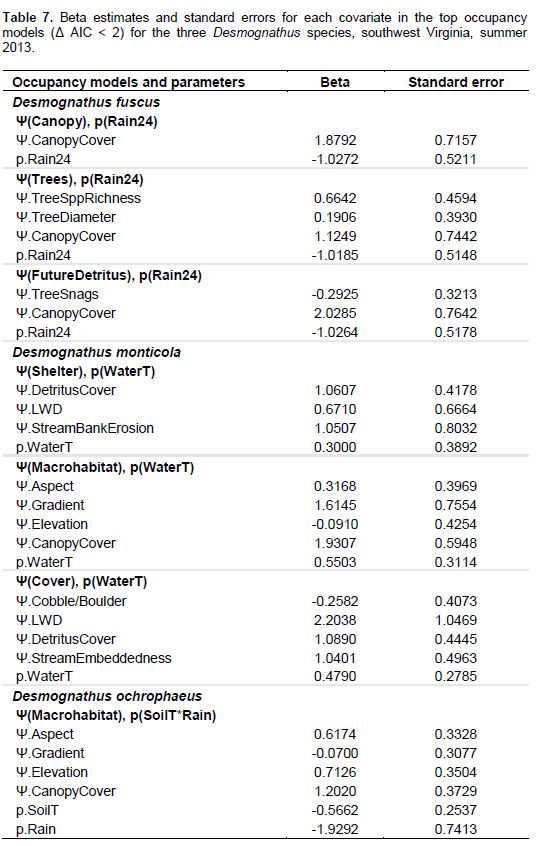
It was found that Desmognathus spp. had similar results with most top models being microhabitat models. Within the microhabitat models, most top models (within a Δ AIC of 4) contained canopy cover as a covariate. Five occupancy models best explained presence of the three Desmognathus species. These models were the Canopy Model, the Trees Model, the Shading Model, the Shelter Model and the Macrohabitat Model. Multiple parameters comprised these models including canopy cover, tree diameter at breast height (DBH), tree species richness, herbaceous cover, sapling/shrub density, detritus cover, LWD and stream bank erosion. Because data were normalized prior to analysis, it was possible to compare the betas, or effect size, of these different parameters and examine correlation trends among models and species. Canopy cover was a parameter in four of the five models.
All the three Desmognathus species were positively correlated with increased canopy cover. Additionally, beta values for canopy cover in the top models were large, indicating a strong effect size. Past research has also shown strong associations between canopy cover and salamander abundance (Davic and Welsh, 2004; Crawford and Semlitsch, 2008; Ward et al., 2008). Desmognathus salamanders are lungless and thereforeare required to constantly have moist skin in order for oxygen exchange across the skin membrane (Petranka, 1998). Lungless salamanders have a high risk of desiccation particularly when foraging away from the stream. Canopy cover not only provides cover from solar exposure lowering soil, stream, and air temperatures, it also increases other microhabitat parameters associated with maintaining a cool, moist environment such as detritus cover. Crawford and Semlitsch (2008) found a positive correlation in riparian areas between detritus depth and D. monticola and E. b. cirrigera. In upland plots in the southern Appalachians of North Carolina, Harper and Guynn (1999) observed more salamanders including D. ochrophaeus and D. aeneus in moist microhabitats with increased detritus depths.
A closed canopy cover is also often an indication of mature, less-disturbed or more recovered forest conditions. Although, forest stand age was not a covariate in this study, Desmognathus spp. were positively correlated with characteristics often found in more mature forests such as high canopy cover, native tree species dominance, high detritus cover and more LWD. In nearby Ecoregions 67 and 69 of West Virginia, Moseley et al. (2008) found a positive relationship between Desmognathus spp. abundance in first- and second-order streams and time since forest harvest in the immediate watershed. Ford et al. (2002) observed that Desmognathus spp. abundance generally was most correlated to tree basal area within forest stands in southern Appalachia. Additionally, in a review of North American literature on amphibian ecology and forest management, deMaynadier and Hunter (1995) suggested increased salamander abundance in older forests functionally are an indirect measure of microhabitat conditions such as LWD, detritus cover and canopy cover.
Results from the present study show that Desmognathus were negatively correlated with parameters associated with open areas/grasslands such as increased herbaceous cover and areas dominated by invasive species. In Maryland, Walz (2002) found decreased abundances of D. fuscus and D. ochrophaeus in agricultural fields and pastures. Wood and Williams (2013b) found lower abundances of Desmognathus in reclaimed grassland and shrublands in the West Virginia coalfields (Ecoregion 69) where there was less detritus, lower stem densities, less LWD, less canopy cover, and an increase in invasive herbaceous species such as Lespedeza spp. as compared to forested or partially forested sites. Invasive herbaceous species could be indicative of recent disturbance, or it may be that established invasive plant species simply do not produce the necessary microhabitat (leaf litter, cover, and LWD) to provide the cool, moist habitat needed for salamanders (Lemke et al. 2012).
With the exception of the Non-Mining Model for D. monticola, large-scale land use models were not supported for Desmognathus spp. occupancy. This indicates that local riparian areas are more important for Desmognathus spp. than watershed or subwatershed land uses. In the Ecoregion 69 portion of Kentucky, Maigret et al. (2014) found that Desmognathus spp. abundances were lower in areas without a timber harvest stream buffer zone than in undisturbed areas or areas with a 7.6 m stream buffer zone. Pearl et al. (2005) found that in the Pacific Northwest wetland and riparian conditions are most influenced by native salamander species occupancy and land use within a 1000 m radius had little effect on salamander occupancy.
The Non-Mining Model for D. monticola was the only large-scale land use with empirical support for the Desmognathus spp. This model consists of the number of houses and total km of roads in the subwatershed. Beta results show that the kilometers of road had a much larger effect size than the number of houses. Similarly, Ward et al. (2008) also found a negative correlation between Desmognathus spp. abundance and roads in central Appalachia. Roads often tend to cause forest fragmentation, which decreases canopy cover and detritus (Ward et al., 2008). Additionally, deMaynadier and Hunter (2000) found that forested sites without roads generally had twice the number of salamanders than roadside sites. They suggested that movement of both terrestrial and aquatic salamanders may be limited by roads. Impassible culverts can impede upstream salamander movement, and may prevent migration altering salamander community structure upstream of road-stream crossings (Ward et al., 2008; Anderson et al., 2014).
Roads, particularly paved roads, may also alter water chemistry parameters such as conductivity. Runoff from roads containing de-icing salt has been shown to travel over 170 m from the road into wetlands in the Adirondack Mountains of New York (Karraker et al., 2008). In vernal pools within 50 m of roads, Karraker et al. (2008) reported declines in Ambystoma maculatum (Spotted Salamander) abundance thought to be caused by increased salinity from road salt. Nonetheless, Jones et al. (2015) found no differences in the toxicity of road salt versus NaCl to Rana clamitans (Green Frog) or E. bislineata. They also found no mortality for E. bislineata from conductivity until concentrations reached ~10,000 us/cm (Jones et al., 2015). Additionally, Izzo (2013) found that road salt had no lethal effect on D. fuscus or E. bislineata until chloride concentrations exceeded 5,000 mg/l (~9,000 us/cm). The conductivity range for this study was 13-1660 us/cm. This study did not find conductivity (in the Chemistry Model) to have a strong relationship with salamander occupancy. The Chemistry Model only had weak empirical support for one species, D. ochrophaeus, with a Δ AIC of 3.53.
Eurycea spp. occupancy results showed that occupancy was negatively associated with the percent mining in the whole HUC-12 watershed, whereas the percent forested in the HUC-12 watershed was positively correlated with Eurycea spp. occupancy. Additionally, microhabitat models for Eurycea spp. did not have any empirical support. These results also found that other than the Recent Mining HUC-12 Model, none of the other potential mining-related parameters had empirical support (e.g. the Chemistry Model, the Valley Fill Model, and the Surface Mining Model) for Eurycea spp. In the Piedmont of North Carolina, Willson and Dorcas (2003) found that upland watershed disturbances (> 20% of watershed), such as agriculture and residential areas, caused declines in Eurycea (larval and adult) abundances, but that riparian buffer size and quality was not correlated to relative abundance. Similarly, Miller et al. (2007) found in the Piedmont of North Carolina that larval E. cirrigera abundances were negatively associated impervious surfaces in the watershed from residential and urban areas, however, abundances were not influenced by 50 m forested riparian buffers.
This difference in response of Eurycea spp. and Desmognathus spp. to microhabitat and landscape-level land use may be a function of movement and habitat selection. Adult Eurycea spp. are more terrestrial than the Desmognathus species, and E. b. cirrigera have been shown to migrate over 100 m away from the stream, whereas D. fuscus are largely sedentary having small home ranges of 1.4 to 3 m2 (Barthalmus and Bellis, 1972; Ashton, 1975; MacCulloch and Bider, 1975). Larval E. bislineata have been shown to move in stream. Small first-year larval tend to drift downstream, whereas larger second-year larval will move both upstream and downstream equally (Bruce, 1986; Petranka, 1998). The ability of E. bislineata to move long-distance may also explain why recent mining in the HUC-12 watershed had more of an influence on Eurycea spp. occupancy than recent mining in the subwatershed.
Predation and competition may also contribute to differences in results between Eurycea spp. and Desmognathus spp. Predation may have both direct mortality and indirect behavioral effects on stream salamanders, particularly on Eurycea spp., the smallest stream salamander regionally with the most potential aquatic predators. For example, Barr and Babbitt (2002) found that densities of E. b. bislineata were dramatically lower (9.4/m2 as compared to 54/m2) when the predatory Salvelinus fontinalis (Brook Trout) was present. However, Eurycea spp. may be more susceptible to predation from other stream salamanders than from fish predation. A stomach content analysis showed that up to 50% of G. porphyriticus, and 18% of D. quadramaculatus (Black-bellied Salamander) stomachs contained larval E. b. cirrigera (Bruce 1972). Predation from other stream salamanders may not only decrease survival rates, but may also influence microhabitat selection and behavior. Without predators, E. b. wilderae (Blue Ridge Two-lined Salamander) were evenly distributed in pools and riffles, however, when D. quadramaculatus, a large predator species that prefers riffles, were present, E. b. wilderae were more abundant in pools than in riffles (Beachy, 1993). Keen (1982) found when D. monticola, a predator of D. fuscus, was present, activity levels of D. fuscus dropped significantly.
Others have also hypothesized that decreases in some salamander species abundances when predatory fish and/or salamander species are present may not come from predation but rather from competition (Hairston, 1980; Davic, 1983; Barr and Babbitt, 2002; Bruce, 2011). Small-bodied, generalist salamanders such as Eurycea spp. are opportunistic generalists with diets largely consisting of pollutant-tolerant benthic macro-invertebrates such as Chironomids (Burton, 1976; Petranka, 1984; Muenz et al., 2008; Barrett et al., 2012). Eurycea spp. is perhaps better able to tolerate poor water quality and riparian habitat conditions where prey items (Chironomids) are often readily available. These disturbed conditions not only provide a plentiful food source for Eurycea spp., but may also provide a competitive release from larger salamander such as Desmognathus spp. and G. porphyriticus (Barrett et al., 2012; Surasinghe and Baldwin, 2015). However, the presence of Eurycea spp. in disturbed areas may also be influenced by predatory release (Ransom and Jaeger, 2006). More research that limits natural variability is needed to examine interspecific competition and predation among salamander species.
Overall, this research found effects of mining on stream salamanders to be equivocal; however, this is not to imply that mining does not affect stream salamanders. Although, the top model for Eurycea spp. was the HUC-12 Recent Mining Model, none of the species in this study had strong empirical support for many of the other mining-related parameters such as the percent active or past mining in the subwatershed, area of valley fill, conductivity, and the number of settling ponds. Past studies have found negative correlations between stream salamanders and coal mining (Hamilton, 2002; Wood and Williams, 2013a, b; Muncy et al. 2014). Still, these studies all examined the effects of mining versus reference conditions, and the resulting decreases in salamander assemblages may be a reflection of general riparian disturbance and not factors particularly unique to coal mining. The research helped differentiate effects of coal mining and general riparian disturbance on stream salamanders by using a gradient of land uses and disturbances. This gradient of sites showed poor riparian quality lead to declines in Desmognathus spp. abundance regardless of the type of large-scale land use associated with that riparian disturbance.
Although, stream salamander recovery from coal mining disturbance has not specifically been examined in this study, these results suggest that Desmognathus spp. occupancy may improve as riparian habitat quality increases post-mining. Proper mine land reclamation can decrease the amount of time needed for recovery. Currently, most mine land reclamation in central Appalachia has two terminal land uses: reforestation or grass-shrubland. Reforestation techniques such as those recommended by Appalachian Regional Reforestation Initiative (ARRI) suggest planting high-value hardwood trees on reclaimed mine land in order to produce commercially valuable crop trees along with quick-growing early successional trees for soil stability and initial wildlife value. Currently, the ARRI protocol does not have different management plans for riparian areas and upland areas. Additionally, current regulatory reclamation standards for grassland-shrubland land uses do not require reforestation of riparian areas. Regardless of target land use, in order to promote stream salamander recovery following mining, specific riparian reclamation guidelines are warranted. For example, quick-growing early successional tree species such as Betula lenta (Black Birch), Liriodendron tulipifera (Yellow Poplar), Salix nigra (Black Willow), Platanus occidentalis (American Sycamore), Populus deltoids (Eastern Cottonwood) and Pinus strobus (White Pine) would quickly establish a canopy cover, stabilize the soil, and impede invasive plant species (Davis et al., 2012) for more extensive list of suitable tree species adapted for moist/wet sites. Because, timber harvest within a riparian zone has been shown to cause changes in benthic macroinvertebrate, stream salamander, and fish densities and community structure (Jones et al., 1999; Rios and Bailey, 2006; Maigret et al., 2014), emphasis on riparian tree species prioritization could be based on those with soil retention/development and/or wildlife value instead of crop tree value. By developing a differential reforestation management plans for upland and riparian areas, a closer approximation of a native Appalachian forest may be achieved more quickly, and in turn provide high-quality refuges for stream salamanders and other biotic components of these aquatic ecosystems.
CONFLICT OF INTERESTS
The authors have not declare any conflict of interest.
REFERENCES
|
Anderson JT, Ward RL, Petty JT, Kite JS, Strager MP (2014). Culvert effects on stream and stream-side salamander habitats. Int. J. Environ. Sci. Dev. 5:274-281. |
|
|
Ash AN. (1988). Disappearance of salamanders from clearcut plots. J. Elisha Mitchell Sci. Soc. 104:116-122. |
|
|
Ashton RE, Jr. (1975). A study of the movement, home range, and winter behavior of Desmognathus fuscus. J. Herpetol. 9:85-91. |
|
|
Barr GE, Babbitt KJ. (2002). Effects of biotic and abiotic factors on the distribution and abundance of larval two-lined salamanders (Eurycea bislineata) across spatial scales. Oecologia 133:176-185. |
|
|
Barrett K, Samoray ST, Helms BS, Guyer C (2012). Southern two-lined salamander diets in urban and forested streams in western Georgia. Southeastern Naturalist 11:287-296. |
|
|
Barthalmus GT, Bellis ED (1972). Home range, homing, and the homing mechanism of the salamander, Desmognathus fuscus. Copeia pp. 632-642. |
|
|
Beachy CK (1993). Guild structure in streamside salamander communities: A test for interactions among larval Plethodontid salamanders. J. Herpetol. 27:465-468. |
|
|
Bruce RC. (1986). Upstream and downstream movements of Eurycea bislineata and other salamanders in a southern Appalachian stream. Herpetologica 42:149-155. |
|
|
Bruce RC (1972). Variation in the life cycle of the salamander Gyrinophilus porphyriticus. Herpetologica 28:230-245. |
|
|
Bruce RC (2011). Community assembly in the salamander genus Desmognathus. Herpetol.Monogr. 25:1-24. |
|
|
Burnham KP, Anderson DR. (2002). Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach. Springer-Verlag, USA. |
|
|
Burton TM (1976). An analysis of the feeding ecology of the salamanders of the Hubbard Brook Experimental Forest, New Hampshire. J. Herpetol. 10:187-204. |
|
|
Bury RB, Corn PS. (1989). Logging in western Oregon: Responses of headwater habitats and stream amphibians. For. Ecol. Manag. 29:39-57. |
|
|
Crawford JA, Semlitsch RD. (2008). Abiotic factors influencing abundance and microhabitat use of stream salamanders in southern Appalachian forests. For. Ecol. Manag. 255(5):1841-1847. |
|
|
Davic RD (1983). An investigation of salamander guild predation in a North Carolina stream: an experimental approach. Ph.D. Dissertation, Kent State University, Kent, Ohio. |
|
|
Davic RD, Welsh HH. (2004). On the ecological roles of salamanders. Annu. Rev. Ecol. Evol. System. 35:405-434. |
|
|
Davis V, Burger JA, Rathfon R, Zipper CE, Miller CR (2012). Selecting tree species for reforestation for Appalachian mined land. The Appalachian Regional Reforestation Initiative: Forest Reclamation Advisory, Forest Reclamation Advisory No. 9. |
|
|
deMaynadier PG, Hunter ML Jr. (1995). The relationship between forest management and amphibian ecology: A review of North American literature. Environ. Rev. 3:230-261. |
|
|
deMaynadier PG, Hunter ML Jr. (2000). Road effects on amphibian movements in a forested landscape. Natural Areas J. 20:56-65. |
|
|
Fisher SG, Grimm NB, Marti E, Holmes RM, Jones JB (1998). Material spiraling in stream corridors: a telescoping ecosystem model. Ecosystems 1:19-34. |
|
|
Ford WM, Chapman BR, Menzel MA, Odom RH (2002). Stand age and habitat influences on salamanders in Appalachian cove hardwood forests. For. Ecol. Manag. 155:131-141. |
|
|
Fowler DK, Hill DM, Fowler LJ (1985). Colonization of coal surface mine sediment ponds in southern Appalachia by aquatic organisms and breeding amphibians in Wetlands and Water Management of Mined Lands Proceedings (Brooks RP, Samuel DE, Hill JB eds.). The Pennsylvania State University, University Park. pp. 261-285. |
|
|
Freda J (1986). The influence of acidic pond water on amphibians: A review. Water, Air, Soil Pollution 30:439-450. |
|
|
Hairston NG (1980). Species packing in the salamander genus Desmognathus: What are the interspecific interactions involved? Am. Nat. 115:354-366. |
|
|
Hairston NG, Wiley RH (1993). No decline in salamander populations: A twenty-year study in the Southern Appalachians. Brimleyana 18:59-64. |
|
|
Hamilton MS (2002). Effects of developmental activities on streamside salamander communities in Boone County, West Virginia. Master Thesis, Marshall University, Huntington, West Virginia. |
|
|
Harper CA, Guynn DC (1999). Factors affecting salamander density and distribution within four forest types in the southern Appalachian Mountains. For. Ecol. Manage. 114:245-252. |
|
|
Herricks EE, Cairns J (1974). Rehabilitation of streams receiving acid mine drainage. V.P.I., Water Resources Research Center Bulletin 66:1-184. |
|
|
Izzo GD (2013). Investigating the toxicity of road salt to stream salamanders in urban watersheds: The role of stormwater management. Master Thesis, Towson University, Towson, Maryland. |
|
|
Jansen KP, Oakes CM, Colyer FD (2004). Success of reclamation practices to support amphibian populations. Proceedings of the National Meeting of the American Society of Mining and Reclamation. |
|
|
Jones B, JW Snodgrass Ownby DR (2015). Relative toxicity of NaCl and road de-icing salt to developing amphibians. Copeia 2015:72-77. |
|
|
Jones EBD, III, Helfman GS, Harper JO, Bolstad PV (1999). Effects of riparian forest removal on fish assemblages in southern Appalachian streams. Conserv. Biol. 13:1454-1465. |
|
|
Karraker NE, Gibbs JP, Vonesh JR. (2008). Impacts of road deicing salt on the demography of vernal pool breeding amphibians. Ecol. Appl. 18:724-734. |
|
|
Keen WH (1982). Habitat selection and interspecific competition in two species of plethodontid salamanders. Ecology 63:94-102. |
|
|
Kroll AJ, MacCracken TC, McBride J, Light P, Peterson Bach J (2010). Basin-scale JG surveys of stream-associated amphibians in intensively managed forests. J. Wildl. Manag.74:1580-1585. |
|
|
Lacki MJ, Hummer JW, Webster HJ (1992). Mine-drainage treatment wetland as habitat for herptofaunal wildlife. Environ. Manage. 16:513-520. |
|
|
Lemke D, Schweitzer CJ, Tazisong IA, Wang Y, Brown JA (2012). Invasion of a mined landscape: What habitat characteristics are influencing the occurrence of invasive plants? Int. J. Mining Reclam. Environ. 27(4):275-293. |
|
|
MacCulloch RD, Bider JR. (1975). Phenology, migrations, circadian rhythm and the effect of precipitation on the activity of E. b. bislineata in Quebec. Herpetologica 31:433-439. |
|
|
MacKenzie DI (2006). Modeling the probability of resource use: The effect of, and dealing with, detecting a species imperfectly. J. Wildl. Manag. 70:367-374. |
|
|
MacKenzie DI, Nichols JD, Royle JA, Pollock KP, Bailey LL, Hines JE (2006). Program PRESENCE. Occupancy estimation and modeling:inferring patterns and dynamics of species occurrence. Academic Press, USA. |
|
|
Maigret TA, Cox JJ, Schneider DR, Barton CD, Price SJ, Larkin JL (2014). Effects of timber harvest within streamside management zones on salamander populations in ephemeral streams southeastern Kentucky. For. Ecol. Manage. 324:46-51. |
|
|
Maxwell AE, Strager MP, Warner TA, Zegre NP, Yuill CB (2014). Comparison of NAIP orthophotography and RapidEye satellite imagery for mapping of mining and mine reclamation. GISci. Remote Sensin 51:301-320. |
|
|
Middlekoop JT, Watts T, Schorr M (1998). Acid mine drainage and its effects on physicochemical conditions and salamander populations in a Cumberland Plateau stream. J. Tennessee Acad. Sci. 73:36. |
|
|
Miller JE, Hess, GR, Moorman CE (2007). Southern two-lined salamanders in urbanizing watersheds. Urban Ecosystems 10:73-85. |
|
|
Minear RA, Tschantz BA (1976). The effects of coal surface mining on the water quality of mountain drainage basin streams. J. Water Pollut. Control Fed. 48:2549-2569. |
|
|
Moseley KR, Ford WM, Edwards JW, Schuler TM (2008). Long-term partial cutting impacts on Desmognathus salamander abundance in West Virginia headwater streams.For. Ecol. Manag. 254:300-307. |
|
|
Muenz TK, Golladay SW, Smith LL, Vellidis G (2008). Diet and abundance of southern two-lined salamander larvae (Eurycea cirrigera) in streams within an agricultural landscape, southwest Georgia. Southeastern Nat. 7:691-704. |
|
|
Muncy BL, Price SJ, Bonner SJ, Barton CD (2014). Mountaintop removal mining reduces stream salamander occupancy and richness in southeastern Kentucky, USA. Biological Conservation 180:115-121. |
|
|
Noble CV, Berkowitz JF, Spence J (2010). Operational draft regional guidebook for the functional assessment of high-gradient ephemeral and intermittent headwater streams in western West Virginia and eastern Kentucky. ERDC/EL TR-10-11. US Army of Corps of Engineers, Vicksburg, Mississippi. |
|
|
OHEPA (2012). Field evaluation manual for Ohio's primary headwater habitat streams. Version 3.0. Ohio Environmental Protection Agency, Division of Surface Water, Columbus, Ohio. |
|
|
Pauley TK, Mitchell JC, Buech RR, Moriarty J (2000). Ecology and management of riparian habitats for amphibians and reptiles. In. Riparian Management in Forests of the Continental Eastern United States (Verry ES, Hornbeck CA, Dolloff CA, eds.). CRC Press, USA. pp. 169-191. |
|
|
Pearl CA, Adams MJ, Leuthold N, Bury RB. (2005). Amphibian occurrence and aquatic invaders in a changing landscape: Implications for wetland mitigation in the Willamette Valley, Oregon, USA. Wetlands 25:76-88. |
|
|
Peterman WE, Semlitsch RD. (2009). Efficacy of riparian buffers in mitigating local population declines and the effects of even-aged timber harvest on larval salamanders. For. Ecol. Manage. 257:80-14. |
|
|
Petranka JW (1984). Ontogeny of the diet and feeding behavior of Eurycea bislineata larvae. J. Herpetol.18:48-55. |
|
|
Petranka JW (1998). Salamanders of the United States and Canada. Smithsonian Institute Press, USA. |
|
|
Petranka JW, Eldridge ME, Haley KE (1993). Effects of timbering harvesting on southern Appalachian salamanders. Conserv. Biol. 7:363-370. |
|
|
Resetarits WJ (1997). Differences in an ensemble of streamside salamanders (Plethodontidae) above and below a barrier to brook trout. Amphibia-Reptilia 18:15-25. |
|
|
Rios SL, Bailey RC. (2006). Relationship between riparian vegetation and stream benthic communities at three spatial scales. Hydrobiologia 553:153-160. |
|
|
Russell KR, Mabee TJ, Cole MB. (2004). Distribution and habitat of Columbia torrent salamanders at multiple spatial scales in managed forests of northwestern Oregon. J. Wildl. Manag. 68:405-417. |
|
|
Schorr MS, Dyson MC, Nelson CH, VanHorn GS, Collins DE, Richards SM (2013). Effects of stream acidification on lotic salamander assemblages in a coal-mined watershed in the Cumberland Plateau. J. Freshwater Ecol. 28:339-353. |
|
|
Southerland MT, Jung RE, Baxter DP, Chellman IC, Mercurio G, Vølstad JH (2004). Stream salamanders as indicators of stream quality in Maryland, USA. Appl. Herpetol. 2:23-46. |
|
|
Surasinghe TD, Baldwin RF (2014). Ghost of land-use past in the context of current land cover: evidence from salamander communities in streams of Blue Ridge and Piedmont ecoregions. Canadian J. Zool. 92:527-536. |
|
|
Surasinghe TD, Baldwin RF (2015). Importance of riparian forest buffers in conservation of stream biodiversity: Responses to land uses by stream-associated salamanders across two southeastern temperate ecoregions. J. Herpetol. 49:83-94. |
|
|
Sweeten SE (2015). Effects of land use and microhabitat on stream salamander occupancy and abundance in the southwest Virginia coalfields. PhD Dissertation, Virginia Polytechnic Institute and State University, Blacksburg. |
|
|
Walz A (2002). The study of associations between Maryland's amphibian species and habitat parameters at local and landscape scales. PhD Dissertation, University of Maryland, College Park. |
|
|
Ward RL, Anderson JT, Petty JT (2008). Effects of road crossings on stream and streamside salamanders. J. Wildlife Manag. 72:760-771. |
|
|
Welsh HH, Droege S. (2001). A case for using plethodontid salamanders for monitoring biodiversity and ecosystem integrity of North American forests. Conserv. Biol. 15:558-569. |
|
|
Welsh HH, Ollivier LM (1998). Stream amphibians as indicators of ecosystem stress: A case study from California's redwoods. Ecol. Appl. 8:1118-1132. |
|
|
Welsh HH, Hodgson GR, Lind AJ (2005). Ecogeography of herpetofauna of a northern California watershed: Linking species patterns to landscape processes. Ecography 28:521-536. |
|
|
Willson JD, Dorcas ME (2003). Effects of habitat disturbance on stream salamanders: Implications for buffer zones and watershed management. Conserv. Biol. 17:763-771. |
|
|
Wood PB, Williams JM (2013a). Impact of valley fills on streamside salamanders in southern West Virginia. J. Herpetol. 47:119125. |
|
|
Wood PB, Williams JM (2013b). Terrestrial salamander abundance on reclaimed mountaintop removal mines. Wildl. Soc. Bullet. 37:815-823. |
|
|
Woodward SL, Hoffman RL (1991). The nature of Virginia. Virginia's Endangered Species (K. Terwillinger, ed.). McDonald and Woodward Publishing, USA. pp. 23-48. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


