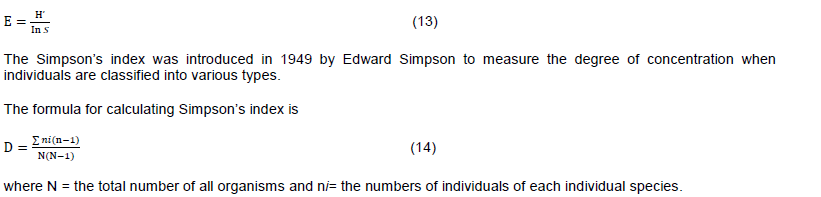ABSTRACT
The Kimbi Fungom National Park is reported to be relatively less diverse compared to other national parks of Cameroon. This might be due to the configuration of the landscape and habitat types. The main objective of this study is to determine the species composition, structure and diversity in the heterogeneous landscape of the Kimbi Fungom National Park defined by different habitat types. The park was divided into 20 blocks and 10 blocks were randomly selected for this flora survey. In each block, a 1 km line transect was established with 5 quadrates of 20 × 20 m. These transects were laid to cut across four major vegetation types (lowland rainforest, gallery forest, woodland and grassland savanna). A total of 2831 stems ³1 cm circumference were identified and measured. This belongs to 222 species, in 54 families. Fabaceae, Rubiaceae and Euphorbiaceae were found to be the most dominant plant families. Tree density and basal area were lowest in the grassland while it was highest in the lowland forest. Species composition and diversity varied with vegetation type and landscape configuration. The lowland and gallery forest were characterized by high diversity and similar species composition shown by the Shannon diversity index of 3.8 and forest 3.75, respectively. Three species from our survey were known to be endemic to Cameroon (Allophyllus bullatus, Cleistopholis staudtii and Magnistipula butayei) while 6 species were known to be vulnerable; Afzelia africana, Afzelia bipindensis, Allophyllus bullatus, Entandrophragma angolense, Hallea stipulosa, and Quassia sanguine.
Key words: Flora composition, vegetation structure, species diversity, heterogeneous landscape.
Vegetation is formed by plant species growing as the result of a long development process consistent with climate and the places they inhabit, and which constitutes a fundamental part of ecosystems (Velázquez and Romero, 1999; FGDC, 2005). Ecological diversity is the degree at which life forms varies within the context of a particular ecosystem, biome, or entire planet (Uno et al., 2001). This encompasses all species of plants, animals and microorganisms, the ecosystem, and ecological processes of which they belong. It is a general term for the extent of nature’s variety, including both number and frequency of ecosystems, species, or genes in a given assemblage. Wilson (1992) defines biodiversity as the variety of organisms considered at all levels from genetic variants belonging to the same species through arrays of species to arrays of genera, families, and still highertaxonomic levels.
Studies of the main tropical forest ecosystems have shown that African rainforests have relatively poor diversity compared to the highest diversity regions of Asia and the Americas (Parmentier et al., 2007). However, based on this overall pattern of diversity, current understanding of the local-scale community-assembly mechanisms for tropical African tree communities is very limited and complicated by previous sampling designs. For instances, inventories based on 1-ha plots spread across a wide area capture fewer than half of the local species, with many represented only by a single individual. In addition, most of these inventories focus on large trees with dbh 10 cm (Hall and Swaine, 1981; Hardy and Sonke, 2004) and in some cases only include selected taxa (Hall et al., 2004). These small plots limit the identification of habitats at scales that could provide meaningful inferences on plant populations and also preclude comparisons of degrees of habitat specificity with other tropical forests thus having the misconception of poor diversity of the African rainforest.
Besides other African countries, Cameroon is one of the most diverse countries in terms of plant, with over 7,850 plant species (Onana 2011). From these species, 815 species are endangered (Onana and Cheek 2011). The Cameroon heterogeneous landscape presents different vegetation types among which are the Biafran forest with high rainfall, the Congolese forest, and the semi-deciduous forest with low rainfall (Letouzey, 1985). Thus, Cameroon encompasses an intricate mosaic of diverse habitats with moist tropical forest dominating the south and south-east and covering 54% of the country, mountain forest and savannah in the highlands and sub-Sahelian savannah and near desert in the far north (Sunderland et al., 2003). The vegetation of Cameroon ranges from lowland evergreen rainforest, semi-deciduous, deciduous, savannah woodland, and savannah grassland forest, at different altitudinal gradient of lowland to sub-montane, alpine and montane forest (Letouzey, 1985; Achoundong, 2007). It forms part of the Guineo-Congolian region of endemism (White, 1983). Following these features, the forest of the cross-border region of Cameroon and Nigeria are highly diverse with a high degree of endemism (Davis et al., 1994). Further studies conducted by Barthlott et al. (1996) ranked Cameroon among the top countries in tropical Africa for plant species diversity per degree square. Similar studies equally confirmed the high diversity of endemism of plant species, as found in the 50 ha plot in central Korup National Park, Cameroon with close to 500 tree species (Thomas et al., 2003) and over 250 liana species. Most of this high diversity is usually preserved in protected areas through gazettement. Though the flora is highly studied, new species are recorded every year (Lachenaud et al., 2013).
Landscape configuration significantly influence local conditions, especially soil processes and hydrology, that
may affect the processes of growth, mortality, and recruitment (Daws et al., 2002; Miyamoto et al., 2003).
Variation in demographic performance among habitats can translate into species’ associations with their preferred habitats (Harms et al., 2001; Russo et al., 2008; Yamada et al., 2007). Thus, resolving the role of habitat partitioning in the maintenance of high species diversity in ecosystems will depend in part on detecting species–habitat associations (Webb and Peart, 2000; Potts et al., 2004). Differences among species in their habitat associations, coupled with habitat heterogeneity, will contribute to the maintenance of high diversity by allowing species to coexist by specializing on different habitats.
Diversity studies carried out in Cameroon have covered many parts of the country but left out certain regions despite their richness in plant diversity (Mbolo, 2002; Ndenecho, 2005). An example of such a region is the newly created Kimbi Fungom National Park (K-FNP) in the North West Region of Cameroon characterized by a hilly topography with a rich diversity of flora and fauna. This area is part of the Cameroonian Highlands ecoregion which encompasses the mountains and highland areas of the border region between Nigeria and Cameroon (Stuart, 1986; Ndenecho, 2009). The area falls within the Afromontane archipelago-like regional centre of endemism that spans the entire African continent (Ndenecho, 2009). The forests in the area are refugia in montane and sub-montane environments. The montane forests are of great ecological significance and contain several endangered species of plants and animals (Alpert, 1993; Ngwabuh, 2002). This region holds some of the globally threatened and endemic species such as the critically endangered Nigeria-Cameroon chimpanzee (Pan troglodytes Ellioti) and the pangolin (Smutsia gigantea). It also holds some tree species with high conservation values. This rich ecoregion located in the Western Highlands of Cameroon has been under serious human influence for over three decades. Conservation efforts have tended to emphasize the protection of biodiversity and to improve local livelihoods. Based on this, some questions were asked such as; which is the most diverse vegetation type in the park? What is the structure of the different habitats? What is the similarity between the vegetation types of the park?
From the aforementioned questions, this study’s main objective was to describe the flora K-FNP. Specifically, this study sets out to;
i) Assess the species composition of the park;
ii) Evaluate the structure of the different vegetation types; and
iii) To ascertain species diversity in park.
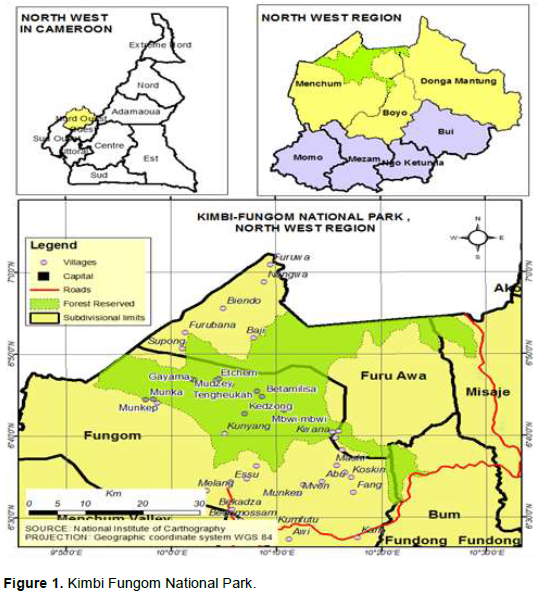
The Kimbi-Fungom National Park is located between latitude 6.5- 6.9° N and longitude 9.8-10.5° E in the North West Region of Cameroon covering a total land surface of 95,380 ha. This national park is located in three divisions of the North West Region of Cameroon cutting across four sub divisions. These divisions are Menchum, Boyo and Dongo Mantung and found in the respective sub divisions of Fungom and Fru Awa, Misaje and Bum. The Kimbi-Fungom National Park (K-FNP) is a newly created national park and the only national park in the region (Figure 1).
Area of the park?
This region experiences two seasons; a long rainy season from mid March to mid November and a short dry season from mid November to mid March. The wettest months are July, August and September and the driest months being January and February. Hawkins and Brunt (1995) described the climate as a “sub-montane cool and misty climate” with an annual mean maximum temperature of 20 to 22°C and mean minimum of 13 to 14°C. Annual rainfall varies between 1780 and 2290 mm with most of the rainfall occurring between July and September. A dry season occurs from mid - October to mid-March (Tata, 2015).
Geographically, the Kimbi-Fungom National Park has a heterogeneous landscape. The Fungom area lies east of Weh-Esu and South of Esu to Kung and Fang reaching a height of 1524 m. This area is made-up of woody savanna with hills running from Weh to Kuk. It is characterized by a rugged terrain from steep rolling hills into extensive flat valley at lower altitude. The Munkep-Gayama axis is an extensive valley about 6 km wide in the Munkep area to over 10 km in Gayama zone. It is in the midst of chains of some steep rocky hills which are almost impassable. The lowlands take another orientation from Munkep at a place called ‘Last Town’ towards the eastern forest. The valley starts behind the eastern forests where it extends for more than 15 km wide to over 30 km long. This extensive eastward valley is fertilized by alluvial silts from the Southern near Escarpment chain of long hills that stretched from the West towards the East in the Fungom Reserve. This relief has greatly influenced the vegetation types and distribution within the park. The Munkep Gayama axis lies on an extensive valley between chains of two hills. These valleys gradually protrude into near long rolling steep hills which are characterized by the woody vegetation. The hills are sandwich by galley forests which form the basis for the numerous tributaries in the park.
It is drained by a wide range of rivers and streams, notably the Kimbi, the Katsina Ala, and the Kendassamen Rivers, along with significant streams that include: the Batum, Akum, Bissaula, Kenda, Yemene and Imia. These streams flow into the bigger ones that eventually flow through the Katsina Ala River and finally enter the River Benue (Tata, 2015).
The soil types in this area include; acrisols, andosols (Black soils of volcanic landscapes), ferrasols (red and yellow tropical soils with a high content of sequioxide), leptosol (shallow soil) and nitisols (deep, well-drained, red, tropical with a clayey subsurface) (Birdlife, 2010).
The vegetation is principally lowland tropical rainforest at the Fungom low altitude area of the park and gradually progress into tropical deciduous forest to savanna and the grassland savanna. It has many tree species of economic values.
Data collection
Data collection on the flora took place in two different times; November to December 2017 for the dry season and May to July 2018 for the wet season. The park was divided into 20 equal blocks of 7 × 7 km each (4900 ha) with ten blocks randomly selected for this study. In each block, a 1-km transect as described by Tchouto (1999) and Buckland et al. (2007) was established. In each transect, 5 quadrates of 20 × 20 m were established at interval of 200 m (this gives a total of 0.2 ha of land sampled per block). The quadrates along transects were placed in alternate manner (that is, if quadrate one is on the left of the transect, quadrate two is placed at the right). A total of fifty 20 × 20 quadrates were sampled giving 2 ha of total land covered (representing 0.0002 ha of total park). These transects were laid to cut across four major vegetation types (lowland forest, gallery forest, woody savanna and grassland) at different elevations. Two other vegetation types were identified; the swampy forest where raffia forest grows and the secondary forest, which was fallowed farmlands. We sampled all plants ≥1 cm stem diameter in all quadrates. In each quadrate, four subplots of 1 × 1 m were placed at the corners to sample understory and herbs (Oosting, 1956; Condit, 2008).
Identification
of plants was done in the field using various methods. The trees were identified using a combination of characters such as the general form of the tree (butresses, roots systems, bark texture; slash colour, smell and exudates, leaf type and shape) as well as the flowers, and fruits of the trees. In each transect, records of all species of vascular plants, excluding tree dwelling epiphytes were taken. For trees that were unable to be identified, the leaves were collected and put in a plant press for the Limbe Botanic Garden herbarium. Tree structural data were collected and recorded in each quadrate using a datasheet file. Each live and dead tree within the plot was identified and was measured at 1.3 m dbh. The dbh of all trees were measured using the dbh metal tape. The height of trees were measured using the hypsometer (Vitax) and also by estimation (average estimates of all field researchers). Some trees generally posed a lot of difficulties in measuring the dbh at 1.3 m, due to the configuration of their buttresses, lianes and stems at 1.3 m. In such a situation, the dbh was measured at a distance either above or below 1.3 m. Field manuals, field books, text books all on plants were used to help in the identification of the plants/trees in the field.
At the start, each geographical coordinates were taken with the help of a GPS. All individual species were counted and recorded in each quadrate on a prepared data sheet by name for identified species and codes for unidentified species. Materials used here included; a portable field press, collecting bags, a machete, a sonny camera, a GPS, field note book, a 30 m measuring tape, a diameter tape and a rope. We used the recommended GPS (Global-Positioning-System) Garmin 60CSx, which are so accurate in the forest, inexpensive and works adequately under forest canopy (Condit, 2008).
Species composition
A total of 2831 individual trees identified with dbh of ≥1 cm were identified and measured. These identified species were found to be grouped into 54 families with content of 222 species. The species were distributed through four main vegetation types (lowland rainforest, gallery forest, woody savanna and grassland savanna) and secondary and swampy forest which were found in any of the four main vegetation types. These six vegetation types were identified based on species composition and structural characteristics. A total of 129 species were identified in the lowland forest, 124 in the gallery forest, 52 in the woody savanna, 56 in the secondary forest, 40 in the swampy forest, and 49 in the grassland savanna.
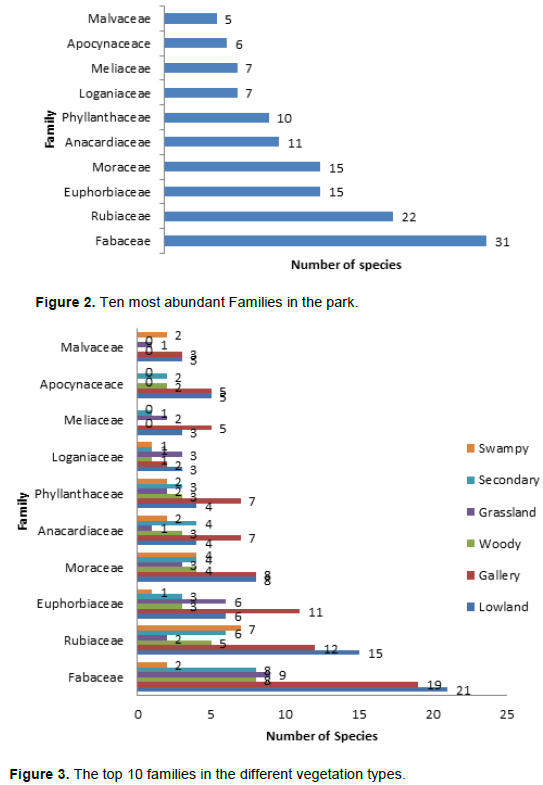
The families with the highest number of species in descending order for the entire study area was Fabaceae, Rubiaceae, Moraceae, Euphorbiaceae, Anacardiaceae Phyllanthaceae, Loganiaceae, Meliaceae, Apocynaceae and Melvaceae (Figure 2). These 10 families accounted for 47.51% of total species sampled and contributed 40.55% of total plant sampled in the study area. Two of these families, Fabaceae and Rubiaceae, were the most diverse families, which accounted for 24.23% of overall species richness and contributed 53 species that accounted for 23.98% of the total number of plant sampled. Fabaceae has 31 species, Rubiaceae 22, Euphorbiaceae and Moraceae both had 15 species and the least in the top 10 which is Malvaceae had 5 species (Figure 2).
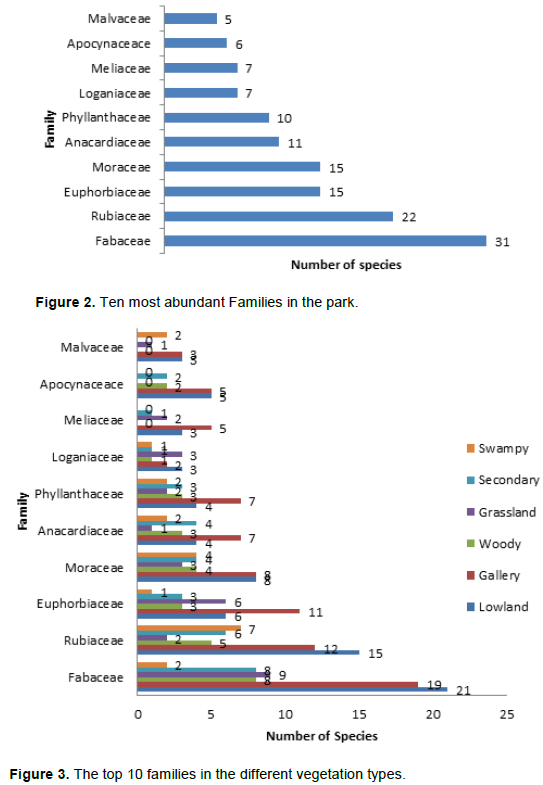
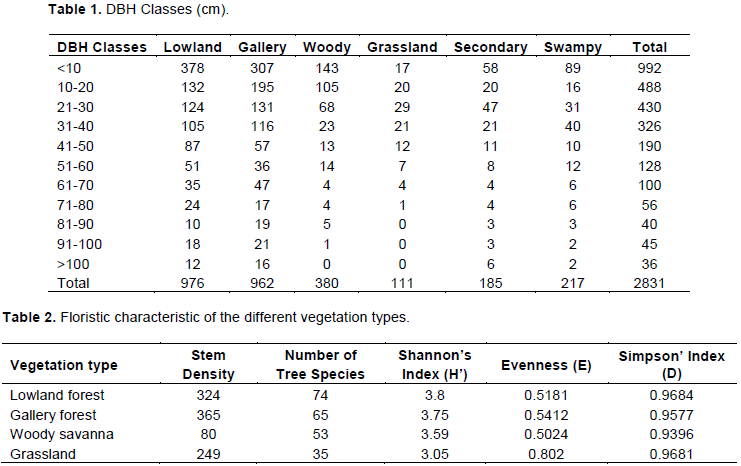
Going by vegetation type, Fabaceae, Rubiaceae and Euphorbiaceae still dominate lowland, gallery forests. Fabaceae had 21 species in the lowland forest, 19 in the gallery forest, 8 in the woody savanna, 9 in the grassland, 8 in the secondary forest and no species in the swampy forest. Rubiaceae on the other hand, had 15 species in the lowland forest, 12 in the gallery forest, 7 in the swamp, 6 in secondary forest, 5 in the woody savanna and 2 in the grassland. Figure 3 summarizes the distribution of the 10 most abundant families in the different vegetation types.
From the 222 species found, 34 (15.32%) were common only in the lowland tropical rainforest, 19 (8.56%) were common only to gallery forest, 7 (3.15%) were common to the woody savanna, 2 (0.9%) common to swampy forest, 2 (0.9%) to secondary forest and 7 (3.15%) only common to grassland savanna. Only one species (Albizia species) was common in all 6 vegetation types. Ten species were common in both lowland forest and gallery forest. The remaining 140 species (63.06) were common in 3 to 5 vegetation types.
Structure
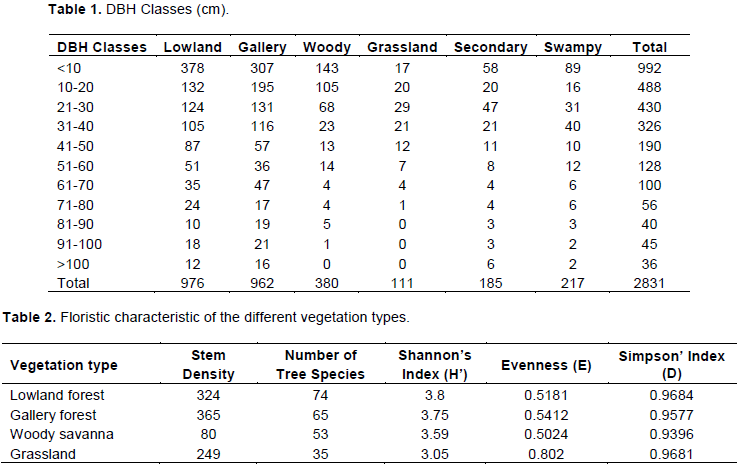
A total of 2831 stems with circumference of 1 cm from 222 species were recorded across 4 main vegetation types and 2 others (secondary and swampy forests) in the park. The distribution of stems by the circumference classes in the different vegetation types differ. Most individual plants (992) were concentrated in the first class of less than 9.9 cm. From this number, 378 (38.1%) were concentrated in the lowland forest, 307(30.9%) in the gallery forest, 148 (14.8%) in the woody savanna, 17 (1.7%) in the grassland, 58 (5.8%) in the secondary forest, and 89 (9%) in the swampy forest. The stems above the 10 cm class were concentrated in different classes; 13.5% were concentrated in 20-30 cm in lowland forest, 20.27% concentrated in the 10-20 cm class in the gallery forest, 28.62% concentrated in the 10-20 cm class in the woody savanna and 21.21% were concentrated in the 21-30 cm class in the grassland, 25.41% were concentrated the 21-30 cm class in the secondary forest and 18.43% were concentrated in the 31-40 cm class in the swampy forest. Only the lowland forest, gallery forest and the secondary forest had species in the last two classes of above 100 cm (Table 1).
The distribution of the number of stems differs among circumference classes. The variation was determined by the result of the variance which shows a significant difference (p < 0.001). The number of stems with big circumference from classes 90-100 and above 100 cm was very low in all parts of the National Park. Among the range of species, 36 stem had large dbh class of above 100 cm with seven stems having a remarkable large average circumference of 140 and beyond: Ceiba pentandra (200 cm), Pseudospondias microcarpa (200 cm), Albizia spp. (180 cm), Ricinodendron heudelotii (150 cm), Maranthes glabra (150 cm), and Erythrophleum ivorense (140 cm).
Species diversity
Comparison was only possible for lowland, gallery forest, grassland-savannah and woody-savannah. This is because they were well represented in the sample size than secondary and lowland forest. Based on this, the analysis was done using sample size of 0.2 ha. Plot size was considered to be 20 × 10 m whereas quadrate was considered to be 20 × 20 m; therefore, following our method we expect to have five quadrates in a modified plot sampling of 20 × 100 m. The total number of individuals (N) as well as species richness (S) was high in the lowland forest and the gallery forest than in the woody savanna and grassland. According to Margalef’s index of species richness, representing an intermediate mathematical measure between S/N and S, the lowland forest was the most diversified, followed by the gallery forest, grassland savanna, woody savanna in that order. Shannon’s measure of evenness did not differ significantly among different vegetation types (P<0.001). Shannon-Wiener’s information index, which combines species richness and evenness into a single value, indicated that the diversity of all vegetation types were
not very different, but the grassland was poor because its value is less than 3.5. Simpson index which is a measure of dominance (D) showed that the lowland forest remained very rich than other vegetation types. As D increases, diversity (in the sense of evenness) decreases (Table 2).
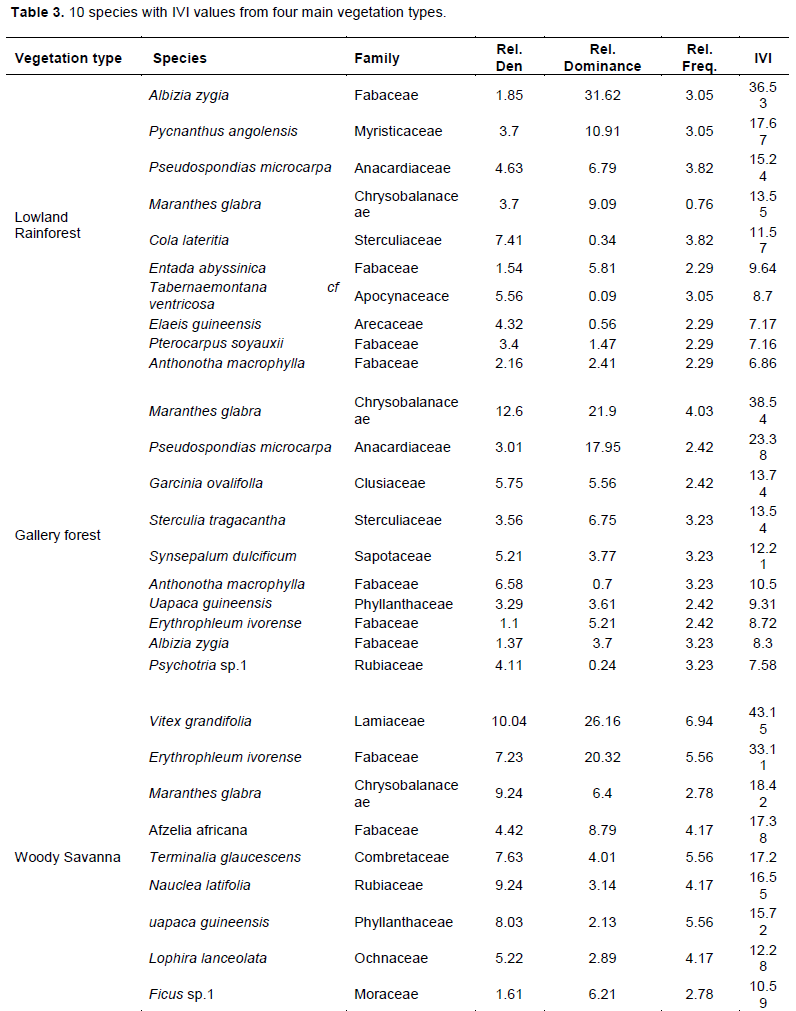
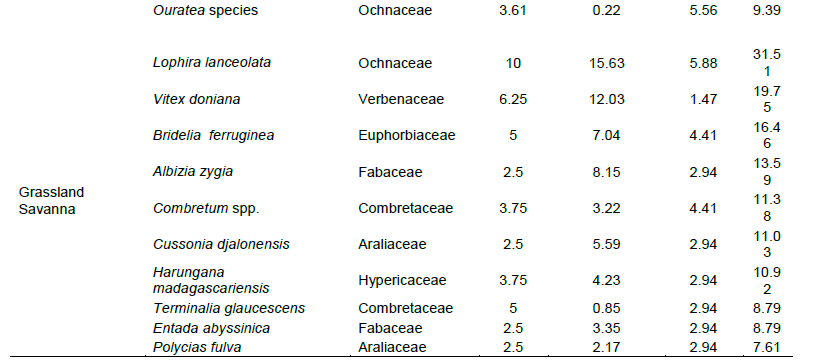
In accordance with vegetation type, Albizia zygia, Pycnanthus angolensis, P. microcarpa, M. glabra and Cola lateritia were found to dominate the lowland forest, while M. glabra, P. microcarpa, Garcinia ovalifolla, Sterculia tragacantha and Synsepalum dulcificum dominated the gallery forest. The woody savanna was dominated by Vitex grandifolia, E. ivorense, M. glabra, Afzelia africana, and Terminalia glaucescens while the Lophira lanceolata, Vitex doniana, Bridelia ferruginea, A. zygia and Combretum species. dominated the grassland savanna. This is shown by their IVI. This phytosociological analysis of assemblages from habitats showed that, for the first 10 species of each vegetation type grouped with highest IVI (importance value Index), the relative values of density, frequency and dominance, when expressed as IVI indexes, were numerically similar (Table 3).
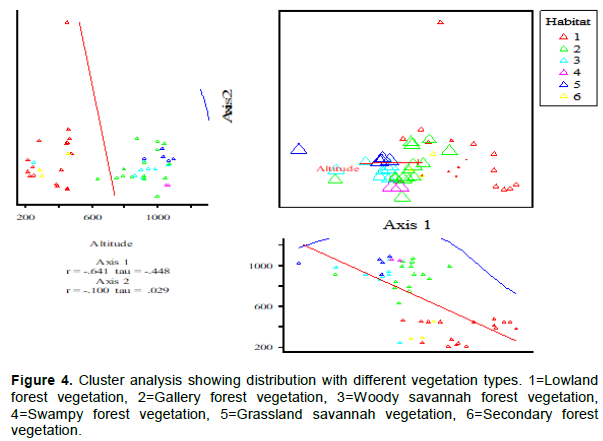
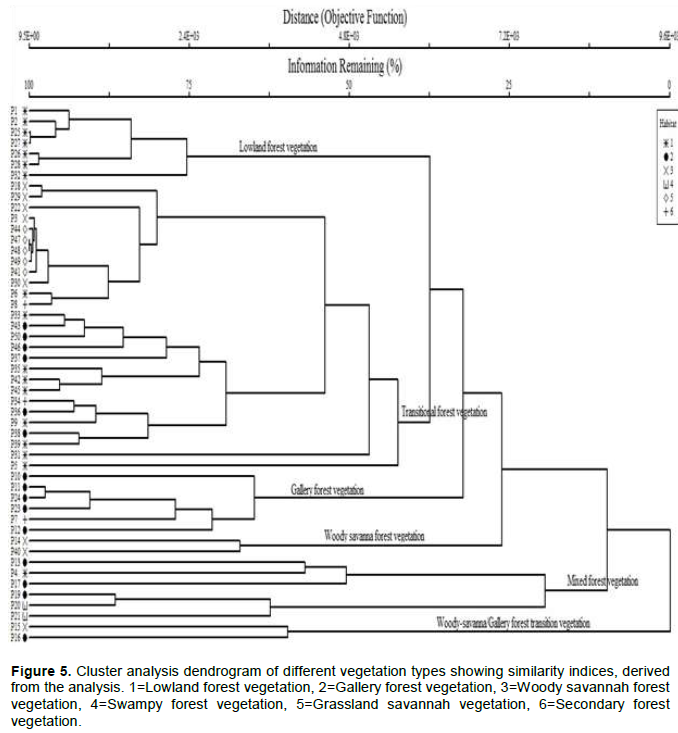
Cluster analysis indicated the presence of transitional and mixed forest amidst the normal four vegetation types. The vegetation types were heterogeneous but composed of some samples close to each other. Except for the lowland where we discovered an outlier with one outstanding quadrate (15) that has purely the characteristic of lowland forest, the lowland forest showed closeness in species composition with the secondary forest and to an extent with gallery forest and woody savanna. This is due to the fact that these vegetation types (secondary, woody and gallery) are also found in low altitudes and share many species which are found in the lowland forest. Most of the species are located below 600 m asl. The gallery forest is again more closed to woody savanna and is mostly located at altitude between 600 and 1000 m asl. Gallery forests were found to stretch from lowland into very highlands. The grassland represented by 5 showed closeness with the swampy forest and were found to be dominated at elevation of 1000 m and above. Some of the swampy forests observed in the park were located in highland areas where raffia forests dominate (Figure 4).
The floristic composition within these forest types were more or less variable, forming two additional vegetation types, with similarity percentages ranging from 30 to 57%, transitional forest that had a similarity value at the 25% limit. The lowland forest gallery forest and woody savanna were more similar with similarity percentage ranging from 50 to 75%. The transitional (lowland, secondary and gallery) forest was more similar to the mixed forest (woody, swamps and grassland) with similarity percentage ranging from 20 to 45%. These vegetation types are close to each other. The lowland is closer to gallery and woody savanna in terms of altitude. This is shown on the dendrogram (Figure 5).
Fabaceae, Rubiaceae, Euphorbiaceae. Moraceae, Anacardiaceae, and Phyllanthaceae were the most abundant families observed in the park across the different vegetation types. In this study, Fabaceae was the most dominant plant family with 31 species. This is in line with Sainge (2016) observation the park. This confirmed the idea that Fabaceae always fall among the three most dominant families in the world. Fabaceae is the most diverse plant family in the world with a wide distribution of sort, registering 770 genera and 19,500 sorts, and considered the third largest family of angiosperms in species numbers after Asteraceae and Orchidaceae in the global context (Beech et al., 2017; Azani et al., 2017). These taxa are always present whenever native floras are sampled in the Amazon (Matos and Amaral 1999). Fabaceae has been noted to always appear among the three most abundant families in the Amazon both in old and recent forest fragments, as well as various types of forests (Rankin-de-Merona et al., 1992; Santos et al., 2013). In studies using DBH ≥ 10 cm, Fabaceae is considered hyperdominant, where other families are not part of hyperdominant groups because most of their individuals are small. In addition, Fabaceae had the greatest species richness and it is commonly cited as the richest family in other Amazon forest types (Pitman et al., 2002; Oliveira et al., 2004; ter Steege et al., 2006). Myrtaceae and Fabaceae were considered by Miranda et al. (1993) as the richest families in forest areas within the Roraima savannas.
Rubiaceae was the second most abundant family in our study area with 22 species. Its abundance had also been recorded in other parts of Cameroon. For instance, Fonge et al. (2013), in their study of the diversity, distribution, and abundance of plants in Lewoh-Lebang in the Lebialem Highlands of Southwestern Cameroon, found Rubiaceae as the most dominant family and suggested that Rubiaceae could be the most dominant tree family in the Guinean equatorial forest. These findings were also recorded by Ndam et al. (2001) and Fonge et al. (2013) who reported that the Rubiaceae was the most dominant tree family in the Mount Cameroon region. Kenfack et al. (2007) also report Rubiaceae to be the most dominant tree family in the Korup National Park. The abundance of Rubiaceae in forest landscapes has also been reported for Assam and the North East Frontier Agency (present Arunachal Pradesh) where Chowdhury (2005) listed 150 species of Rubiaceae from the state of Assam which shows both the great range of this family and/or the capacity of its members to adapt to forested areas. The family Rubiaceae is widespread and occurs in all major regions of the world except Antarctica and shows maximum proliferation in low to mid altitude humid forests. The family comprises 13,143 species falling under 611 genera by which it is the fourth largest Angiosperm family (Davis et al., 2009).
According to the structure of species in the national park, the distribution of trees in circumference classes was uneven. We recorded 11 dbh classes in our study indicating that, structurally, lowland forest and gallery forests are probably mature, stable and highly likely to continue perpetuating their constituent species. The smallest diameter trees (1-9.99 cm dbh) were 994 (35.11%) abundant and those >10 cm were 1837 (64.5%) showing the ecological importance of small-trees in the structure, diversity and biomass to tropical forests (Memiaghe et al., 2016). This is slightly different from Sainge (2016) based on this percentages. In Sainge (2016) 17 ha study of the species structure of the park, 5152 (92.81%) trees with dbh ≥ 10 cm, and 399 (7.19%) trees with dbh 1-9.9 cm were recorded. Going by the different classes > 10 cm, the class 10-20 cm and 21-30 cm were more abundant than the other classes. These results were similar to those of Savadogo et al. (2007) in Tiogo Forest who demonstrated that at dbh of > 10 cm, a great number of stems were recorded from the circumference class 10 - 30 to 30 - 50 cm, indicating the high number of small trees and the higher contribution of shrub species which developed small circumference. This is partly due to rampant devastation of the forests basically, the suppression and the falling of big trees in addition to other factors that limited the sustainability of species (Peters, 1997). The record of average of circumference and basal area were low in the grassland in the K-FNP. This was due to overgrazing and bush fire which influenced biodiversity by reducing the number of stems desired, affected species diversity and their size (Moleele and Perkins, 1998; Boussim et al., 2009). The density of the desired species reduced due to the devastating effects and lack of reafforestation in the park, thus explaining the low rate of young plants renewal in the woody and grassland (Ntoupka, 1999).
Also, vegetation is said to be rich if it has a Shannon diversity value ≥ 3.5 (Kent and Coker, 1992). Whittaker (1970) stated that vegetation diversity is dependent on two characteristics of the vegetation. One is the number of species present, identified as S, which indicates species richness, whereas the other describes the distribution of the individuals in the population among the species, symbolized by N. In our study site, 3 out of four vegetation types had Shannon- Weaver diversity indices greater than 3.5 except the grassland which has a diversity index of 3.05. This shows that the different ecosystems are rich in species diversity. The lowland forest was the most diverse and also the S followed by the gallery forest. This could be due to the fact that the lowland forests were large enough and also receive more nutrients from the surrounding hill which support the growth of diverse plant species while most gallery forests were relatively undisturbed though anthropogenic factors such as agriculture and hunting are widespread in the park. Most gallery forests are very steep, and were neither good for farming or grazing; also, the abandonment of farming activities in some areas of the park as lands had been left to fallow for a very long time (Fonge, 2011). Although large plantations of cocoa, palms, maize, and millets were observed to be cultivated in the park, a high diversity index was still recorded for the different vegetation types, meaning that some species or families might have been erased due to these high anthropogenic factors. This high diversity index is in line with Sainge (2016) observations Shannon diversity index of 4.51 with a species richness of 86%. It was concluded that, the Kimbi Fungom Forest though poor in species compares to the wet Rumpi Hills Forest, but it has a high flora diversity and species richness in the North West Region.
For two or more areas to be considered alike, they must have at least 25% of species in common (Muller-Dubois and Ellenberg, 2003). Based on this criterion, our cluster analysis revealed high similarity levels among the different vegetation types. While the lowest similarities obtained were between lowland and gallery forest, the remaining comparisons revealed similarity values greater than 75%. Similarity in species composition occurs when the analysis is focused on the same type of vegetation (Rodrigues and Nave, 2000). One of the factors that influenced the grouping was forest size and proximity to other vegetation types: Transitional forest was very close with lowland forest, while others were distant from each other, indicating that distance is an aggregation factor (Kuntz et al., 2009). Figure 5 shows a dendrogram showing similarities between the different vegetation types. The distance correlation (ward linkage) between lowland forest and gallery forest is minimal and this shows that lowland forest and gallery forest have many plants species that are similar and their similarity index was 75% while the grassland was less similar to all the other vegetation types.
The majority of the taxa found in the studied area are of conservation value and importance. They occur mostly in the intricate mosaic of lowland and gallery forest, and the ecological fragility and anthropogenic pressure on the lowland forest suggest that these ecotypes are of considerable conservation value. Out of the 222 species recorded, 5 species (2.80%) were vulnerable according to the IUCN (Sainge, 2016). These species included: A. africana, Afzelia bipindensis, Allophyllus bullatus, Entandrophragma angolense, Hallea stipulosa, Quassia sanguine. However, A. africana was still abundant locally in the woody savanna of the Kimbi Fungom National Park. These species thus need high conservation effort.
The Kimbi Fungom National Park is an ecoregion with four major vegetation types; lowland rainforest, gallery forest, woody savanna and grassland savanna. These four main vegetation types had sub vegetation types like the swampy forest which could be found in any of the major vegetation type, and the secondary which were heavily disturbed gallery forests. Species richness was high in the lowland forest than any other forest types. The most abundant family recorded was Fabaceae with 31 species. Most of the forest had a high density of small species of less than 9.9 cm. The renewal of species through the regeneration was weak, the vulnerability of young plants to grazing, drought and farming and most importantly bush fire slow down the sustainability of woody species. This is termed an ecological, and environment problem that is contributing to the degradation of the floras in the park. The woody and grassland savannas need high conservation effort as they are always burnt, and are also fertile grazing environments.
The management, preservation systems including local population and habitat regeneration will therefore be highly advantageous to save this protected area from destruction, save their flora and fauna species from local extinction and to maintain a viable population size in the face of growing anthropogenic activities. It should be noted that the K-FNP species richness and its diversity are under serious threats due to the anthropogenic pressure resulting from both cash crops and food crop production and the lack of government interest since its creation before 2015. Serious habitat destruction and illegal logging are still very active in the park as the exploitation of the Pterocarpus erinaceus remained the greatest problem. The lowland and the woody savanna need urgent conservation effort. Measures should therefore be taken toward good management and monitoring of this park through frequent patrol and the recruitment of more forest guards. Measures to support the regeneration of tree species should also be taken, in order to increase the main abundance of trees and threatened species. However, successful restoration requires involvement from many disciplines and stakeholders, from government and ecologists to local communities, and from decision makers to ordinary people. This will improve the species pocket of the park and hence the park will remained a scientific laboratory of biodiversity experimentation.
The author has not declared any conflict of interests.
REFERENCES
|
Achoundong G (2007). "Vegetation," In: N. Houstin and C. Seignobos, Ed., Atlas of Cameroon' Les éditions Jeune Afrique, Paris pp. 64-65.
|
|
|
|
Azani N, Babineau M, Bailey CD, Banks H, Barbosa AR, Pinto RB, Boatwright JS, Borges LM, Brown GK, Bruneau A, Candido E (2017). A new subfamily classification of the Leguminosae based on a taxonomically comprehensive phylogeny The Legume Phylogeny Working Group (LPWG). Taxon 66(1):44-77.
Crossref
|
|
|
|
|
Alpert P (1993). Conserving biodiversity in Cameroon. Ambio 22:33-1071.
|
|
|
|
|
Barthlott W, Lauer W, Placke A (1996).Global distribution of Species diversity in vascular plants: towards a world map of phytodiversity. Erkunde band 50:317-328.
Crossref
|
|
|
|
|
Beech E, Rivers M, Oldfield S, Mith PP (2017). Global tree search: The first complete global database of tree species and country distributions. Journal of Sustainable Forestry 36(5):454-489.
Crossref
|
|
|
|
|
BirdLife International (2010). Important Bird Areas factsheet: Mbi Crater Faunal Reserve Mbingo forest. Downloaded from http://www.birdlife.org on 12/04/2013.
|
|
|
|
|
Boussim J, Ouédraogo A, Lankoandé B (2009). Etude des impacts écologiques dans les unités d'aménagement forestier des régions du Centre-Nord et du Centre-Ouest. In: Kabré AM, Somda J, Savadogo M and Nianogo AJ (eds), Bois-énergie au Burkina Faso: Consolidation des moyens d'existence durable (2000-2009). [Study of ecological impacts in the adjustment forest unit of the Center-North and the Mid-west areas. In: Kabré AM, Somda J, Savadogo MR, Nianogo AJ (eds): Wood-fuel in Burkina Faso: Consolidation the way of sustainability], Ouagadougou, Burkina Faso: Bureau, UICN-Burkina Faso pp. 115-139.
|
|
|
|
|
Buckland ST, Borchers DL, Johnston A, Henrys PA, Marques TA (2007). Line transect methods for plant surveys. Biometrics 63:989-998.
Crossref
|
|
|
|
|
Chowdhury S (2005). Assam's Flora, Present Status of Vascular Plants. Assam Science Technology and Environmental Council, Guwahati, 368 p.
|
|
|
|
|
Chuyong GB, Kenfack D, Harms KE, Thomas DW, Condit R, Comita LS (2011),"Habitat specificity and diversity of tree species in an African wet tropical forest" Plant Ecology 212(8):1363-1374.
Crossref
|
|
|
|
|
Condit R (2008). Methods for estimating above¬ground biomass of forest and replacement vegetation in the tropics.Center for Tropical Forest Science Research Manual 73 p.
|
|
|
|
|
Daws MI, Mullins CE, Burslem DFRP, Paton SR, Dalling JW (2002). Topographic position affects the water regime in a semideciduous tropical forest in Panama. Plant Soil 238:79-90.
Crossref
|
|
|
|
|
Federal Geographic Data Committee (FGDC) (2005). Nacional Vegetation Classiffication Standard Hierarchy Revisions Working Group. Federal Geographic Data Committee, Vegetation Subcommittee. OverView July 2005. In:
View.
|
|
|
|
|
Focho DA, Nkeng EA, Fonge BA, Fongod AN, Muh CN, Ndam TW, Afegenui A (2009). "Diversity of plants used to treat respiratory diseases in Tubah, northwest region, Cameroon," African Journal of Pharmacy and Pharmacology 3(11):573-580.
|
|
|
|
|
Focho DA, Ndam WT, Fonge BA (2009). "Medicinal plants of Aguambu-Bamumbu in the Lebialem highlands, southwest province of Cameroon," African Journal of Pharmacy and Pharmacology 3(1):1-13.
|
|
|
|
|
Fonge BA, Focho DA, Egbe EA, Tening AS, Fongod AN, Neba GA (2011). "The effects of climate and edaphic factors on plant colonisation of lava flows on Mount Cameroon," Journal of Ecology and the Natural Environment 3(8):255-267.
|
|
|
|
|
Fonge BA, Tchetcha DJ, Nkembi L (2013). Diversity, Distribution, and Abundance of Plants in Lewoh-Lebang in the Lebialem Highlands of Southwestern Cameroon. International Journal of Biodiversity 2013:13.
Crossref
|
|
|
|
|
Hall JB, Swaine MD (1981). Distribution and ecology of vascular plants in a tropical rain forest. Dr W Junk Publishers, The Hague.
Crossref
|
|
|
|
|
Hall JS, McKenna JJ, Ashton PMS, Gregoire TG (2004). Habitat characterizations underestimate the role of edaphic factors controlling the distribution of Entandrophragma. Ecology 85:2171-2183.
Crossref
|
|
|
|
|
Hardy OJ, Sonke B (2004). Spatial pattern analysis of tree species distribution in a tropical rain forest of Cameroon: assessing the role of limited dispersal and niche differentiation. Forest Ecology and Management 197:191-202.
Crossref
|
|
|
|
|
Harms KE, Condit R, Hubbell SP, Foster RB (2001). Habitat associations of trees and shrubs in a 50-ha neotropical forest plot. Journal of Ecology 89:947-959.
Crossref
|
|
|
|
|
Kenfack D, Thomas W, Chuyong G, Condit R (2007). "Rarity and abundance in a diverse African forest," Biodiversity and Conservation 16(7):2045-2074.
Crossref
|
|
|
|
|
Kent M, Coker P (1992). Vegetation Description and Analysis, Belhaven Press, London, UK.
|
|
|
|
|
Kuntz SH, Ivanauskas NM, Martins SV, Silva E, Stefanello D (2009). Análise de similaridade florística entre florestas do alto rio Xingu, da bacia amazônica e do planalto.
|
|
|
|
|
Letouzey R (1985). Notice de la carte phytoge’ographique du Cameroun au 1:500.000. Institut de la Carte Internationale de la Vegetation, Toulouse.
|
|
|
|
|
Matos FDA, Amaral IL (1999). Análise ecológica de um hectare em floresta Ombrófila de Terra- Firme, Estrada da várzea, Amazonas, Brasil. Acta Amazonica 23:365-379.
Crossref
|
|
|
|
|
Mbolo M (2002). "La collecte et l'analyse des donn’ees statistique sur les produits forestiers non ligneux: une ´etude pilote au Cameroun," D’epartement des forets. In Programme produit forestiers, non ligneux, FAO, Rome, Italie.
|
|
|
|
|
Memiaghe HR, Lutz JA, Korte L, Alonso A, Kenfack D (2016). Ecological importance of small-diameter trees to the structure, diversity and biomass of a tropicsl evergreen forest at Rabi, Gabon. PloS ONE 11:e0154988.
Crossref
|
|
|
|
|
Miranda IS (1993). Estrutura do estrato arbóreo do cerrado amazônico em Alter do Chão, Pará, Brasil. Revista Brasileira de Botânica 16:143-150.
|
|
|
|
|
Miyamoto K, Suzuki E, Kohyama T, Seino T, Mirmanto E, Simbolon H (2003). Habitat differentiation among tree species with small-scale variation of humus depth and topography in a tropical heath forest of Central Kalimantan, Indonesia. Journal of Tropical Ecology 19:43-54.
Crossref
|
|
|
|
|
Moleele NM, Perkins JS (1998). Encroaching woody plant species and boreholes: is cattle density the main driving factor in the Olifants Drift communal grazing lands, south Barodewol Botswana? Journal of Arid Environments 40:245-253.
Crossref
|
|
|
|
|
Mueller-Dombois D, Ellenberg H (1974). Aims and Methods of Vegetation Ecology. John Wiley and Sons, New York, 547p.
|
|
|
|
|
Ndam N, Nkefor JP, Blackmore P (2001). "Domestication of Gnetum africanum and G.buchholzianum (Gnetaceae), Overexploited wild forest vegetables of the Central African Region," Systematics and Geography of Plants 71(2):739-745.
Crossref
|
|
|
|
|
Ndenecho EN (2005). Conserving biodiversity in Africa: Wildlife management in Cameroon. Loyola Journal of Social Sciences 2:209-228.
|
|
|
|
|
Ndenecho EN (2009). Ecological Planning and Ecotourism Development in Kimbi Game Reserve, Cameroon. Journal of Human Ecology 27(2):105-13.
Crossref
|
|
|
|
|
Ngwabuh BA (2002). Annual Report of the Kimbi Game Reserve. Bamenda: NW Delegation for Environment and Forest.
|
|
|
|
|
Ntoupka M (1999). Impacts des perturbations anthropiques (pâturage, feu et coupe de bois) sur la dynamique de la savane arborée en zone soudano-sahélienne nord du Cameroun. Thèse de doctorat. Université Paul Valéry. (Impacts of anthropic disturbances (pastures, bushfire and wood-cutting) on the dynamics of the sounado-Sahelian savannah zone in Gonoray Cameroon. PhD Dissertation. University Paul Valery] P 233.
|
|
|
|
|
Onana JM (2011). The Vascular Plants of Cameroon.A Taxonomic Checklist with IUCN Assessments. Flore Du Cameroun Volume 39 "Occasional volume".
|
|
|
|
|
Onana JM, Cheek M (2011). The Red Data Book of the Flowering Plants of Cameroon.RBG, Kew. 578 p.
|
|
|
|
|
Oosting HJ (1956). The study of plant communities. W. H. Freeman and Co., San Francisco.
|
|
|
|
|
Parmentier I, Malhi Y, Senterre B (2007). The odd man out? Might climate explain the lower tree alpha-diversity of African rain forests relative to Amazonian rain forests? Journal of Ecology 95:1058-1071.
Crossref
|
|
|
|
|
Pitman NCA, Terborgh JW, Silman MR, Nu-ez PV, Neill DA, Cerón CE, Palacios WA, Aulestia M (2002). A comparison of tree species diversity in two upper Amazonian forests. Ecology 83:3210-3224.
Crossref
|
|
|
|
|
Potts MD, Davies SJ, Bossert WH, Tan S, Nur Supardi MN (2004). Habitat heterogeneity and niche structure of trees in two tropical rain forests. Oecologia 139:446-453.
Crossref
|
|
|
|
|
Rankin-de-Merona J, Prance GT, Hutchings RW, Silva MF, Rodrigues WA, Vehling ME (1992). Preliminary results of a large-scale tree inventory of upland rain forest in the Central Amazon. Acta Amazonica 22:493-534.
Crossref
|
|
|
|
|
Rodrigues RR, Nave AG (2000). Heterogeneidade florística das matas ciliares.In: Rodrigues RR, Leitão-Filho HF (Ed.). Matas ciliares: conservação e recuperação. Edusp/Fapesp, São Paulo, pp. 45-71.
|
|
|
|
|
Russo SE, Brown P, Tan S, Davies SJ (2008). Interspecific demographic trade-offs and soil-related habitat associations of tree species along resource gradients. Journal of Ecology 96:192-203.
Crossref
|
|
|
|
|
Sainge MN (2016). Patterns of distribution and Endemism of Plants in the Cameroon Mountains: A case study of Protected Areas in Cameroon: Rumpi Hills Forest Reserve (RHFR) and the Kimbi Fungom National Park (KFNP). Tropical Plant Exploration Group (TroPEG) Cameroon.
|
|
|
|
|
Savadogo P, Tigabu M, Sawadogo L, Odén PC (2007). Woody species composition, structure and diversity of vegetation patches of a Sudanian savanna in Burkina Faso, Bois et Forêts des Tropiques 294(4):5-20.
|
|
|
|
|
Sunderland TCH, Comiskey JA, Besong S, Mboh H, Fonwebon J, Dione MA (2003.)"Vegetation Assessment of Takamanda Forest Reserve, Cameroon," Smithsonian Institution.
|
|
|
|
|
Tata FT (2011). Stakeholder Participation in the Establishment of Biosphere Reserves: The Case of Local Communities in the Proposed Kimbi-Dumbo-Nyos Conservation Zone, Cameroon. Discussion Paper presented by Dr. Thomas Fofung Tata at the International Expert Workshop on Managing Challenges of Biosphere Reserves in Africa, Dresden, Germany, June 2011.
|
|
|
|
|
Tata FT (2015). The Kimbi-Fungom National Park; -A situation report. Green works company Ltd. P.O. Box, Bamenda, Cameroon.
|
|
|
|
|
Ter Steege H, Pitman NC, Phillips OL, Chave J, Sabatier D, Duque A, Molino JF, Prévost MF, Spichiger R, Castellanos H, Von Hildebrand P (2006). Continental-scale patterns of canopy tree composition and function across Amazonia. Nature 443(7110):444.
Crossref
|
|
|
|
|
Thomas DW, Kenfack D, Chuyong GB, Sainge MN, Losos EC, Condit RS, Songwe N (2003). Tree species of south- western Cameroon: Tree distribution maps, diameter tables, and species documentation of the 50-hectare Korup Forest Dynamic Plot. Washington, D.C.
|
|
|
|
|
Uno G, Storey R, Moore R (2001). Principles of Botany, McGraw-Hill, 2001.
|
|
|
|
|
Velázquez A, Romero FJ (eds) (1999). Biodiversidad de la región de monta-a del sur de la Cuenca de México: bases para el ordenamiento ecológico. UAM-X-SEMARNAP. ISBN: 754 24 2867 8.
|
|
|
|
|
Webb CO, Peart DR (2000) Habitat associations of trees and seedlings in a Bornean rain forest. Ecology 88:464-478
Crossref
|
|
|
|
|
Whittaker RH (1970). Communities and Ecosystems. Collier-MacMillan, Canada, Ltd, Toronto, Canada.
|
|
|
|
|
White F (1983). The vegetation of Africa. UNESCO, Paris.
|
|
|
|
|
Wilson EO (1992). The Diversity of Life. Cambridge, MA: Belknap.
|
|
|
|
|
Yamada T, Zuidema PA, Itoh A, Yamakura T, Ohkubo T, Kanzaki M, Tan S, Ashton PS (2007). Strong habitat preference of a tropical rain forest tree does not imply large differences in population dynamics across habitats. Journal of Ecology 95:332-342.
Crossref
|
|