Full Length Research Paper

ABSTRACT
Interesting morphological traits in tree organs are essential for selecting the best plant germplasm. Variation in morphometric traits of leaf, fruit and seed of multipurpose tree Diospyros mespiliformis were studied in two climatic zones in Benin using 735 trees from 4 major habitats (provenances) in each climatic zone. Morphological trait measurements were combined with architectural parameters and analyzed using two-way ANOVA, principal components analysis and hierarchical clustering. Results indicated that significant differences were observed between leaves, fruits and seed morphological traits within climatic zones and habitats in terms of dimensions across the study area. Compared to the other three habitats (low dimensions), leaves from woodland showed large dimensions (µL_leaf=130 mm, µl_leaf=50 mm). Soudano Guinean zone recorded the highest fruits morphological traits values (µd_fruit= 36 mm µl_fruit=30 mm) while Soudanian zone had the lowest (µd_fruit= 12 mm, µl_fruit=8 mm). More seeds per fruit were recorded in woodlands and parklands. Out of these, hierarchical analysis grouped ebony morphotypes into four clusters. There was no significant correlation between the number of seeds per fruits and other fruit traits. However, significant and strong positive correlations were found between morphometric characters, bearing and architectural parameters (R = 0.96). The provenance significantly affected variations in organ-related traits. Correlation relationships suggested morphotypes for breeding improvement. Interesting and desired characteristics delineating individuals and populations can guide future selection of targeted ebony trees with the aim of improving high-value species in agroforestry domestication program.
Key words: Diversity, domestication, environmental factors, morphological descriptors, multivariate analysis.
INTRODUCTION
Removal of forest flora contribute to crucial global environmental concerns such as climate change and loss of biodiversity in addition to the regional and local problem (Romeiras et al., 2018; Okpanachi et al. (2019); Wade et al., 2018; Tiokeng et al., 2019). Local conservation agencies must contend with the loss of genetic resources and associated traditional knowledge (Catarino et al., 2019). Forests are subject to haphazard modification following anthropogenic pressures including tree felling for agriculture. Over time, human activities have led to the destruction of economic and culturally important tree species (Kimpouni et al., 2019). In order to reduce the risk of extinction, urgent actions are needed to develop protective, domestication, propagation and valorization programs for food and woody tree species.
Also, all forms of domestication begin with the exploitation of the existing natural variability while selecting, with the help of the populations, the topics to the most interesting phenotypes for the considered criteria, designated as ' trees plus' (Dicko et al., 2019). Morphological variability study is appropriate for overall genetic improvement and tree varietal selection activities. It enables the identification of attractive morphological descriptors; traits linked to the origin of seeds sources and possible genetic groups. Indeed, the phenotypic variation observed for plants is generally a response to differences in climatic conditions that reflects either adaptive evolution or phenotypic plasticity, or a combination of both. In Benin Republic, the variability of woody savannah and indigenous fruit species is poorly studied, when ecosystems that support them are highly threatened and native species are lost, along with their gene pools.
Diospyros mespiliformis (Ebenaceae) is an evergreen tree, found in the tropical forest of Sub-Saharan Africa; from Ethiopia to Swaziland (Wallnöfer, 2001). It is also found in Angola, Nigeria, South Africa, Tanzania, Uganda, Yemen, Republic of Zambia and Zimbabwe (Arbonnier, 2002). D. mespiliformis, commonly known as African ebony tree, is an indigenous fruit tree species widespread in the Soudano Guinean and Soudanian zones in Benin. The fruits of the species are edible, while the bark, leaves, stem and roots are employed for various purposes (Gnonlonfin at al., 2018). Wide environmental and geographical variation often occurs within the natural range of the plant species (Akoegninou et al., 2006). Adaptation of a species to this variation may produce different morphological and physiological characteristics, resulting in the development of ecotypes. Therefore, the conservation of a forest species requires knowledge on morphological variability to differentiate individuals and target interesting morphotypes. With an increasing demand for products and by-products of ebony trees, their supply and diversity are threatened by increasing deforestation. Therefore, the necessity for domestication (which involves selection of elite trees for multiplication) is paramount.
As all species are distributed by habitat fragmentation or other anthropogenic factors, Beninese ebony diversity is declining (Abasse et al., 2011). Therefore, enriching quantitative databases and monitoring programs for conservation is of great significance. Moreover, ebony species play a central role in many ecosystems. There is paucity of studies on ebony morphology in Benin. Based on a literature search, there is evidence indicating the existence of a number of local types differing in habitat, vigor, and size, quality of fruits and vitamin content of species leaves, fruits and seeds (Mkwezalamba, 2015). However, gaps in research still exist in eliciting different morphological traits, check-listing plus-trees, cataloging and monitoring of ebonies. Consequently, it therefore is highly essential to evaluate in detail the species’ genotype at the morphological level with the aim of providing knowledge on which source to base plus-tree selection for plant resource tree quality improvement and conservation. This is with the view to conserve the germplasm of indigenous plant species and contain biodiversity loss that may lead to extinction. Specifically, this study aims to: (1) identify different morphological groups within D. mespiliformis based on morphological traits of interest and; (2) analyze the influence of habitat-type and climatic zone on the morphological variability of D. mespiliformis
MATERIALS AND METHODS
Study area
This study was carried out in the two northern climatic zones of Benin Republic where the species is predominant. This zone lies between latitudes 7° 30’ N and 12°40’ N, and longitudes 1°6’ E and 3°45’ E. The population of the study area is estimated at 2,941,180 inhabitants (INSAE, 2015). Livelihood activities carried out by the inhabitants include subsistence agriculture, ranching, fishing and hunting, trade and craft (Dicko, 2016).
Characteristics of D. mespiliformis
D. mespiliformis is one of the most important native in the wild species introduced in agro-forestry systems in Sahelian Africa (Gnonlonfin et al., 2018). Adult trees can grow up to 20 -50 m in height, and 150-300 cm diameter in breast height. Leaves are alternate; shiny-green above and paler beneath, 3.5-19 cm long, 1.5-7.5 cm wide, oblong elliptic. Flowers are pentamerous, white and fragrant. Flowering starts in April-May during the rainy season and fruits reach maturity in November-January during the dry season. Fruits are usually globose, fleshy, up to 3 cm in diameter, greenish and pubescent when young, yellowish to orange yellow and glabrous when ripe with dark brown seeds (Arbonnier, 2002).
Sampling population and data collection
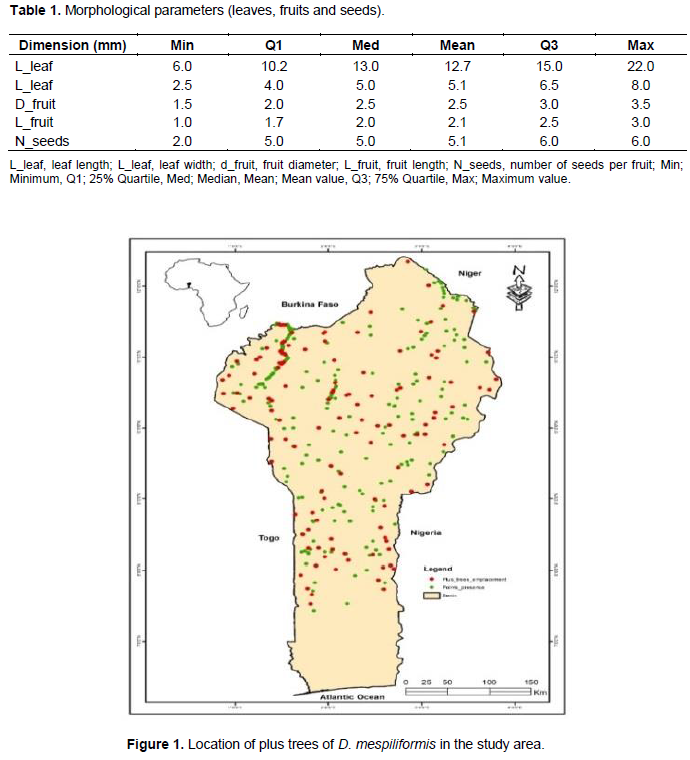
All structural and morphological characteristics were recorded using the KoBo toolbox for smartphones according to Olajide (2019). A multi-stage sampling procedure was applied which involved purposive sampling. Data was collected from areas where D. mespiliformis occurred from April 2017 to October 2018. A total of 22 villages were selected based on the presence of at least one adult individual D. mespiliformis (DBH > 10 cm). Along with the selected villages and for each habitat (woodlands “tree with heavy DBH” savannah “tree with weak DBH” in natural or protected area and parklands in agricultural systems or non-protected area and inundated forests) (Arbonnier, 2002), four circular or rectangular plots of 1000 m2 were randomly installed. This resulted in 16 plots of 1000 m2 per village. In total, 263 circular and 88 rectangular plots were installed along the study area. In each plot, vegetation type and species other than D. mespiliformis (adult individual and regeneration) (Arbonnier, 2002; Akoegninou et al., 2006) were recorded using species key determination or in-site recognition based on field experience. All D. mespiliformis trees were inventoried and registered via a Global Positioning System (GPS). The recorded geographical coordinates of the central tree in the plot were charted onto a map of Benin (Figure 1).
Morphological data of D. mespiliformis populations
For morphological characterization, the following variables were collected according to Ouinsavi (2010): organ descriptive parameters (leaf length from the stalk to the apex of the leaf, leaf width at the center of the leaf, mature fruit diameter (globosely form of the fruit), and fruit length using Vernier caliper and the number of seeds per fruit (by counting) combined with bearing and architectural parameters (stem diameter at breast height, total height, bole height, and crown diameter). Bearing and architectural parameters of the sampled trees were measured while organ descriptive parameters were measured on five leaves and fruits each, randomly collected from adult trees. For each fruit, number of seeds was recorded. A total of 1755 leaves and 1585 fruits were measured (Ouinsavi, 2010; Dadegnonet et al., 2015).
Environmental factors (climatic zones and habitats)
The study area lies between two climatic zones of Benin: Soudanian climatic zone (between 9°45’ - 12°25’ N) and Soudano-Guinean climatic zone (between 7°30’ - 9°45’ N). Rainfall in these two zones is unimodal (Adomou, 2005).
The Soudanian climatic zone is a woodland and savannah region with more ferruginous soils. The mean annual temperature in this zone is 35°C while mean annual rainfall varies between 900 - 1100 mm. The Soudano-Guinean climatic zone is a transitional zone between the sub humid Guinean and Soudanian zones. This zone is characterized by a vegetation mosaic of forest islands, gallery forests and savannahs.
Data treatment and analysis
For organ descriptive parameters (leaves and fruits dimensions) measurements, data were averaged for individual trees before undertaking the series of multivariate analysis using appropriate procedures. Then a generalized linear model was used to test the effect of climate zones and habitats on tree organ descriptive parameters (leaves and fruits dimensions). For the number of seeds per fruit the study used a generalized linear model of the Poisson family test in R.
Two-way ANOVA was used to analyze the effect of climatic zone and habitat on morphological variation. Five significant codes were used : ***Pr=0.001, **Pr=0.01, *Pr=0.05, Pr=0.1’Pr=1. Thus, principal component analysis (PCA) was performed on the untransformed morphometric data using the correlation matrix (Kouyaté, 2005; Rindyastuti, 2021). Correlogram and dendrogram were finally generated to explain the degree of relationship between bearings and architectural and tree organ descriptive parameters (Juma et al., 2020).
RESULTS
Morphological parameters and occurrence of ebony trees in Benin
The largest values for leaf length (220 mm) and width (80 mm as well as fruit diameter (30 mm) and width (60 mm) and the number of seed per fruit (6), were in woodlands and parklands of Soudano-Guinean transition zone; while the lowest (60, 25, 15, 10, 20 and 2 mm) respectively were obtained in inundated forests of Soudanian zone (Table 1 and Figure 1).
Effect of environmental climatic zones and habitats on leaves, fruits and seeds variation leaf characteristics
The pairwise comparison of leaf dimensions of the ANOVA output indicated a high degree of variability in leaf shapes at tree level. Thus, five leaf shapes that is, elliptic, oblong, oblong-elliptic, oblolanceolate-elliptic and lanceolate-elliptic were recorded across the 735 trees studied in the two climatic zones and four habitats (Figure 2). About 74% of the trees exhibited leaf shapes commonly listed in the description of D. mespiliformis (Figure 3).
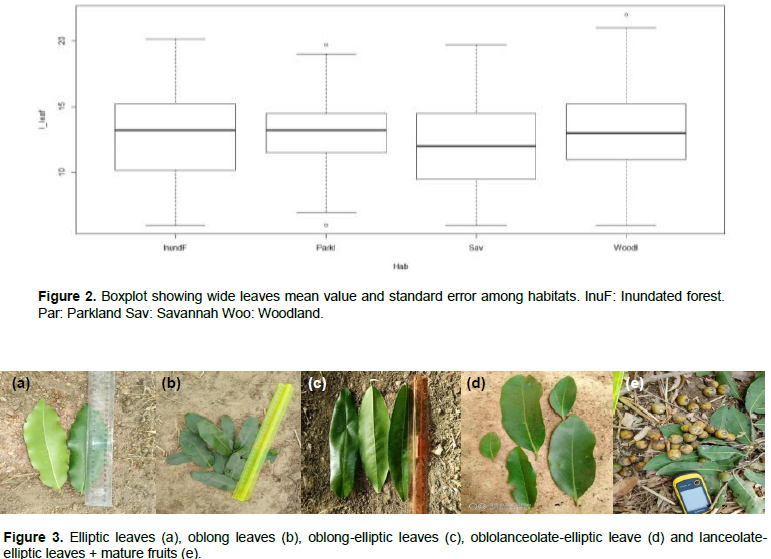
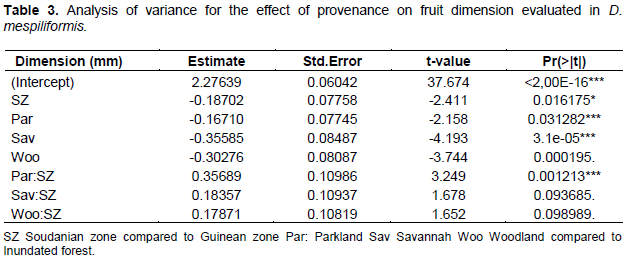
Table 2 indicated significant differences in morphological effect of leaf dimensions. We note that between Soudano-Guinean climatic zone and Soudanian zone, respectively, habitat associated probability is low at Pr < 0.0013. Significant differences observed among climatic zones and habitats for leaf length and width indicated high variation in leaf form across climatic zones and habitats. Indeed, field observations revealed that leaves from the Soudano Guinean transition zone were larger than those harvested in Soudanian zone. Likewise, leaves from swampy forests were larger than those from other habitats. This can be explained by the fact that Soudano Guinean transition zone is the zone with higher rainfall than the Soudanian zone. This suggests that D. mespiliformis is a wide range tree and several leaf traits can be selected from any climatic zone or habitat: there is a potential for genetic selection among individuals based on leaf morphology (Table 2 and Figure 1). R proposed also the boxplot of mean value with standard error associate to each habitat (Figure 1). For instance, this figure shows that the three habitats (Inundated forests, parkland and savannah were not significantly different according to the plant part dimensions. Also, for the habitats woodland, savannah and inundated forests dataset on plant organ dimensions of these three habitats were not significantly different. Plant part dimensions (leaves, fruits) of parkland were significantly different from other habitats (woodland).
Fruit characteristics
An analysis of fruit characteristics of ebony plants revealed 4 fruit shapes. Data on the distribution of the four most common fruit shapes are shown in Figure 4.

Results indicated significant morphological effect for fruit dimensions. Significant differences observed among climatic zones and habitats for fruit diameter and width indicated high variation in fruit form across climatic zones and habitats. Indeed, field observations revealed that fruits from Soudano Guinean transition zone were larger than those harvested in Soudanian zone. Likewise, fruits from swampy forests were larger than those from other habitats. This can be explained by the fact that Soudano Guinean transition zone is the zone that is better provided in rain than Soudanian zone. This suggests that D. mespiliformis is a wide range tree and several fruit traits can be selected in any climatic zone or habitat: there is a potential for genetic selection among individual based on the form of fruit (Table 3 and Figure 2).
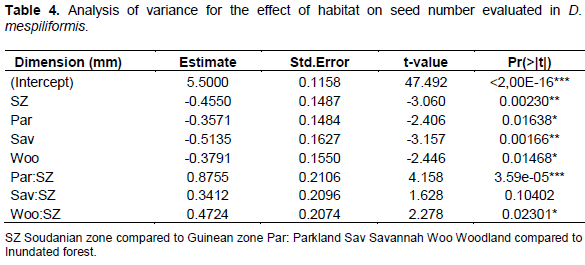
Seeds number per fruit
Table 4 shows the result of analysis of variance of number of seeds as influenced by climatic zones and habitats. Number of seed differed significantly with respect to all zones and habitats. Number of seed was about 2-3 for Soudanian zone and 4-6 for Soudano-Guinean transition zone. However, this was not different in savannah, woodland and parkland habitats compared to swampy forests which was significantly different.
In total, significant differences for a variety of traits of economic importance (mainly fruit form and number of seeds per fruit) are a basis of genetic selection programs (genetic research). For the morphological parameters (leaf length and width, fruit diameter and length and the number of seeds per fruit) the generalized linear model (Poisson regression model) indicated significant difference between habitats (Tables 3 and 4) and climatic zones (Tables 2 to 4) (Pr<0.05); while there is no significant difference between habitats and climatic zones for leaf dimensions (Pr>0. 05).
Morphological variations among individuals of D. mespiliformis
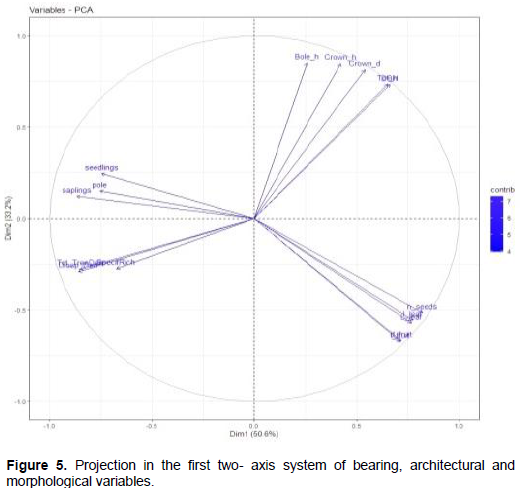
In order to confirm the previous association between quantitative variables, Principal Components Analysis (PCA) was performed on bearing and architectural parameters as well as tree organs descriptor parameters. The principal component analysis showed that morphological variation (83.8%) was explained by the first two principal axes (Figure 5). Morphological trait coefficient (that is, Eigen vectors) indicated that DBH, total height, bole height, crown diameter and height were the loading variables that are positive in the first axis while leaf dimensions (length and width), fruit dimensions (diameter and length) and number of seeds were the loaded variables that are negative on the second principal axis (Figure 5). Along the principal component axis 1, most of individuals from savannah and parkland occupied the right side whereas the mixed group (individuals from all habitats) occupied the center as well as the left side (Figure 5). The populations on first axis were differentiated on the basis of tree height and leaf width. On the principal axis 2, most of the individuals from inundated forest and woodland occupied the middle lower part while the mixed populations occupied the middle around the central point. It is concluded that there is a great link between certain bearing parameters and tree organ descriptive parameters.
Scatter plot matrix of all bearing, architectural and morphological parameters
To classify individuals into homogenous groups with similar characteristics, database on bearing and architectural characteristics and tree organ descriptive parameters were used to make a correlation matrix. This correlation matrix was subjected to pairwise analysis to determine the correlation coefficients between the sixteen variables and the significance levels using the performance analytics package (Figure 6).
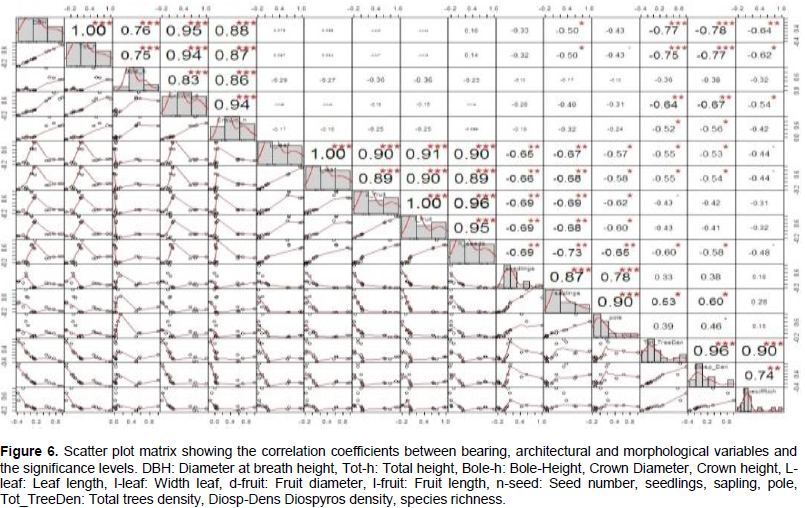
This graph reveals that variables such as DBH, total height, bole height, crown diameter and crown height are highly positively correlated on one hand and leaf length was highly correlated with leaf width and the same regarding dimensions of fruit. The absence of a high link between bearing and architectural parameters and tree organ descriptive parameters indicates the possible way of homogenous groups of individuals of D. mespiliformis from different habitats.
Hierarchical clustering and principal component’s analysis
The dendrogram ensuing from the Ward D2 method sorted the 735 ebonies samples originating from the two climatic zones into four major groups (G1, G2, G3 and G4) with each group containing samples from at least three habitats (Figure 7). The largest group was G1 followed by G4 and G2 and lastly G3; the 735 ebonies samples were sorted into each of these group. Sorting individuals into the groups was based on tree descriptive organ which unveiled four homogenous groups: The first cluster composed mainly individuals from all habitats with middle fruits and leaves; the 2nd cluster contained mainly individuals from parkland and inundated forests with large fruits and leaves; 3rd group composed mainly individuals from inundated forests and savannah with small and middle fruits and leaves and the 4th group is composed of individuals from riparian soil from all habitats but only located in the north east (Natitingou-Tanguieta-Cobbly-Toukountouna) with small fruits and leaves only. Individuals from all habitats which composed group 1 have the largest organ dimensions size (leaves mean 22-8, fruit 3-3.3) and are both from Banikoara parkland and Alafiarou woodland (Figures 6 to 8). In particular, the lack of clear grouping of the 735 samples in relation to their habitats of origin indicate the exchange of seeds between several provenances (here swampy forests, woodlands, savannah and parklands) (Figures 7 and 8).
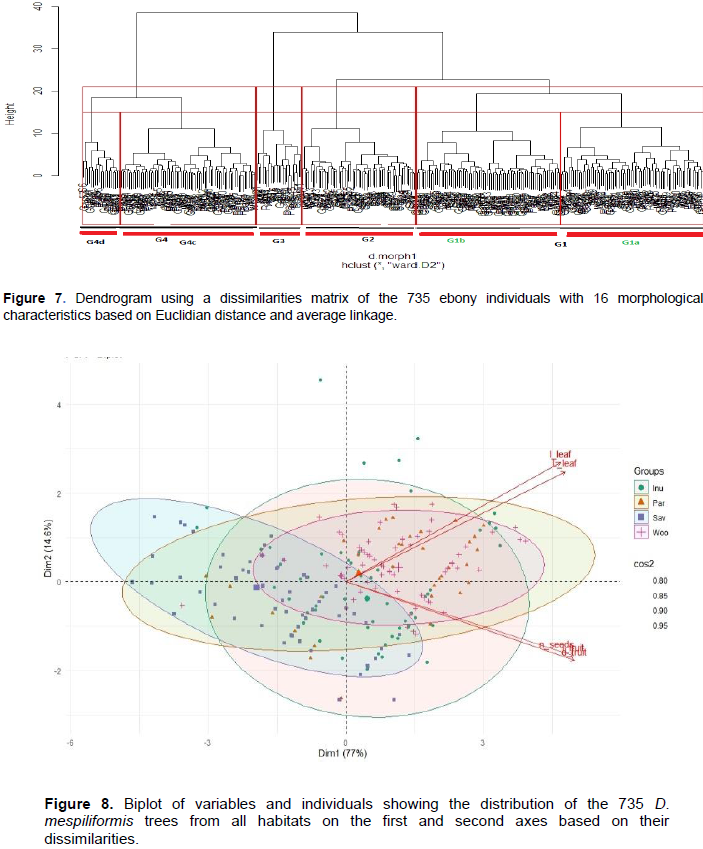
The PCA on the ebony qualitative and quantitative morphological traits revealed extensive variation among the sampled trees at the habitat level, with four clear grouping of samples (Figure 6). The first two axes explained 91.6% of the total variation, corresponding to 77.00 and 14.6% for the first and second axis, respectively.
DISCUSSION
Leaves and fruits shapes (qualitative) variation across provenances
Knowledge on intraspecific diversity of ebony trees is fundamental in order to fulfill the goal of meeting the demands of subsistence as well as the wellbeing of farmers (Assogbadjo et al., 2005). Qualitative and quantitative approaches were used determine the relationship between morphological structure according to the considered climatic zone and habitats. Regarding the qualitative approach, several shapes of ebony tree organs (leaves and fruits) were identified through climatic zone and habitats in the study area. This diversity of leaf shapes could be due to phenotypic factors influenced by climatic zone diversity and inside site conditions. The results of the study contribute in filling the gap of information on African ebony phenotypic diversity in leaf and fruit characteristics for use in domestication and tree improvement processes (Maroyi, 2018a, b). The results of the present research work indicated that morphological traits in leaf and fruit were strongly influenced by environmental factors.
Leaves and fruits measurements (quantitative) variation among environmental factors
The important variability between climatic zones and habitats observed for some characters as the measurements of the leaves and fruits could be bound more to environmental conditions than to genotypic factors. This justification finds its foundation in the fact that an interrelationship would exist enters the quantitative morphological features and the genetic data of the individuals of the species (IPGRI, 2016). The micro - variations of soil characteristics and to a certain extent, anthropic effects and the parasitic attacks that can return the shrunken plant can be added to the main factor (Gbemavo et al., 2015). The weak variability intra and inter under - populations observed for the morphological fruit descriptors (diameter and length) with exception to the number of seeds by fruit could be explained by the crossed fertilization, since D. mespiliformis is a dioïc species. Thus, fruit and leaf dimensions variability observed among individuals is probably due to the effects of the environment (Kouyaté, 2005).
Concerning the four morphotypes (under - populations) of D. mespiliformis gotten by the ascending hierarchical classification of Ward D2, a strong variability intra and inter morphotypes (under - populations) of most studied morphological descriptors was observed. Moreover fruit and leaf variability indicates that these four groups of D. mespiliformis showed phenotypic differences. This suggested a strong genetic diversity of the species. These results is in agreement with the study of Boukary et al., 2010) on D. microcarpum, Assogbadjo et al. (2005) on Adansonia digitata; Mkwezalamba (2015) on the fruits of Sclerocarya birrea and Abasse et al. (2011) on Balanites aegyptiaca, but not in agreement with the study of Gbemavo et al. (2015) on the fruits and the seeds of the Jatropha curcas. This strong variability to the level of the fruits can be bound in particular also to the phenotypic characters shown in the survey. Indeed, Gbemavo et al. (2015) showed other factor such as the period of flowering, the type and number of inflorescence, the color and the texture of the leaves are the quantitative features that contribute to the variations of the plant. These characters are not considered in this survey. To these factors, can be added weak influences of the environment and genotypic interaction x environment on the studied characters.
Based on these results and for a better valorization of the species in order to get better outputs of fruits and the pulp (only edible part of the fruit with interesting qualities), it is necessary to proceed with a selection inside the group 1 of Diospyros mespiliformis individuals from gallery (inundated or swampy) forests and parklands of Dunkassa, Bembérèkè, Kérou, Banikoara, Kouandé and Alafiarou where the fruits are thicker (Diameter > 3 cm) with an important mass of the pulp (Boukary et al., 2010).
The largest and most important component of ebony fruit is found in its pulp. Ebony fruit pulp has several chemical constituents and consequently many dietetic attributes (Kalinganire et al., 2007; Vinceti et al., 2013). Presently, the product is not fully utilized. It is usually considered as a waste by product of juice making. Once promoted, ebony pulp juice could be improved through selection that may significantly elevate the life of many rural masses.
Thus, quantitative traits require classical breeding to achieve high genetic gains (Voss-Fels et al., 2019). The diversity of fruit sizes found in this study has unveiled the high polymorphism existing in ebonies populations. Generally all the populations possess fruit sizes that are of economic value and can be used for domestication purposes (Juma et al., 2020). However, further research is required to assess the frequencies of different fruit types between and within provenances.
In general, sale of fruits is based on size (weight, length, width) (Yimer, 2015); bigger fruits fetch higher prices (field observation).Tree breeding may target trees with bigger fruits. However, there seems to be no relationship between the taste and size of the fruit which complicates the selection criteria. It has been reported by Katsvanga et al. (2007) that high fruit diversity attributes among sites could be attributed to climatic, edaphic, genetic and cultural factors. In the case of our sampling sites, there were huge differences in environmental factors that may be linked to differences in fruit weight related parameters observed. Domestication process involves moving genotypes from one site to another. Presently, it is not known how genotypes would respond once planted in an exotic habitat.
Here we used quantitative approach to set in evidence a relation between morphological structure according to the considered climatic zone and habitats. Several shapes of ebony tree organs (leaves and fruits) have been identified through climatic zone and habitats in the study area. They have been regrouped in four big groups whose measurements of a fruit diameter and length vary from one group of size to the other. Nearly all the forms of ebony fruits described by Wallnöfer (2001) and Arbonnier (2002) such as globose, bluntly ovoid, sharply ovoid, large ovoid, shallow sulcate and oblong-cylindrical exist within the nation. This implies rich diversity that does not require an infusion of external genetic material for immediate domestication and genetic improvement programs. The patterns of fruit sizes presently found cannot be used to specifically classify populations as ecotypes due to overlapping in multiple comparison tests. Will the fruit quality (shape, size, seed number, pulp color, nutrition and weight) be consistent when seed is moved from one ecological zone to another? In this case, provenance and family evaluation are prerequisite for successful large domestication programs. Some of the questions to answer in an evaluation program will be whether there are relationships between altitude, latitude, rainfall, temperature, relative humidity, stress period and edaphic factors on the fruit attributes (Katsvanga et al., 2007).
Overall morphological variation in D. mespiliformis
Without showing any present geographical trend (except altitudinal range for Atacora chains’ population), our find indicates that ebony morphological traits are influenced by climatic zones and habitats (environmental factors). In addition, a substantial part positive correlation was observed between the diameter and height of D. mespiliformis and tree organ dimensions. Studies of the morphological variation of D. mespiliformis were carried out on 701 natural individuals spread throughout Benin country located in West Africa. The morphological traits studied revealed relatively high variation in bearing and architectural parameters on the one hand and in tree descriptive organ on the other hand. This indicates that a fairly significant level of traits exchange occurs between populations individuals from different habitats and that a significant part of traits diversity of the species is of intra-population origin. The matrices of means distances among parameters show values that are very high (comprising between 0.32 and 0.95), which indicates that the populations display a morphological dissimilarity and leads to the assumption that they belong to several group (to be confirmed with genetic/molecular analysis). This level of diversity and this genetic structure might essentially be due to the difference in habitats/climatic zones of the species, without any bottleneck or slowing down of the biology of reproduction (incidence of synchronism and increased rate of fertilization) and the actions of man who has developed the parks of African ebony tree over time and helped the gene flow between them. This suggests that although D. mespiliformis prefer inundated forests because of permanence of water, individuals with plant parts of high dimensions are not found in this favourable (hospitable) habitat. Since D. mespiliformis is a plant with high light demanding, absence of high light in inundated forests explained in part low dimensions of tree organs.
CONCLUSION
Results showed that study area localities sheltered individuals of interested phenotypes (qualities) with well geo-referenced plus trees. This study highlights that adaptation of the species to environmental and geographical variation produce different morphological characteristics within its leaves and fruits. The significant difference in provenance (climatic zones and habitats) observed provided opportunities to select and conserve interesting materials from these locations. As findings from this more in-depth study, D. mespiliformis plant resources is used for a variety of purposes such as food, wood, charcoal, furniture, housing material, and medicine to mention just a few, the plant resource must be domesticate for its sustainable conservation. This research is the first report to assess the diversity within and among D. mespiliformis species in Benin using morphological descriptors. Our results should be considered for plant breeding and genetic resource conservation programs. Further investigations must be undertaken in order to determine the biochemical characteristics of each species morphotypes.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEGEMENTS
The authors appreciates the Federal Ministry of Education and Research (BMBF) and West African Science Service Centre on Climate Change and Adapted Land Use (WASCAL) for providing partial scholarship in form of Thesis Grant.
REFERENCES
|
Abasse T, Weber JC, Katkore B, Boureima M, Larwanou M, Kalinganire A (2011). Morphological variation in Balanites aegyptiaca fruits and seeds within and among parkland agroforests in Eastern Niger. Agroforestry Systems 81(1):57-66. |
|
|
Adomou (2005). "Vegetation patterns and environmentalgradients in benin Implications for biogeography and conservation. Wageningen University and Research. |
|
|
Akoegninou A, Van der Burg WJ, Van der Maesen LJG (2006). Analytical flora of Benin (No. 06.2). Backhuis Publishers. |
|
|
Arbonnier (2002). Arbres, Arbustes et lianes des zones sèches d'Afrique de l'Ouest: 565. |
|
|
Assogbadjo AE, Patrick BS, VAN DAMME (2005). Caractères morphologiques et production des capsules de baobab (Adansonia digitata L.) au Bénin. EDP Sciences 60:14. |
|
|
Boukary OD, Hélène IJ, Doyle Mc Key, Martine Hossaert-Mckey, Marie HC (2010). Variation des caractères biométriques des graines et des plantules de neuf provenances de Tamarindus indica L. (Caesalpinioideae). EDP Sciences 65(3):153-167. |
|
|
Catarino L, Romeiras MM, Bancessi Q, Duarte D, Faria D, Monteiro F, Moldão M (2019). Edible Leafy Vegetables from West Africa (Guinea-Bissau): Consumption, Trade and Food Potential, mdpi.com |
|
|
Dadegnon S, Gbemavo C, Ouinsavi C, Sokpon N (2015). Morphological Variation and Ecological Structure of Chrysophyllum albidum G.Don. International Journal of Plant and Soil Science 5(1):25-39. |
|
|
Dicko (2016). Influence des pressions anthropiques sur la structure des populationsde Pentadesma butyracea au Bénin. Vetigo[VertigO] La Revue Electronique en Sciences De L'environnement 16(3):22. |
|
|
Dicko A, Natta AK, Biaou HSS, Akossou A (2019). Assessing Morphological Traits Variation and Fruit Production of Lophira lanceolata (Ochnaceae) in Benin. |
|
|
Gbemavo CJ, Gandji K, Gnangle CP, Assogbadjo AE, Glèlè Kakaï RL (2015). Variabilité morphologiques et conservation des morphotypes de Jatropha curcas Linn. (Euphoriaceae) au Bénin. Journal of Agriculture and Environment for International Development 109(1):55-69. |
|
|
Gnonlonfin L, Ouinsavi C, Gouwakinnou G, Fandohan B, Houetchegnon TO (2018). Traditional knowledge and cultural importance of Diospyros mespiliformis in Northern Benin. Proceedings of 6th NSCB Biodiversity Conference; Uniuyo 2018 (200 - 207pp). Available at: |
|
|
INSAE (2015). RGPH4: Que retenir des effectifsde population en 2013?. Available at |
|
|
IPGRI (2016). Descripteurs de la tomate (Lycopersicon spp.), IPGRI. |
|
|
Juma I, Nyomora A, Hovmalm HP, Fatih M, Geleta M, Anders SC, Ortiz RO (2020). Characterization of Tanzanian Avocado Using Morphological Traits. Diversity MDPI 12(64):22. |
|
|
Kalinganire A, Weber JC, Uwamaria A, Kone B. (2007). "Improving Rural Livelihoods through Domestication of Indigenous Fruit Trees in the Parklands of the Sahel. Fruit trees 10, 186-203. |
|
|
Katsvanga CTA, Jim L, Gwenzi D, Muhoni L, Masuka P, Moyo M (2007). Characterisation of community identified uapaca kirkiana phenotypes for domestication. Journal of Sustainable Development in Africa 9(4):11. |
|
|
Kimpouni V, Mamboueni JC, Tsoungould GFM, Mikoko EN (2019). Environment and livelihood of the Kouni community of the Kayes sub-prefecture (Bouenza, Congo). Ethnobotany Research and Applications 18:1-15. |
|
|
Kouyaté AM (2005). Aspects ethnobotaniques et etude de la variabilite morphologique, biochimique et phenologique de detarium microcarpum guill. & perr. au mali. Sciences Ingénieurs Section Agronomie, Gent University. Doctor. |
|
|
Maroyi A (2018a). Diospyros lycioides Desf.: Review of its botany, medicinal uses, pharmacological activities and phytochemistry, Asian Pacific Journal of Tropical Biomedicine 8(2):130. |
|
|
Maroyi A (2018b). Euclea crispa: Review of its botany, ethnomedicinal uses, and pharmacological properties. |
|
|
Mkwezalamba I, Munthali CR, Missanjo E (2015). Phenotypic Variation in Fruit Morphology among Provenances of Sclerocarya birrea (A. Rich.) Hochst. International Journal of Forestry Research, 2015. |
|
|
Okpanachi O, Aguoru CU, Azua ET, Olasan JO (2019). Biodiversity and species distribution of indigenous under-utilized fruit and vegetable crops in the north Senatorial District of Nasarawa State, Nigeria. International Journal of Agricultural Research and Food Production 4(3). |
|
|
Olajide (2019). "Data Collection with KoboToolbox." |
|
|
Ouinsavi C, Sokpon N (2010). Morphological Variation and Ecological Structure of Iroko (Milicia excelsa Welw. C.C. Berg) Populations across Different Biogeographical Zones in Benin. International Journal of Forestry Research vol. 2010, Article ID 658396 10 p. |
|
|
Romeiras MM, Duarte MC, Francisco-Ortega J, Catarino L, Havik P (2018). "Recovering Plant Data for Guinea-Bissau: Implications for Biodiversity Knowledge of West Africa." Diversity 10(4):109. |
|
|
Tiokeng B, Zapfack L, Nguetsop VF, Saha Z, Nchongboh GC (2019). Sacred Forests in the Western Highlands-Cameroon: Ethnobotany Role and Indigenous Conservation of Biodiversity Advance research journal of multidisciplinary discoveries 35(1):54-59. |
|
|
Vinceti B, Termote C, Ickowitz, A, Powell B, Kehlenbeck K, Hunter D (2013). The contribution of forests and trees to sustainable diets. Sustainability 5(11):4797-4824. |
|
|
Voss-Fels KP, Cooper M, Hayes BJ (2019). Accelerating crop genetic gains with genomic selection. Theoretical and Applied Genetics 132(3):669-686. |
|
|
Wade TI, Ndiaye O, Mauclaire M, Mbaye B, Sagna M, Guissé A, Goffner D (2018). Biodiversity field trials to inform reforestation and natural resource management strategies along the African Great Green Wall in Senegal. New Forests 49(3):341-362. |
|
|
Wallnöfer B (2001). The biology and systematics of Ebenaceae: a review. Annalen des Naturhistorischen Museums in Wien. Serie B für Botanik und Zoologie pp. 485-512. |
|
|
Yimer A (2015). Factors Affecting Fruit Supply in the Market: The Case of Habru Woerda, North Wollo, Ethiopia. European Journal of Business and Management 7(4):12. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


