Full Length Research Paper

ABSTRACT
Experiments were conducted at Nyambaka in the Adamawa Region of Cameroon, from March 2017 to November 2018, for assessing the apicultural potential of Bidens steppia, Cordia africana, Pittosporum viridiflorum and Psychotria mahonii. In order to improve the beekeeping productivity, it is important to investigate the diversity of bee plants in a given apiary site. To that end, the foraging behaviour of Apis mellifera workers was studied on the flowers of each plant species twice a week during the whole blooming period. The abundance of opened flowers per plant, floral products harvested by foragers, mean duration of floral visit, mean density of foragers, sugar content of nectar of each plant species, and number of effective visits of A. mellifera on flowers were assessed. Results indicated that, honeybee workers harvested nectar of each plant species; B. steppia and P. viridiflorum were intensely foraged for pollen harvesting too. The abundance of workers/1000 flowers varied from 123 on P. mahonii to 724 on C. africana. The mean value of the sugar content of nectar oscillated from 19.50% (C. africana) to 38% (B. steppia). C. africana and P. mahonii were highly nectariferous plant species while B. steppia was very highly polliniferous and slightly nectariferous and P. viridiflorum very highly nectariferous and slightly polliniferous. During its foraging activity, workers improved the pollination possibilities of plant species. By planting or protecting these plant species, a bee-friendly garden can be preserve for providing nectar flow and pollen availability for beekeeping purpose.
Key words: Apiary, apicultural value, beekeeping, bee plant, foraging behaviour, sugar content.
INTRODUCTION
Beekeeping is one of the most important cultural and economic activities in the Adamawa region of Cameroon (Ingram, 2011). The highest quantity of honey consumed or marketed in this country is from this region which has a suitable climatic condition for the proliferation of honeybees (INADES, 2000). Despite the favorable agro-ecology of honey production and the high number of bee colonies this region is endowed with, the honey production and productivity level in Cameroon is still very low (Dongock et al., 2017). Yet, sustainable beekeeping can be improved through the understanding and conservation of plants producing nutrient for the honeybees mainly in terms of nectar and pollen (El-Nebir and Talaat, 2013).
Apis mellifera is one of the bees raised on a large scale in beekeeping to produce honey and for pollination. This species comprises 28 subspecies including Apis mellifera adansonii, which has its origin from Africa (Fletcher, 1978). Honeybee workers are attracted commonly to plants that produce nectar and pollen. Nectar is a sweet substance that attracts bees which also need pollen in their diet (Louveaux, 1984). These food sources provide the nutritional requirements of the bee colonies. Nectar as a source of honey provides heat and energy while pollen provides protein, vitamins and fatty substances (Amsalu et al., 2003). During floral visits for nectar harvesting and pollen gathering, honeybees, in turn, pollinate plants; thus they can help in boosting fruit and seed yields of the host plant and then propagate their species (Klein et al., 2007; Allsopp et al., 2008).
The honey and other products of honeybees depend on the availability of floral resources in a given area (Amsalu et al., 2003). Intending to improve the level of honey production both in quantity and quality in the Adamawa region of Cameroon, several findings are available in enhancing beekeeping practice regarding the inventory of bee plants in some sites like Ngaoundere (Tchuenguem et al., 2010; Ingram, 2011; Djonwangwé et al., 2011; Egono et al., 2018; Wékéré et al., 2018) and Ngaoundal (Dongock et al., 2017).
Nyambaka is a small locality in the Adamawa region of Cameroon where beekeeping is still done on a small-scale. In this area, beekeeping practice appears like a commercial enterprise; it offers not only diverse hive products which can be sold in local markets and become an important source of regular income for farmer families, but also provide complementary services, such as plant pollination. Moreover, locally, bee products improve farm family nutrition and can provide traditional health care remedies. There are many plant species that produce a large amount of nectar and pollen for bees to be collected in the locality. Some of these plants are important as they provide bees with a surplus of honey. Small-scale beekeeping is considered as an important occupation that contributes significantly to livelihood security in that region. Yet, most of the honey is produced with traditional hives consisting of bast and grass; besides, bee plants are not yet well known by beekeepers in this area. It is, therefore, an important practice to help bees in their survival by adding to the shrinking inventory of flower-rich habitat in the study locality.
The United Nations World Health Organization estimates that as many as 5.6 billion people, 80% of the world population, utilize herbal medicine for primary health care (Shen et al., 2012). Bidens steppia (Steetz) Sherff (Asteraceae), Cordia africana Lam. (Boraginaceae), Pittosporum viridiflorum Sims (Pittosporaceae), and Psychotria mahonii C.H. Wright (Rubiaceae) are four multipurpose plant species which are often harvested for local use as food and medicine in Nyambaka. In the locality, different preparations of parts of B. steppia plant are commonly purported to treat several categories of illnesses such as diabetes and malaria. In the literature, extracts of B. steppia have antitumor (Sundararajan et al., 2006), anti-inflammatory, antimicrobial (Pereira et al., 1999), antidiabetic, antimalarial (Tobinaga et al., 2009) properties. C. africana is used as firewood. The fruit pulp of this plant is edible and is added as a sweetener to food. The leaves serve as fodder for livestock. Leaf decoctions are administered to treat headache, nose bleeding, dizziness and vomiting during pregnancy, wounds and worms while root decoctions are drunk to treat jaundice (Obeng, 2010). The categories to which P. viridiflorum is used in traditional medicine include wounds, treatment of veterinary ailments, gastrointestinal tract and sexually transmitted diseases, circulatory and inflammatory disorders, as well as diseases such as cancer, tuberculosis, and malaria (Madikizela and McGaw, 2017). As for P. mahonii, fresh rhizomes are chewed and the juice swallowed to treat intestinal tract diseases and worms.
In addition to their medicinal importance, all four plant species studied have flowers that produce nectar and pollen available for bee species. It is well known in the literature that the productivity of the honeybee colonies is proportional both to the abundance and attractiveness of the nectariferous and polliniferous plants present in the environment of the apiary (Williams and Carreck, 1994; Van’t et al., 2005). Thus, sustainable beekeeping in a given region requires detailed knowledge of the apicultural value of plant species that grow there for their optimal management (Dongock et al., 2017). Moreover, honeybee being a bio-indicator species (Porrini et al., 2003), medicinal plants are an interesting source for the production of honey with medicinal biological activity very close to their floral origin (Liberato et al., 2011). That is why it is interesting to associate a crop of medicinal plants with beekeeping. However, there is no available data published on the relationships between African honeybees and many local and medicinal plant species including B. steppia, C. africana, P. viridiflorum and P. mahonii in Cameroon. The main objective of the present research work was to determine the apicultural status of these plant species in Nyambaka, to improve the beekeeping potential and enhance the value of the plants studied in this locality. For each plant species, we recorded the foraging activity of A. mellifera on flowers and estimated its apicultural value.
MATERIALS AND METHODS
Study site and biological material
The present study was carried out from March 2017 to November 2018 in Nyambaka, a village located in the South of Ngaoundere, the capital of the Adamawa region of Cameroon. This region is located between the 6th and 8th degrees of latitude north and between the 11th and 15th degrees of longitude east; it belongs to the high-altitude Guinean Savannah agro-ecological zone (Djoufack et al., 2012). The climate is characterized by a rainy season (April to October) and a dry season (November to March), with an annual rainfall of approximately 1500 mm, a mean annual temperature of 22°C, and a mean annual relative humidity of 70% (Amougou et al., 2015) which are suitable climatic conditions for the beekeeping practice. Plants chosen for observations were located in an area of 1.5 km in diameter, centered on a Kenyan top-bar hive of A. mellifera colony. The hive was located at the following coordinates: 6°89’62’943’N, 14°09’28’038’E, 1136 m a.s.l.
The animal material included many insect species naturally present in the environment. The number of honeybee colonies located in the area varied from 60 in March 2017 to 69 in November 2017 and from 42 in March 2018 to 72 in November 2018. Apart from the honeybee colonies located in the experimental site, other colonies around the experimental site have not been inventoried since the radius action of foragers may exceed 12 km around the hive (Louveaux, 1984). The vegetation was represented in the study site by crops, ornamental plants, hedge plants and native plants of savannah and gallery forests.
Study of the foraging activity of A. mellifera on flowers
From March 2017 to November 2018, the foraging behaviour of A. mellifera workers was recorded on flowers of different plant species. Data were registered twice a week, between 07:00 a.m. and 06:00 p.m., at three-time intervals per day: 07:00-11:00 a.m., 11:00 a.m.-03:00 pm and 03:00-06:00 p.m. For any plant species visited by the honeybee browsers and for each investigation date, the following parameters were registered for each time frame and, whenever possible: the number of effective visits (the bee came into contact with the stigma) (Jacob-Remacle, 1989; Freitas, 1997), the mean duration of visits using a stopwatch (Tchuenguem et al., 2004). The density of foragers (highest number of individuals foraging simultaneously on a flower or 1000 flowers) was also assessed. The density of foragers was recorded following the direct counting on the same dates and daily periods as for the registration of the duration of individual flower visits; for this purpose, some foragers were counted on a known number of flowers. The density of foragers per 1000 flowers (A1000) was then calculated using the following formula: A1000 = ((Ax/Fx)*1000) where Fx and Ax are the numbers of opened flowers and the number of foragers effectively counted on these flowers at a given time x (Tchuenguem et al., 2004). The disruption of the activity of foragers by competitors and the attractiveness exerted by other plant species on A. mellifera was assessed by direct observations.
Evaluation of the sugar content of the nectar of different plant species
The concentration in total sugar of the nectar is an important parameter for the attractiveness of the honeybee concerning many flowers (Philippe, 1991). This parameter was determined with a handheld refractometer (0-90% Brix) and a thermometer that gave the ambient temperature, from March 2017 to November 2018. A. mellifera workers in full activity of nectar harvesting were captured on flowers and anesthetized by introducing them in a small jar containing cotton moistened with chloroform. The nectar was then removed from honeybee crop by exerting pressure on the bee abdomen placed between the thumb and the forefinger of the experiment; the nectar in the mouth was then expelled and its concentration in total sugars measured in g/100 dry matter (Tchuenguem et al., 2007). The registered values obtained were corrected according to the ambient temperature, using a table provided by the device leaflet (Cruden and Hermann, 1983).
Evaluation of the apicultural value of different plant species
As for other plant species, the apicultural value of each plant species studied was assessed using data on the flowering intensity, the degree of attractiveness of A. mellifera workers to nectar and/or pollen (Villières, 1987; Népidé et al., 2016).
Evaluation of the influence of Apis mellifera on pollination
To evaluate the ability of A. mellifera to act as a pollinator of each plant species during the nectar harvesting, the number of time a forager came into contact with the stigma of the visited flower was noted (Freitas, 1997). This approach allows highlighting the involvement of A. mellifera in self-pollination and cross-pollination (Zumba et al., 2013; Potts et al., 2015).
Statistical analysis
Data were subjected to descriptive statistics using SPSS 16. The Student’s t-test was used for the comparison of means between two samples and Chi-square (χ2) for the comparison of two percentages. The analysis of variance (ANOVA) was used for multi-comparison of means.
RESULTS
Characteristics of the plant species studied
Tables 1 and 2 described plant species studied and the relative abundance of opened flowers per month for each of these plant species during both observation years. It is indicated from these tables that apart from C. africana which is mainly cultivated in the locality for the fruit production and shady purposes, B. steppia, P. viridiflorum and P. mahonii are respectively a shrub and small trees which grow spontaneously in the savannah.
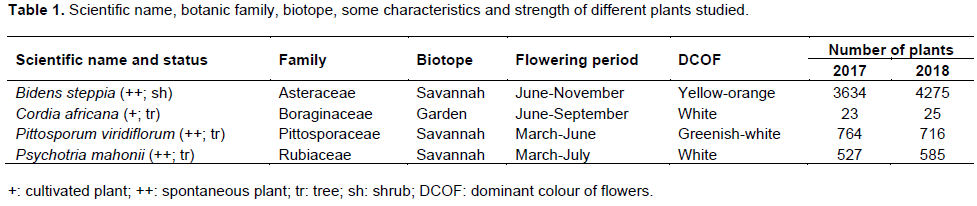
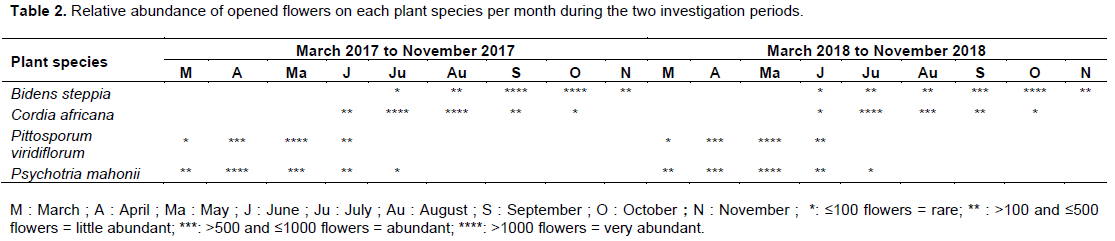
The color of the flowers of these plant species varied from orange-yellow, greenish-white and white respectively for B. steppia, P. viridiflorum, C. africana and P. mahonii. Furthermore, the number of these plant species varied from about 25 for C. africana to about 4275 for B. steppia. These important numbers of plant species in the studied area enabled the availability of a large amount of floral mass during their blossoming.
A. mellifera foraging activity on flowers
Floral products harvested, intensity and frequency of collection of different products
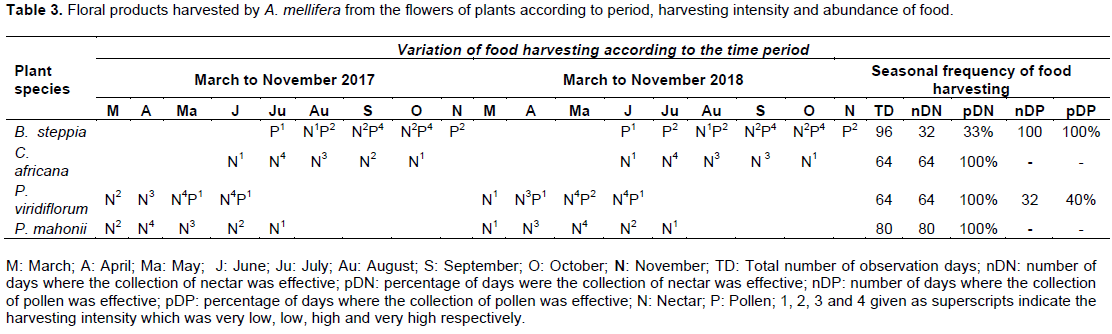
The identity of the food harvested by A. mellifera workers from the flowers of each plant species investigated and the intensity and frequency of the collection of different food resources are presented in Table 3. The main results from this table indicated that: (a) B. steppia was weakly visited for nectar gathering while its pollen was highly attractive for A. mellifera workers (b) P. viridiflorum was slightly visited for its pollen while its nectar was abundantly harvested by honeybee foragers; (c) C. africana and P. mahonii were only visited for supplying nectar needs of A. mellifera as their pollen was scarcely collected. In general, the intensity of nectar or pollen collection varied with plant species and for a given plant species with the time.
The distribution of nutritive substances harvested by A. mellifera on flowers in terms of nectar and/or pollen of a given plant species according to each observation time interval is reported in Table 4. A. mellifera workers foraged nectar and/or pollen of B. steppia, C. africana, P. viridiflorum and P. mahonii almost during the whole daily period, from 06:00 a.m. till 06:00 p.m. and during all the blooming period of each plant species studied. This is an illustration that these plant species are important and abundant sources of nutrients for A. mellifera.

Density of foragers
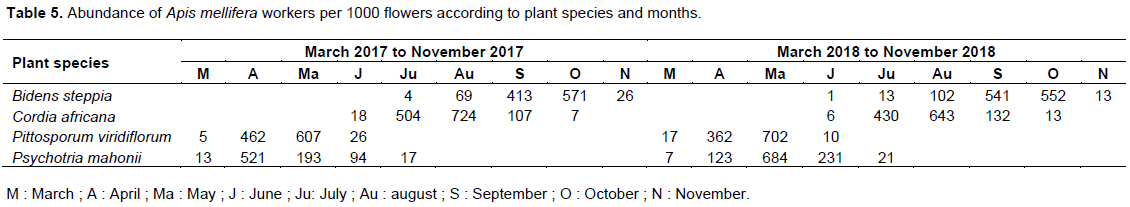
The values of the density of foragers are reported in Table 5. The highest number of A. mellifera workers foraging simultaneously per flower was one for each plant species. The mean abundance per 1000 flowers in 2017 was 216 (n = 240; s = 122; maximum = 517) on B. steppia, 272 (n = 197; s = 143; maximum = 727) on C. africana, 271 (n = 146; s = 131; maximum = 643) on P. viridiflorum and 167 (n = 205; s = 94; maximum = 727) on P. mahonii. In 2018, the corresponding figures was 203 (n = 268; s = 118; maximum = 552) on B. steppia, 244 (n = 217; s = 128; maximum = 643) on C. africana, 272 (n = 116; s = 142; maximum = 702) on P. viridiflorum and 213 (n = 189; s = 121; maximum = 684) on P. mahonii. The optimal value of the density of A. mellifera workers corresponded with the month of the peak of blossoming of each plant species studied which are: September and October for B. steppia, July to August for C. africana, April and May for P. viridiflorum and P. mahonii respectively. The difference of the mean density of foragers was not significant for each plant species according to the year.
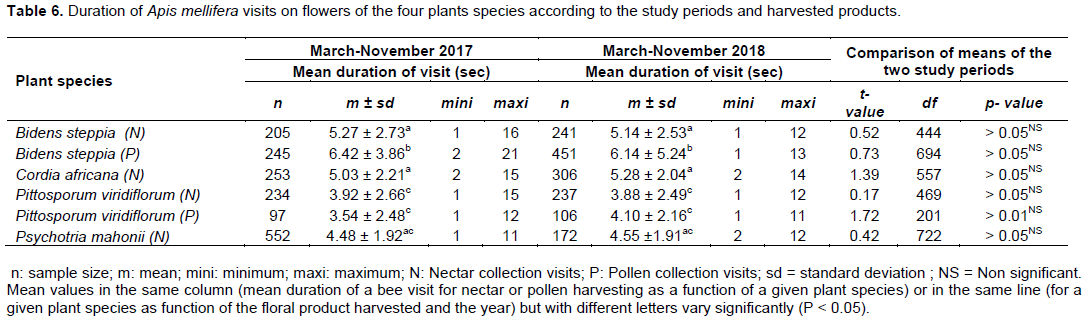
Duration of visits
Results from Table 6 highlighted the variation of the mean duration of A. mellifera visit as a function of the floral product harvested and for a given plant species according to the year. As pollen grains of C. africana and P. mahonii were not so interesting for foragers, only the mean duration of a visit for pollen collection by A. mellifera on B. steppia and P. viridiflorum were registered in both years. Overall, the duration of a forager visit varied with the floral product searched for and for a given product with the host plant species studied. The difference was significant between pollen and nectar gathering on B. steppia in 2017 (t = 5.23; df = 448; P < 0.05) and 2018 (t = 3.97; df = 690; P < 0.05), between nectar collection among plant species in 2017 (F = 6.52; df = 3, 1240; P < 0.05) and 2018 (F =5.93; df = 3, 952; P < 0.05) and between pollen collection on B. steppia and P. viridiflorum in 2017 (t = 7.63; df = 340; P < 0.05) and 2018 (t = 6.57; df = 555; P < 0.05). Workers of A. mellifera were disturbed during their foraging activity by other flower-visiting insects or abiotic parameters such as the wind and the rainfall. Some disturbances have resulted in the interruption of some honeybee visits and consequently reduced the time spend on the corresponding flower, thus obliged foragers to move swiftly from flower to flower.
Influence of neighboring flora
During the observation periods of each of the four plant species under investigation, flowers of many other plant species growing in the study area were visited by A. mellifera for nectar (ne) and/or pollen (po). Among these plants were, Manihot esculenta (Euphorbiaceae: ne), Tithonia diversifolia (Asteraceae: ne), Bidens pilosa (Asteraceae: ne + po), Stylosanthes guianensis (Fabaceae: ne + po), Hibiscus rosa-sinensis (Malvaceae: ne + po), Sida rhombifolia (Malvaceae: ne + po), Terminalia schimperiana (Combretaceae: ne + po), Terminalia macroptera (Combretaceae: ne + po), Sesbania pachycarpa (Fabaceae : ne + po), Mimosa invisa (Mimosaceae: po), Mimosa pudica (Mimosaceae: po), Senna mimosoides (Fabaceae: po), and Zea mays (Poaceae: po). During one foraging trip, a forager was not observed moving from the flowers of a given plant species studied to the neighboring plant species and conversely.
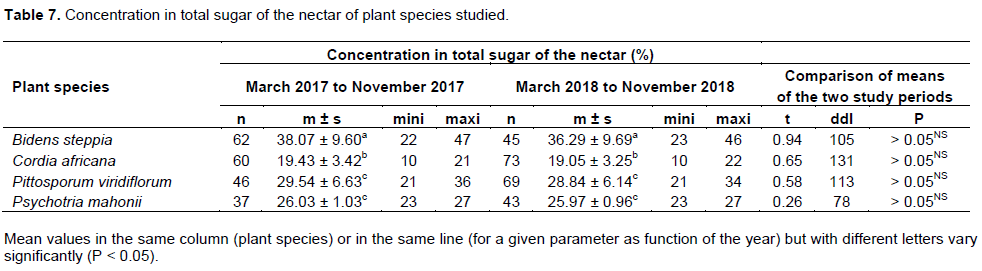
Concentration in total sugar of the nectar of plant species studied
Results from Table 7 reported the mean concentration in total sugar of B. steppia nectar was 38.07% (n = 62; s = 9.60) in 2017 and 36.29% (n = 45; s = 9.69) in 2018. The difference between these means is not significant (t = 0.94; df = 105; P > 0.05). The mean concentration in total sugar of the C. africana nectar was 19.43% (n = 60; s = 3.42) in 2017 and 19.05% (n = 73; s = 3.25) in 2018. The difference between these two means is not significant (t = 0.65; df = 131; P > 0.05). The mean concentration in total sugars of P. viridiflorum nectar was 29.54% (n = 46; s = 6.63) in 2017 and 28.84% (n = 69; s = 6.14) in 2018. The difference between these means is not significant (t = 0.58; df = 113; P > 0.05). The mean concentration in total sugar of P. mahonii nectar was 26.03% (n = 37; s = 1.03) in 2017 and 25.97% (n = 43; s = 0.96) in 2018. The difference between these means is not significant (t = 0.26; df = 78; P > 0.05). The difference was significant between the four plant species concerning this parameter in 2017 (F = 7.14; df = 3, 201; P < 0.05) and in 2018 (F =5.31; df = 3, 226; P < 0.05). Overall, the concentration in total sugar of the nectar varies following plant species.
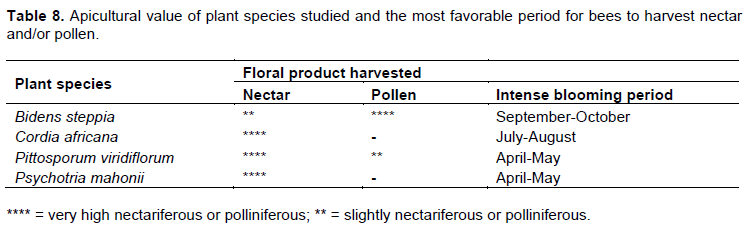
Apicultural value of the plant species
During the flowering period of each plant species studied, we recorded distinct levels of activity of A. mellifera workers on the flowers. There were a high availability of flowers, a high density of workers per tree or shrub, a good nectar collection on all plant species and high pollen harvest on B. steppia but a low pollen collection on P. viridiflorum. Moreover, in the dry season, which is the main period of honey flow, individual tree of P. viridiflorum and P. mahonii investigated could produce more than 70.000 flowers. On the other hand, during the rainy season (period of the food shortage within honeybee colonies), individual tree of B. steppia and C. africana could averagely produce up to 15.000 flowers. Considering these data, the plant species studied can be classified based on their apicultural value as follows: (a) very highly nectariferous: C. africana, and P. mahonii; (b) very highly polliniferous and slightly nectariferous: B. steppia (c) very highly nectariferous and slightly polliniferous: P. viridiflorum. Table 8 summarizes the appropriate period for the optimal nectar and/or pollen availability in Nyambaka in 2017 and 2018. Thus, in this study area, the nutritional requirements of honeybees are provided by B. steppia from August to September, C. africana from June to August while P. viridiflorum and P. mahonii were important suppliers of nectar to foragers from April to May.
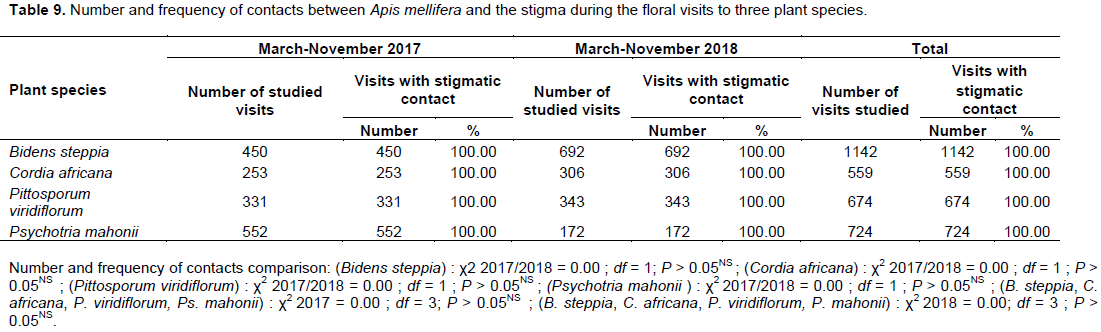
Impact of A. mellifera activity on the pollination of the plant species
During the collection of pollen or nectar on the flowers of the four plant species studied, foragers regularly contacted anthers and carried pollen. With this pollen, they flew frequently from flower to flower. The percentage of the total number of visits during which worker bees came into contact with the stigma of the visited flower was 100% for B. steppia, C. africana P. viridiflorum and P. mahonii during the 2017 as well as the 2018 investigation period (Table 9). Consequently, A. mellifera workers strongly increase the pollination possibilities of B. steppia, C. africana, P. viridiflorum and P. mahonii.
DISCUSSION
In Nyambaka, an important area for beekeeping practice in the Adamawa region of Cameroon, the four plant species studied have shown their importance as sources of pollen and nectar for African honeybees. Previous works have already been done in other countries which have shown the immense potential that some of these plant species abounded in the local beekeeping practice. That is the case for C. africana which was mentioned in the Western Amhara region of Ethiopia as a major honeybee plant (Tesfa et al., 2013); moreover, the pollen of this plant species was identified as the secondary pollen source in four honey samples collected from Salika village in Ethiopia (Tewelde, 2006). The collection of the nectar of P. psychotroides by A. mellifera, another plant species belonging to the genus Psychotria, has already been reported in previous investigations in the Adamawa Region (Dongock et al., 2017). In Benin, Yédomonhan et al. (2009) found that, on the flowers of Psychotria spp., A. mellifera harvested both pollen and nectar widely. In the southern highland of Tanzania, the flowers of B. steppia and P. viridiflorum were mentioned as an important source for both pollen and nectar collection all day long and during the whole flowering period (Latham, 2015). In Bujumbura (Burundi), B. steppia flowers were mainly visited by the honeybee (Ndayikeza et al., 2014) for both nectar and pollen collection. Overall, the substance harvested by A. mellifera from flowers (nectar or pollen) on a given plant species can vary with regions. The variations observed in this study could be explained mainly by the real needs of the colonies from which originated honeybee workers (Segeren et al., 1996). The good nectar collection observed in the four plant species is the fact that, bees can collect nectar with sugar concentrations below 15-85% under natural conditions (Roubik and Buchmann, 1984).
From our fieldwork, the colors of the flowers of the studied plant species are among the most attractive for honeybee foragers. According to Bergström (1982), the bright colors are the most attractive for the flower-visiting insects in general and honeybees in particular because those are among the colors they can easily perceive. Indeed, bees have a much broader range of color vision. Their ability to see ultraviolet light gives them an advantage when seeking nectar and pollen (Dyer and Garcia, 2014). Thus, to identify the most profitable flowers, and avoid non-rewarding flowers like mimics (Dafni, 1984), bees make use of several cues including color (Hempel et al., 2001) information. Furthermore, several other parameters such as the accessibility of nectar and/or pollen, the availability of both products, the floral mass or number of flowers or inflorescences bearded by a plant species are important for the good practice of beekeeping elsewhere (Segeren et al., 1996).
Another important aspect from the present study is the possibility of the mastery of the period when the blossoming of each plant species studied is effective and optimal yearly. Indeed, the knowledge of these time intervals makes it possible to establish the apicultural calendar in avoiding pollen and nectar scarcity and shortage (Chigere et al., 2014). For this purpose, the supply of A. mellifera foragers with nectar and/or pollen in a well-known period of the year is already possible for B. steppia, C. africana, P. viridiflorum and P. mahonii in the study site.
The observed high abundance of foragers per 1000 flowers recorded in this study could be attributed to the ability of honeybees to recruit a great number of workers for the exploitation of high-yield food sources (Frisch, 1969; Kajobe, 2006). Honeybees can smell or detect pollen or nectar odor (Free, 1970) using sensory receptors located on the flagellum of their antennae. Worker honeybees dance inside the nest after a successful foraging trip to communicate with their congeners, information about the food odor, the distance and the direction from the hive to the food source (Frisch, 1967). The round dance is performed when the resource is within 100 meters from the hive, while the wagging dance takes place for the resource 100 meters away from the hive (Frisch, 1967).
Signiï¬cant differences observed between the duration of pollen harvesting visit and that of nectar harvesting on the flowers of B. steppia could be explained by the accessibility of each of these floral products. On the flower of B. steppia, pollen is a product in great quantity and is easily accessible to bees. In these conditions, honey bee workers can obtain their pollen load by visiting a few flowers during a foraging trip. That is why A. mellifera spent more time on a flower for pollen harvesting than for nectar in B. steppia. Therefore, on each of the four plant species, A. mellifera spent more time on a flower for nectar collection of C. africana, B. steppia, P. mahonii and P. viridiflorum respectively. The fact that A. mellifera spent significantly different time on a flower of different species for nectar and pollen collection could be explained by the abundance and/or the accessibility to each of these floral products.
The disruptions of visits by other insects reduced the duration of certain A. mellifera visits. This obliged some workers to visit more flowers during a foraging trip, to maximize their pollen or nectar loads. Similar observations were made for A. mellifera workers foraging on flowers of Entada africana (Fabaceae), Eucalyptus camaldulensis (Myrtaceae), Psidium guajava (Myrtaceae) and Trichillia emetica (Meliaceae) (Tchuenguem et al., 2007), Combretum nigricans (Combretaceae), Erythrina sigmoidea (Fabaceae), Lannea kerstingii (Anacardiaceae), Vernonia amygdalina (Asteraceae) (Tchuenguem et al., 2010), Jatropha curcas (Euphorbiaceae), Senegalia polyacantha (Mimosaceae) and Terminalia schimperiana (Combretaceae) (Wékéré et al., 2018)
The present study shows that during one foraging trip, an individual bee foraging on a given plant species scarcely visited another plant species. This result indicates that A. mellifera shows flower constancy (Louveaux, 1984) for the flowers of each of the four plant species studied. This floral constancy in honeybees exists because an individual forager is generally capable of memorizing and recognizing the shape, color, and odor of the flowers visited during previous foraging trips (Hill et al., 1997; Wright et al., 2002). The flower constancy of A. mellifera has been demonstrated on flowers of several other plant species among which are Jatropha curcas (Euphorbiaceae), Senegalia polyacantha (Mimosaceae) and Terminalia schimperiana (Combretaceae) (Wékéré et al., 2018), Lannea kerstingii (Anacardiaceae) and Ximenia americana. (Olacaceae) (Djonwangwé et al., 2011), and Callistemon rigidus (Myrtaceae) (Fameni et al., 2012). The faithfulness of the honeybee to the flowers of the plant species studied can also be explained, in part, by the fact that their nectar is rich in sugars; Philippe (1991) suggested that foragers could not allow their colony to record a net energy gain if the sugar concentration of the harvested nectar is less than 20%.
Considering this minimum limit, browsers can allow their colony to gain a lot of energy when they collect nectar from plant species studied.
During the collection of nectar and/or pollen on each flower, A. mellifera workers regularly come into contact with the stigma and anthers. They could thus enhance self-pollination, which has been demonstrated for other plant species in the past (Anderson and Symon, 1988; Lewis et al., 1999; Otiobo et al., 2015). A. mellifera could induce cross-pollination through carrying of pollen with their furs, legs and mouth accessories, which is subsequently deposited on another flower belonging to different plants of the same species (Abrol, 2012).
CONCLUSION
At Nyambaka, A. mellifera workers harvested intensely and regularly the nectar and pollen of B. steppia, C. africana, P. viridiflorum and P. mahonii flowers. These results suggest that these plants are the highly nectariferous and polliniferous floral plant able to substantially contribute to maintaining the nutritional needs of the honeybee colony. All these plant species contributed more or less to the feeding and therefore to the strengthening of the honeybee colonies. A. mellifera workers increased the pollination possibilities of each plant species. Based on our results, we recommend: (a) the installation of A. mellifera colonies in environments where one or more of the studied plant species occur abundantly and (b) the plantation and/or protection of each plant species in the surrounding of A. mellifera apiaries. These precautions will allow, in addition to improving the production of honey in the study location, its enrichment in various therapeutic properties for the well-being of the local populations. The impact of A. mellifera on fruit or grain yields of each plant species studied via its pollination efficiency will be looked at in future works.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abrol DP (2012). Pollination biology: biodiversity conservation and agricultural production. Springer Dordrecht Heidelberg, London, UK 792 p. |
|
|
Allsopp MH, de Lange WJ, Veldtman R (2008). Valuing insect pollination services with cost of replacement. PLoS ONE 3(9):3128. |
|
|
Amougou JA, Abossolo SA, Tchindjang M (2015). Variability of precipitations at Koundja and Ngaoundere based on temperature changes of Atlantic Ocean and El NINO. Ivory Coast Review of Science and Technology 25:110-124. |
|
|
Amsalu B, Nuru A, Radloff H (2003). Multivariate morphometric analysis of honeybees in the Ethiopian region. Apidologie 35:71-81. |
|
|
Anderson GJ, Symon D (1988). Insect foragers on Solanum flowers in Australia. Annals of Missouri Botanic Garden 75(3):842-852. |
|
|
Bergström G (1982). Relations chimiques entre les Orchidées et les pollinisateurs. Bulletin de la Société Entomologique de France 90 (5-6 /7-8):1223-1228). |
|
|
Chigere M, Machingauta P, Chiororo L (2014). Threats to the African bees and beekeeping in Zimbabwe. HICC, Harare, Zimbabwe 15 p. |
|
|
Cruden RW, Hermann SM (1983). Studying nectar: Some observations on the art. In: Bentley & Elias Ed., The biology of nectarines. Columbia University press, New York pp. 223-242. |
|
|
Dafni A (1984). Mimicry and deception in pollination. Annual Review of Ecology and Systematics 15:259-278 |
|
|
Djonwangwé D, Tchuenguem FFN, Messi J (2011). Foraging and pollination activities of Apis mellifera adansonii Latreille (Hymenoptera: Apidae) on Ximenia americana (Olacaceae) flowers at Ngaoundéré (Cameroon). International Research Journal of Plant Sciences 2(6):170-178. |
|
|
Djoufack V, Fontaine B, Martiny N, Tsalefac M (2012). Climatic and demographic determinants of vegetation cover in northern Cameroon. International Journal of Remote Sensing 21:6904-6926. |
|
|
Dongock ND, Zra E, Tchuenguem FFN (2017). Bee plant potentials and characteristics in the Ngaoundal subdivision, Adamawa-Cameroon. Agricultural Science Research Journal 7(9):285-296. |
|
|
Egono NCC, Kingha TMB, Fameni ST, Dounia, Tchuenguem FFN (2018). Pollination efficiency of Apis mellifera (Hymenoptera: Apidae) on Helianthus annuus (Asteraceae) flowers at Dang (Ngaoundéré, Cameroon). International Journal of Biosciences 13(3):314-328. |
|
|
El-Nebir MA, Talaat DAM (2013). Identification of botanical origin and potential importance of vegetation types for honey production in the Sudan. Journal of Natural Resources and Environmental Studies 1(2):13-18. |
|
|
Fameni TS, Tchuenguem FFN, Brückner D (2012). Pollination efficiency of Apis mellifera adansonii (Hymenoptera: Apidae) on Callistemon rigidus (Myrtaceae) flowers at Dang (Ngaoundéré, Cameroon). International Journal of Tropical Insect Sciences 32(1):2-11. |
|
|
Fletcher DJC (1978). The African Bee, Apis mellifera adansonii, in Africa. Annual Review of Entomology 23 (1):151-171. |
|
|
Free JB (1970). Insect pollination of crops. Academic press, London, UK 506 p. |
|
|
Freitas BM (1997). Number and distribution of cashew (Anacardium occidentale) pollen grains on the bodies of its pollinators, Apis mellifera and Centris tarsata. Journal of Apicultural Research 36(1):15-22. |
|
|
Frisch KV (1967). The dance language and orientation of bees. The Belknap Press of Harvard University Press, Cambridge, UK 235 p. |
|
|
Frisch KV (1969). Vie et mœurs des abeilles. A. Michel (Ed.), Paris, France 556 p. |
|
|
Hempel DIN, Giurfa M, Vorobyev M (2001). Detection of colored patterns by honeybees through chromatic and achromatic cues. Journal of Comparative Physiology 187:215-224 |
|
|
Hill PM, Wells PH, Wells H (1997). Spontaneous flower constancy and learning in honey bees as a function of colour. Animal Behaviour 54(3):615-627. |
|
|
INADES (2000). Rapport des ateliers avec les Apiculteurs de l'Adamaoua, Cameroun. INADES-Formation, Antenne de Maroua (Ed.), Maroua, Cameroun 29 p. |
|
|
Ingram V (2011). Bee plants for Cameroon highlands and Adamawa plateau honey. Center for International Forestry Research (CIFOR) pp. 1-22. |
|
|
Jacob-Remacle A (1989). Comportement de butinage de l'abeille domestique et des abeilles sauvages dans des vergers de pommiers en Belgique. Apidologie 20:271-285. |
|
|
Kajobe R (2006). Pollen foraging by Apis mellifera and stingless bees Meliponula bocandei and Meliponula nebulata in Bwindi Impenetrable National Park, Uganda. African Journal of Ecology 45:265-274. |
|
|
Klein AM, Vaissiere BE, Cane JH, Steffan-Dewenter I, Cunningham SA, Kremen C, Tscharntke T (2007). Importance of pollinators in changing landscapes for world crops. Proceeding of the Royal Society B 274:303-313. |
|
|
Latham P (2015). Plants visited by bees and other useful plants of the Southern Highlands Tanzania. Croft Cottage, Forneth, Blairgowrie, Perthshire, PH10 6SW, UK 303 p. |
|
|
Lewis DH, Considine JA (1999). Pollination and fruit set in the tamarillo (Cyphomandra betacea) (Cav.) Sendt.) floral biology. New Zealand Journal of Crop Horticulture 27(2):101-112. |
|
|
Liberato MCTC, De Morais SM, Siqueira SMC, De Menezes JESA, Ramos DN, Machado LKA, Magalhaes IL (2011). Phenolic Content and Antioxidant and Antiacetylcholinesterase Properties of Honeys from Different Floral Origins. Journal of Medicinal Food 14 (6):658-663. |
|
|
Louveaux J (1984). Domestic bee and their relation with grown plants. In. Pesson P. & Louveaux J. Ed., Pollination and plant production, INRA, Paris, France pp. 527-555. |
|
|
Madikizela B, McGaw LI (2017). Pittosporum viridiflorum Sims (Pittosporaceae): A review on a useful medicinal plant native to South Africa and tropical Africa. Journal of Ethnopharmacology 205:217-230. |
|
|
Ndayikeza L, Nzigidahera B, Mpawenimana A, Théodore M (2014). Dominance d'Apis mellifera L. (Hymenoptera, Apoïdea) dans les écosystèmes naturels et les agro-écosystèmes du Burundi: risque d'érosion de la faune des abeilles sauvage. Bulletin scientifique de l'Institut national pour l'environnement et la conservation de la nature 13:72-83. |
|
|
Népidé NC, Tchuenguem FFN (2016). Pollination efficiency of Apis mellifera adansonii (Hymenoptera: Apidae) on Croton macrostachyus (Euphorbiaceae) flowers at Dang, Ngaoundéré, Cameroon. International Journal of Biosciences 3(9):75-88. |
|
|
Obeng EA (2010). Cordia africana Lam. [Internet] Record from PROTA4U. Lemmens, R.H.M.J., Louppe, D. & Oteng-Amoako, A.A. Ed. PROTA (Plant Resources of Tropical Africa/Ressources végétales de l'Afrique tropicale), Wageningen, Netherlands 24 p. |
|
|
Otiobo ENA, Tchuenguem FFN, Djiéto-Lordon C (2015). Foraging and pollination behavior of Apis mellifera adansonii (Hymenoptera: Apidae) on Physalis micrantha (Solanales: Solanaceae) flowers at Bambui (Nord West, Cameroon). Journal of Entomology and Zoology Studies 3(6):250-256. |
|
|
Pereira RLC, Ibrahim T, Lucchetti L, Da Silva AJR, De Moraes VLG (1999). Immunosuppressive and anti-inflammatory effects of methanolic extract and the polyacetylene isolated from Bidens pilosa L. Immunopharmacology 43 (1):31-37. |
|
|
Philippe JM (1991). La pollinisation par les abeilles: pose des colonies dans les cultures en floraison en vue d'accroître les rendements des productions végétales. EDISUD, La Calade, Aix-en-Provence, France 179 p. |
|
|
Porrini C, Sabatini AG, Girotti S, Ghini S, Medrzycki P, Grillenzoni F, Bortolot-ti L, Gattavecchia E, Celli G (2003). Honey Bees and Bee Products as Monitors of the Environmental Contamination. Apiacta 38:63-70. |
|
|
Potts S, Biesmeijer K, Bommarco R, Breeze T, Carvalheiro L, Franzén M (2015). Status and trends of European pollinators, Key findings of the STEP project. Pensoft Publishers, Sofia, Bulgaria 72 p. |
|
|
Roubik DW, Buchmann SL (1984). Nectar selection by Melipona and Apis mellifera (Hymenoptera: Apidae) and the ecology of nectar intake by bee colonies in a tropical forest. Oecologia 61(1):1-10. |
|
|
Segeren P, Mulder V, Beetsma J, Sommeijer R (1996). Apiculture sous les tropiques. Agrodok 32, 5ème Ed., Agromisa, Wageningen, Netherlands 88 p. |
|
|
Shen T, Li GH, Wang XN, Lou HX (2012). The genus Commiphora: a review of its traditional uses, phytochemistry and pharmacology. Journal of Ethnopharmacology 142(2):319-330. |
|
|
Sundararajan P, Dey A, Smith A, Doss AG, Rajappan M, Natarajan S (2006). Studies of anticancer and antipyretic activity of Bidens pilosa whole plant. African Health Sciences 6(1):27-30. |
|
|
Tchuenguem FFN, Messi J, Brückner D, Bouba B, Mbofung G, Hentchoya HJ (2004). Foraging and pollination behavior of the African honey bee (Apis mellifera adansonii) on Callistemon rigidus flowers at Ngaoundéré (Cameroon). Journal of the Cameroon Academy of Sciences 4:133-140. |
|
|
Tchuenguem FFN, Djonwangwé D, Messi J, Brückner D (2007). Exploitation des fleurs de Entada africana, Eucalyptus camaldulensis, Psidium guajava et Trichillia emetica par Apis mellifera adansonii à Dang (Ngaoundéré, Cameroun). Cameroon Journal of Experimental Biology 3(2):50-60. |
|
|
Tchuenguem FFN, Fameni TS, Pharaon MA, Messi J, Brückner D (2010). Foraging behaviour of Apis mellifera adansonii (Hymenoptera: Apidae) on Combretum nigricans, Eryhrina sigmoidea, Lannea kerstingii and Vernonia amygdalina flowers at Dang (Ngaoundéré, Cameroon). International Journal of Tropical Insect Sciences 30(1):40-47. |
|
|
Tesfa A, Ejigu K, Kebede A (2013). Assessment of current beekeeping management practice and honey bee flora of Western Amhara, Ethiopia. International Journal of Agriculture and Biosciences 2(5):196-201. |
|
|
Tewelde G (2006). Study on identification and establishment of floral calendar of honey bee plants in Atakilty Kebele, gray, Ethiopia. PhD thesis, Addis Ababa University, Ethiopia 72 p. |
|
|
Tobinaga S, Sharma MK, Aalbersbergetal WGL (2009). Isolation and identification of a potent antimalarial and antibacterial polyacetylene from Bidens pilosa. Planta Medica 75(6):624-628. |
|
|
Van't LL, Boot WJ, Mutsaers M, Segeren P, Velthuis H (2005). L'apiculture dans les zones tropicales. Agrodok 32, 6ème Ed., Agromisa, Wageningen, Netherlands 95 p. |
|
|
Villières B (1987). L'apiculture en Afrique Tropicale. Dossier «Le point sur» n°11, GRET, Paris, France 220 p. |
|
|
Wékéré C, Kingha TBM, Dongock ND, Djakbe JD, Faïbawa E, Tchuenguem FFN (2018). Exploitation of Jatropha curcas, Senegalia polyacantha and Terminalia schimperiana flowers by Apis mellifera (Hymenoptera: Apidae) at Dang (Ngaoundéré, Cameroun). Journal of Entomology and Zoological Studies 6(2):2072-2078. |
|
|
Williams IH, Carreck NL (1994). Land use changes and honey bee forage plants. In. Matheson A (ed.), Forage for bees in an agricultural landscape. IBRA, Cardiff, UK pp. 7-20. |
|
|
Wright GA, Skinner BD, Smith BH (2002). Ability of honeybee, Apis mellifera, to detect and discriminate odors of varieties of canola (Brassica rapa and Brassica napus) and snapgragon flowers (Antirrhinum majus). Journal of Chemical Ecology 28:721-740. |
|
|
Yédomonhan H, Tossou MG, Akoègninou A, Demènou BB, Traoré D (2009). Diversité des plantes mellifères de la zone soudano-guinéenne: cas de l'arrondissement de Manigri (Centre-Ouest du Bénin). International Journal of Biological and Chemical Sciences 3(2):55-366. |
|
|
Zumba JX, Myers GO, Clawson EL, Miller DK, Danka RG, Blanche SB (2013). Developing hybrid cotton (Gossypium spp.) using honeybees as pollinators and the roundup ready phenotype as the selection trait. Journal of Cotton Sciences 17:293-301. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


