Full Length Research Paper

ABSTRACT
The papaya tree is not very exploitable on the international market because of the post-harvest decline of the fruits. In this study, we identified arbuscular mycorrhizal fungi (AMF) from the rhizosphere of papaya in two agroecological zones of Cameroon (Njombe-Pendja: NP and Yaounde: Y). Morphological studies on the density, the rate of root colonization, the diversity of AMF species were evaluated. Molecular identification of AMF was performed by sequencing the small subunit (SSU) 18S rDNA gene region from soil samples. These genes were amplified from the AML and NS2 primers. The total number of spores per 100 g of dry soil was significantly different (P<0.05) depending on the sites, ranging from 391 in NP to 790 in Y sites. The frequency and intensity of root colonization was not significantly different in the two sites. Eleven (11) AMF belonging to seven (7) genera (Scutellospora, Gigaspora, Acaulospora, Entrophospora, Funneliformis, Glomus and Racocetra) were recorded. Two AMF species (T1: Scutellospora rosa and T6: Racocetra gregaria) were recorded at NP and absent at Y. The Shannon-Wiener index revealed a low diversity of AMF species. Homogeneity of species was recorded. Molecular analysis reveals that the AMF species obtained have sizes of 250 bp.
Key words: Arbuscular mycorrhizal fungi, Carica papaya L., fungal, molecular and morphological identification, rhizosphere.
INTRODUCTION
Cameroon, commonly known as ‘Africa in miniature’ has favorable ecological conditions for the cultivation of tropical fruit species. The papaya tree (Carica papaya L.) is one of the most cultivated fruit trees of the Caricaceae family (Lim, 2012) in this country. It is a very important species from an economic point of view. It represents about 16% of the total production of tropical fruits in the world (FAOSTAT, 2019). C. papaya L. has very high nutritional and nutraceutical properties (Santana et al., 2019). It is an important source of vitamin A, ascorbic acid, minerals (iron, calcium, potassium, etc.), polysaccharides and proteins (Daagema et al., 2020). Its fruit is highly appreciated by consumers for its taste, visual quality, organoleptic properties, nutritional, therapeutic, digestive and dietary values ??(Bruneton, 2009). It has a lifespan of 5 years with an economic life of 2 years (N'da et al., 2008). Its world production is estimated at 13,016,280 tonnes/year (EUROSTAT, 2020).
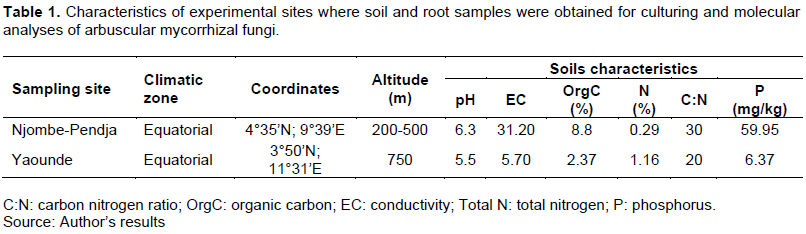
In Cameroon, C. papaya is grown mainly in humid forest with monomodal rainfall, more precisely in the Littoral zone (Njombe-Pendja) where the ecological conditions are ideal (equatorial-type climate; altitude: 20 to 500 m; average temperature 30°C, rainfall 2350 mm, volcanic type soil). National papaya production is estimated at 700 tonnes over an area of ??25 ha. Its still very low export (36 tonnes) (EUROSTAT, 2020) mainly concerns the dried forms of the product, despite the inclusion of this fruit in government strategies for the diversification of agricultural exports from Cameroon. The demand for this fruit on the international market is estimated at 17,519 tonnes/year against local production of around 38 tonnes/year (EUROSTAT, 2020). This low production, despite the very high demand, is linked to numerous biotic and abiotic constraints such as the rapid maturation of the fruit (Zhu et al., 2020), the lack of control of the favorable stage for harvesting the fruit (Greenwald et al., 1998), post-harvest handling (Elik et al., 2019), sensitivity to stress (water), parasitic and fungal diseases (Zhu et al., 2020), and uptake of soil mineral elements such as calcium (Hocking et al., 2016). In Cameroon, depending on the variety, some highly productive papayas unfortunately face increased post-harvest loss, affecting production sometimes going to more than 50% yield loss (FAOSTAT, 2019). However, with the ever-increasing population (7.7 billion in 2020 to 9.7 billion by 2050) (United Nations, 2020), it would be crucial to strengthen the papaya sector by intensifying production and improving the post-harvest quality of papaya fruits. The success of this challenge involves the soil microorganisms naturally associated with papaya. Furthermore, its cultivation requires continuous application of fertilizers for its development and continuous production (Nakasone and Paull, 1998), but the use of inorganic fertilizers in agricultural agroecosystems has generated environmental problems such as eutrophication of water bodies. This has sparked interest in the use of beneficial microorganisms such as arbuscular mycorrhizal fungi (AMF) to improve the productivity of this crop (Cuenca et al., 2007). Throughout the world, most papaya production comes from poorly structured units (home gardens, extensive fields, etc.) for which the farmer places little importance on the diversity of microorganisms found in the rhizosphere. However, knowledge of the activity of these living organisms in the soil (microbes, roots of living plants, etc.), not only constitutes an important portion of total biodiversity, but also a plus on the yield and product quality for international trade (Decaëns et al., 2006). However, knowledge of these microorganisms constitutes part of the solution in reducing post-harvest losses and a decisive step in the perspective of developing papaya cultivation (Marcos et al., 2020). Among the most important soil microorganisms are AMF which form beneficial mutualistic associations between species of the Glomeromycotina (Spatafora et al., 2016; Davison et al., 2018) and the roots of 80% of vascular plants (Smith and Read, 2008). They are considered essential for plant performance and nutrition (Sadhana, 2014; Powell and Rillig 2018) and are vital components of the soil in natural and agricultural systems. They are one of the most common and widespread groups of soil organisms and are symbionts of a majority of plant species. Therefore, AMF diversity and interaction with plants in certain ecosystems are not fully understood (Davison et al., 2015). Nevertheless, some studies report the colonization of papaya by AMF (Sukhada, 1989; Trindade et al., 2006; Khade and Rodrigues, 2008a, 2009a; Sankaralingam et al., 2016). Among these studies, Trindade et al. (2006), Khade et al. (2009b), Rodríguez (2011), and Marcos et al. (2011) identified the species belonging to the genera Gigaspora, Glomus, Acaulospora, Racocetra, Dentiscutata and Funniliformis colonizing papaya, based on the morphological characters of AMF spores.
Identifying AMF species that can be used in papaya cultivation is a future challenge. AMF can be useful in the development of effective methods to reduce post-harvest losses (Kamda et al., 2021). This concept is far from being applied in practice due to the lack of understanding of the functioning of AMF species (Scullion et al., 1998). In addition, the identification of AMF species in the papaya rhizosphere based on molecular methods is not much applied and yet very reliable since it can provide information on the exact type of AMF colonizing papaya. In Cameroon, few studies on this molecular approach concerning the communities of AMF species of papaya have been made (Azcón-Aguilar et al., 2003; Bouazza et al., 2015). Most of the studies were based on the morphological approach. The prospects for the exploitation of these AMF in the reduction of post-harvest losses of C. papaya L. fruits and the production of organic plants in Cameroon are still lacking. The aim of this study is to determine the diversity of AMF species associated with papaya (C. papaya L.) in two different agro-ecological zones of Cameroon.
MATERIALS AND METHODS
Study sites
Two sites were selected to collect soil from the rhizosphere of C. papaya L. They were geographically separated and one represents the most exploited site (Table 1 and Figure 1). The two sites were located in two different agro-ecological zones: (1) the forest zone with the monomodal rainfall, and (2) the bimodal rain forest zone (Figure 1).
Soil and root sampling
Sampling was carried out in two different sites, one located in the humid forest zone with monomodal rainfall and the other in the humid forest zone with bimodal rainfall in Cameroon during the rainy season in March 2019. Plots sized 01 ha with approximately 2,000 papaya plants.ha-1 (one year old) were marked in each site. The soil sampling method is that of Sieverding (1983). Indeed, for each site, an area of ??one hectare was subdivided into 100 quadrants of dimension 10×10 m each. In each of these quadrants, 100 papaya plants were randomly counted. About twenty papaya trees were chosen. In the rhizosphere of each papaya tree, the sampling of soil and roots was carried out between 0 and 25 cm depth in the ground using a den. This method was also applied in the soil outside the rhizosphere (non-rhizosphere) of papaya (with corn as previous crop). The fine roots of the plants were collected at the same time at the rate of 2 g per plant. These roots were then mixed and the whole was preserved in 50% alcohol.
Trap culture
The trap culture was carried out in a greenhouse at the Regional Laboratory of Biological Control and Applied Microbiology of IRAD Nkolbison (Yaounde, Cameroon). Sorghum and cowpea (mycotrophic plants) were used as trap plants. They were grown on a previously disinfected low-nutrient substrate consisting of a mixture of Sanaga sand and black soil in the proportions 3:1 (V/V). Five-liter pots were filled. The mycorrhizal inoculum consisting of soil and roots from each soil sample was placed on the surface of the substrate in the pots. Sorghum and cowpea seeds were disinfected with an 8° bleach solution diluted to 10%, then rinsed thoroughly with sterile distilled water and pre-germinated in the dark. In each pot, four seeds were placed on the inoculum and the whole covered with substrate in order to limit the risk of dehydration. Five replicates were set up per plant soil origin. Trap plant species were chosen because they are well known as AMF host plants and are frequently used as trap crops (Ngonkeu, 2009).
The pots were watered twice a week with deionized water and fertilized with 100 mL of Rorison's low phosphorus nutrient solution 120.02 g/L MgSO4.7H2O, 238.04 g/L Ca (NO3)2.4H2O, 115.38 g/L KH2PO4, 3H2O, 12.500 g/L [Fe EDTA, 1.121 g MnSO4.4H2O, 1.421 g H3BO3, 0.093 g (NH4)6Mo24.4H2O, 0.220 g ZnSO4.7H2O, 0.198 g CuSO4.5H2O]/L (Ngonkeu, 2009). Two weeks after sowing, the plants were thinned and two plants per species were retained. After 6 (six) months (two cycles) of cultivation, watering was stopped and the pots were subjected to water stress for two weeks at room temperature in order to induce the massive production of spores (sporulation). At the end of two growth cycles (three months/cycle), plants were harvested and host plant roots collected for analysis of mycorrhizal colonization rates.
Percentage of colonization of AMF of C. papaya L
Percent colonization of C. papaya L. AMF root length was calculated by the grid intersection method (Ngonkeu, 2009) after elimination in 10% KOH for 15 min at 90°C and staining with blue Trypan (0.05%) for 30 min (Phillips and Hayman, 1970; Brundrett et al., 1996). Frequency of mycorrhization ‘F’ was calculated by F% = 100 (N-no) / N, where ‘N’ is the number of fragments observed and ‘no’ the number of these fragments with no trace of mycorrhization. ‘F’ reflects the importance of the contamination. Intensity of mycorrhization ‘I’ was calculated by I% = (95n5 + 70n4 + 30n3 + 5n2 + n1) / N, where n5, n4 …n1 denote, respectively the numbers of fragments noted 5, 4, ….1 (Trouvelot et al., 1986). AMF spores were isolated from the trapping soil by wet sieving and settling (Ngonkeu, 2009). The spores were identified according to taxonomic criteria (Schenck and Perez, 1990) and also using information from INVAM (http://www.invam.caf.wdu.edu/).
Species diversity of AMF
The diversity of AMF (Castillo et al., 2016) was evaluated through the species richness (S), the Shannon-Wiener diversity index (H') (Shannon-Wiener, 1948); the Pielou fairness index (J’) (Pielou, 1966), the Simpson dominance index (l), and the Simpson diversity index (Ds) (Simpson, 1949). Specific richness refers to the total number of species in a soil sample. The Shannon-Wiener diversity index takes into account both the particular species richness and the abundance of each species. Its expression is H’. Pi is the proportional abundance of the species (Pi = ni / N); ni is the number of individuals of a species in the sample; N is the total number of individuals of all species in the sample.
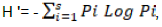
where Pi is the proportion of species "i" to the total population, and S is the total number of species. The Shannon-Wiener index is often accompanied by the Pielou fairness index, which represents the ratio of H' to the theoretical maximum index in the stand H' max. J’ = H’/log(S), with H'max = log2S. This index can vary from 0 to 1; it is maximal when the species have identical abundances in the stand, and it is minimal when a single species dominates the whole stand.
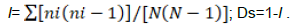
Molecular analysis
Extraction of spore DNA from soil
DNA extraction was performed at the Common Laboratry of Microbiology (LCM) IRD/ISRA/UCAD, Research Center of Bel-Air Dakar/Senegal. 0.5 g of soil from each treatment at both study sites were measure. The kits "FastDNA® SPIN Kit for Soil" designed for use with FastPrep® instruments from MP Biomedicals and "DNeasy Plant Mini kit " were used respectively. DNA extractions were performed according to the manufacturers' instructions. DNA extracts were quantified using a NanoDrope to assess the DNA concentration of the sample for immediate reading of the result. It measures the concentration and purity of DNA extracted with 1 µL of sample indicating an absorbance ratio at 260 and 280 nm. This 260/280 ratio must be close to 1.8 to qualify the samples as pure. Values ??below 1.8 would indicate the presence of impurities or absorbing proteins around the same wavelengths.
PCR amplification of 18S SSU rDNA region
PCR amplification in both DNA extraction processes (from soil), small subunit (SSU) regions of rDNAs was respectively used as target regions for PCR experiments. The reactions were carried out in a final volume of 25 μL containing 5 μL of buffer (5X Green Go-TaqReaction Buffer), 2 μL of dNTPs (10 mM of each dNTP), 0.125 μL of Go-Taq DNA polymerase (2.5 Units /reaction), 1 μL of each primer (front and reverse) (20 pmol/μl) and 14 μL of sterile water. The first amplification of fungal DNA was performed using primer pair NS31 (5' TTG GAG GGC AAG TCT GGT GCC-3') and AML2 (5'GAA CCC AAA CAC TTT GGT TTC C-3') (White et al., 1990). The PCR program was as follows, initial denaturation (94°C, 5 min), followed by 30 cycles of denaturation (94°C, 30 s), primer annealing (58°C, 1 min); elongation (72°C, 1min 20 s) and a final elongation (72°C, 10 min). A negative control without DNA was included in the experiment to serve as a PCR control. The molecular weight marker used is 100 bp. In order to verify the efficiency of the PCR, a negative control using sterile water was used in all the amplifications. The PCR products were visualized by electrophoresis on a 1% (w/v) agarose gel in 1X TAE (Tris Acetate EDTA) buffer, then photographed under UV light.
Data analysis
Data on spore density, mycorrhization rate and AMF diversity were subjected to analysis of variance (ANOVA). Means were compared using Tukey's and Ducan's tests (p<0.05) in R studio software, version 4.0.5.
RESULTS
Number of AMF spores per site
AMF spores were present in all soil samples. Figure 2 shows higher spore densities in non-rhizospheric soils than in rhizospheric soils at both sites. These densities are, respectively around 1009 and 435 spores per 100 g of dry soil in Njombe-Pendja and 836 and 813 spores per 100 g of dry soil in Yaounde. The comparison of the different sites shows that, for the two types of papaya farms, the densities of AM fungus spores also varied according to the sites. Indeed, the highest densities were noted in Yaounde (813 spores/100 g of dry soil) and the lowest in Njombe-Pendja (435 spores per 100 g of dry soil). The total number of spores was not significantly different at the threshold (p<0.05) in the two study sites. However, a significant difference at the threshold (p<0.05) between the number of spores in the soils of the two sites and the negative control (soil/sand substrate) was observed.
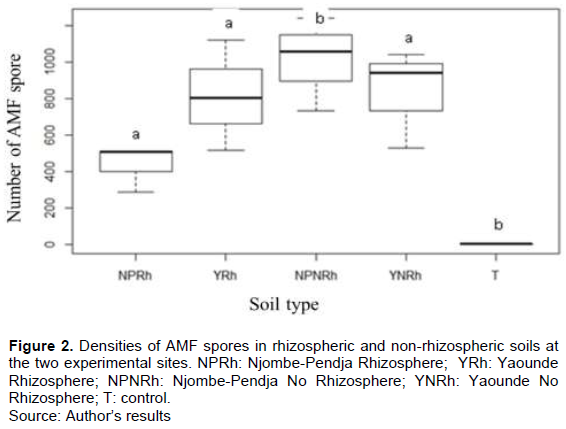
Number of AMF spores according to sites
The comparison of the different sites shows that, for the different types of papaya soil, the densities of AMF spores vary according to the sites (Figure 3). The highest densities were noted in Yaounde (>790 spores/100 g of dry soil) and the lowest in Njombe-Pendja (>391 spores per 100 g of dry soil) for small spores. The difference was not significant at the threshold (p<0.05) for medium and large spores in the two study sites. An abundance of small spores was observed in the different soil types of the two study sites.
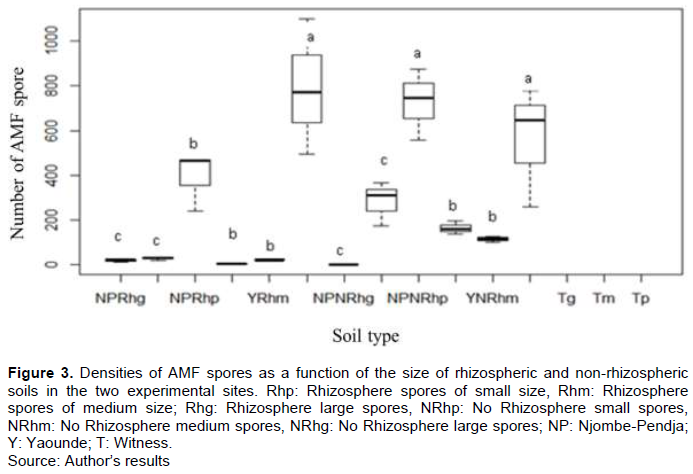
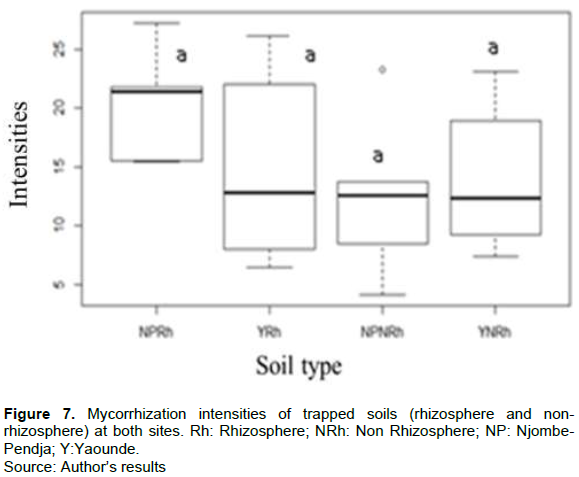
Mycorrhizal colonization of the roots of C. Papaya L.
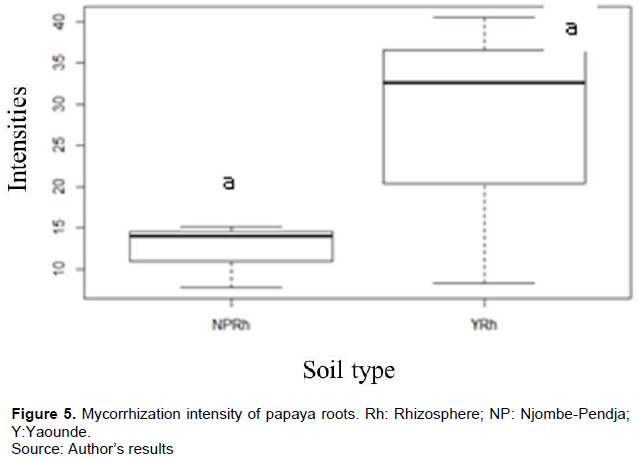
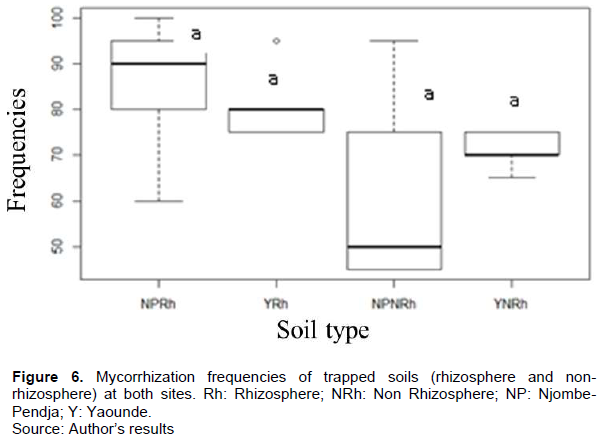
The natural presence of AMF in the roots of C. papaya L. was not significantly different in the different agro-ecological zones (monomodal rainfall zone represented by Njombe-Pendja and bimodal rainfall zone represented by Yaounde). The frequency of colonization was between 83.33 and 93.33%. The very low colonization intensity was between 12.3 and 27.13% (Figures 4 and 5). However, after six months of trap culture with Sorghum (Sorghum bicolor) and cowpea as host plants, the intensity of mycorrhization in the two sites varied between 15 and 21% for rhizospheric soils and 12 and 15% for non-rhizospheric soils. The natural presence of AMF in the roots of C. papaya L. was not significantly different in the different agro-ecological zones. The frequency of colonization was between 83.33 and 93.33%. The very low colonization intensity was between 12.3 and 27.13% (Figures 6 and 7).

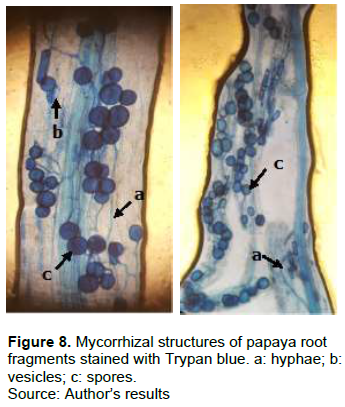
The roots of C. papaya L. showed mycorrhizal structures in the form of vesicles, hyphae and spores (Figure 8).
Morphological characteristics of AMF and community composition
The morphological characters of the isolated spores were studied and the data obtained were compared with the original description of Schenck and Pérez (1990) and with those of the database of reference cultures published on the website http://invam. caf.wvu.edu to identify them. Eleven (11) morphotypes of AMF were identified from the papaya rhizosphere based on morphological characters (Table 2). The identified morphotypes belong to 07 (seven) genera (Scutellospora, Gigaspora, Acaulospora, Entrophospora, Funneliformis, Glomus and Racocetra) classified into four families (Gigasporaceae, Glomeraceae, Scutellosporaceae and Acaulosporaceae). Among the morphotypes belonging to the genus Scutellospora, two have been identified at the species level (Scutellospora rosa and Scutellospora verrucosa). The genus Acaulospora was represented by two species (Acaulospora delicatea and Acaulospora tuberculata). The genus Glomus was represented by two species (Glomus manihotis and Glomus etunicatum). The genera Funneliformis, Racoceta and Entrophospora were each represented by a single species (Funneliformis mosseae, Racoceta gregaria and Entrophospora colombiana, respectively). The genus Gigaspora was represented by one specy (Gigaspora margarita).

The pictures of the spores of some species of AMF (arbuscular mycorrhizal fungi) isolated in the rhizosphere of papaya are presented in Figure 9.

All the AMF spores identified in the two sites belong to the Glomeromycota phylum according to the new classification of Schüβler and Walker (2010) and Redecker et al. (2013).
Specific richness
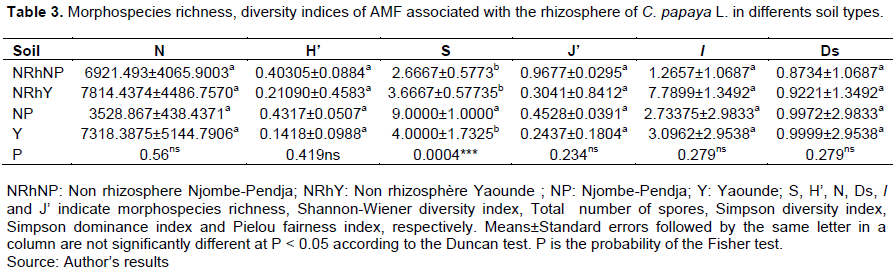
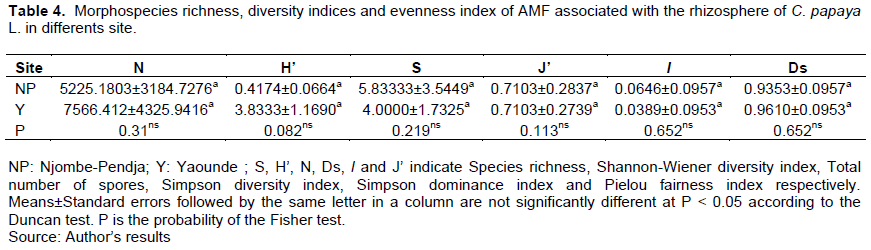
AMF species richness, Shannon-Wiener diversity index, Simpson diversity index, Simpson dominance index, and Pielou fairness index in the rhizosphere of C. papaya L. were assessed in each site (Tables 3 and 4). All these parameters were not significantly different according to Duncan’s test (P < 0.05). The species richness was more signi?cantly different (P < 0.001). Species richness was the same in all different soil types. The species richness of the species between the type of soils was the same. This reflects the homogeneity of species. On the other hand, the species richness of the species between the sites was different, which reflects the heterogeneity of the species between the sites. In addition, low species richness was observed in the soils of NP and Y. This is justified by the Shannon-Wiener index whose values ??tend towards zero (0) with, respectively (H'= 0.43 and 0.14) in the soil types of NP and Y and a high species richness observed at the site of Y. Despite this difference, the analyses revealed that there was no significant difference between the soil types and the study sites. Pielou's fairness index (J'), which measures the proportion of individual organisms within each species, is inequitable between species from different soils and study sites. Indeed, there was no significant difference between the indices of soil types and study sites. The indices tend towards zero with respectively 0.45 and 0.24 in the soils of NP and Y. The values ??of Ds tend towards 1, which shows that the sites are biologically diversified.
Relative abundance
The relative abundance of AMF species in the rhizosphere of C. papaya L. in two agro-ecological zones of Cameroon is as shown in Figure 10. In humid forest with monomodal rainfall (NP), eight morphotypes of AMF (T1: S. rosa; T2: G. margarita, T4: F. mosseae, T5: Glomus spp., T6: R. gregaria, T8: S. verrucosa and T11: G. etinucatum) were isolated and identified. In humid forest with bimodal rainfall (Y) on the other hand, four (4) morphotypes specific to papaya trees (T2: G. margarita; T5: Glomus spp., T9: E. colombiana and T11: G. etinucatum) were isolated and identified. Four AMF morphotypes (T2: G. margarita, T5 Glomus spp., T9: E. colombiana, and T11: G. etinucatum) common to the two study sites were identified. Two morphotypes of AMF (T1: S. rosa and T6: R. gregaria) on the other hand were identified only in NP. This low species richness at NP and Y was confirmed by the Shannon-Wiener index where H' was 0.43 and 0.14, respectively. All the morphotypes represented in the rhizosphere of C. papaya L. in the Yaounde site were found in the NP site and those represented in the NP site were not all present in Yaounde. The T7 morphotype (G. manihotis) was represented in all soil types.

Molecular characterisation of AMF
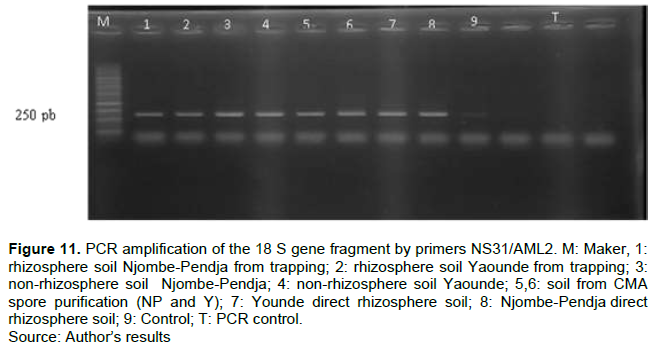
Figure 11 shows the results of the amplification of the 18 S small subunit (SSU) rDNA gene from papaya rhizosphere soil samples using AMF primers NS31 and AML2. This amplification allowed us to clearly distinguish on the electrophoregram (Figure 11) a variable number depending on the soil type. These soil samples contain AMF belonging to Glomeromycotina. The size of the bands is 250 bp showing the presence of fungal communities in the soils of the different sites studied.
DISCUSSION
Spore density of AMF
The rhizospheric soils of the different study sites showed a high density of AMF spores (836 spores/100 g of dry soil) in humid forest with bimodal rainfall in the locality of Yaounde where the soils are acidic (pH = 5.5) and in humid forest with monomodal rainfall in the locality of Njombe-Pendja (435 spores/100 g of dry soil) where the soils are less acidic (pH= 6.1). The soils of this locality which are basic in nature have become less acidic due to the excessive use of chemical fertilizers.
In addition, certain cultural practices such as monoculture lead to a decrease in spores density compared to crop rotation (Oehl et al., 2003; Mathimaran et al., 2007). Monoculture which is regularly practiced in the locality of Njombe-Pendja, justifies our results. Moreover, the low density of AMF spores could be explained by the excessive use of chemical fertilizers (NPK) which are applied in the locality of Njombe-Pendja. Sas-paszt et al. (2020) affirmed this in their work on the diversity of AMF in the rhizosphere of Solaris and Regent Vine plants treated with bioproducts. An abundance of small spores was observed in humid forest with monobinal rainfall than in humid forest with bimodal rainfall. This result is consistent with those of Johnson et al. (2013) and Bossou et al. (2019) who noted that the number of AMF spores is inversely proportional to their size.
Roots colonization and community composition of AMF
The colonization frequencies of papaya roots and soils (rhizospheric and non-rhizospheric papaya) were not significantly different in the two study sites. They were high in both sites and varied between 83.33 and 93.33% for papaya roots and between 81 and 85% for rhizospheric soils and 62 and 73% for non-rhizospheric soils. These results are similar to the findings of Khade et al. (2002) who reported high root colonization (78%) in papaya roots. Much colonization of the roots could be justified by the fact that the papaya crop has a root system characterized by the presence of a taproot and adventitious roots which allow them to draw their nutrients only from the superficial horizons. Thus, AMF allow root development so that they can improve the uptake of water and nutrients from the soil. This result is contrary to those of Chebet et al. (2021) and Jaizme et al. (2006) who obtained a low rate of mycorrhizal infection (43 and 15% to 31%, respectively) in papaya, who also obtained a relatively low colonization rate of papaya roots. This could be due to the fact that the mycorrhizal infection of plants varies significantly from one plant to another, and within the same species.
Species richness and community composition of AMF
Morpho-anatomical characterization techniques are mainly based on morphological traits such as spore size, color and mode of attachment of hyphae, and anatomical features such as the number and diameter of layers making up the spore membrane. Nowadays, this type of characterization remains widely used in studies on diversity (Wang et al., 2019; Vieira et al., 2020). The use of these techniques allowed us to identify 11 morphotypes of AMF in the rhizosphere of papaya grown in soils collected from two sites in Cameroon. These AMF morphotypes belong to 7 genera (Gigaspora, Racocetra, Scutellospora, Entrophospora, Glomus, Acaulospora, and Entrophospora) classified into four families (Gigasporaceae, Acaulosporaceae, Scutellosporeae and Glomeraceae). This result is similar with the number of morphotypes but different at the gender level from the work of Trindade et al. (2006) who had identified 11 morphotypes of the papaya rhizosphere in Brazil belonging to the genera Acaulospora, Dentiscutata, Gigaspora, Glomus and Racocetra. It is contrary to the results of Sankaralingam et al. (2016) who had identified species of the genus Glomus and Acaulospora in the Rhizosphere of papaya. This specific richness was lower than that obtained for others such as squash cultivation in Benin and Cameroon by Mbogne et al. (2015), Bambara pea cultivation in Cameroon by Temegne et al. (2017), maize cultivation in Benin by Bossou et al. (2019), and on Vigna umguiculata in Senegal by Diop et al. (2021). Species richness would therefore vary from one crop to another. The four genera of AMF spores found during this study are identical to those identified by Mbogne et al. (2015) and Johnson et al. (2013), Bossou et al. (2019), and Diop et al. (2021). These four genera are generally those identified, in the rhizosphere of other cultures, by different microbiologists from the West African and central regions (Ngonkeu et al., 2013; Temegne et al., 2017). The genus Glomus was represented in all the soils collected in the two study sites. This distribution of the genus Glomus is comparable to previous observations made in Cameroon (Ngonkeu et al., 2013, in Brazil (Samba-Mbaye et al., 2020; Teixeira et al., 2017; Vieira et al., 2020), Morocco (Sellal et al., 2016) and China (Song et al., 2019). The predominance of species of the genus Glomus in all sites suggests a better adaptation of this genus either to the most hostile conditions such as drought, salinity and other environmental stresses (Blaszkowski et al., 2018), or to a wide range of ecological niches (Houngnandan et al., 2009). Indeed, the Glomus genera would propagate much more by spores which are forms of resistance of AMF to difficult conditions while the Gigaspora and Scutellospora genera would propagate more with other types of propagules such as hyphae and extra mycelial fragments roots (Brito et al., 2012). It would also be due to the fact that its development cycle is not affected by repeated cultivation on land compared to those of minority genera such as Acaulospora, Gigaspora and Scutellospora (Oehl et al., 2003). The low species diversity would also be due to the fact that, in our study, the determination of the diversity index was made on the basis of the morphological characterization of the spores obtained after trapping culture. Indeed, the trap culture only partially reveals the diversity of AMF in the soil (Bever et al., 2001; Wubet et al., 2004). Moreover, some AMF species could not be revealed due to an established selective association between the host plant and the fungus (Kardol et al., 2006).
Relative abundance
In humid forest with monomodal rainfall (NP), seven morphotypes of AMF (T1: S. rosa; T2: G. margarita, T4: F. mosseae, T5: Glomus spp., T6: R. gregaria, T8: S. verrucosa, and T11: G. etinucatum) were isolated and identified. In humid forest with bimodal rainfall (Y) on the other hand, four (4) morphotypes specific to papaya trees (T2: G. margarita; T5: Glomus spp. T9: E. colombiana and T11: G. etinucatum) were isolated and identified. Two species of AMF (T1 and T6) were recorded at NP and absent at Y. This low difference in species richness in the two sites could be explained by the physico-chemical properties of the soil at these sites. Indeed, the irregular spatial distribution of the spores of arbuscular mycorrhizal fungi and the complex structure of the constituents of the rhizosphere affect the density of the spores (Zhao et al., 2001). In the two types of soils (Y and NP), the mean values ??of the Shannon-Wiener and Pielou indices varied from 0.14 to 0.43, respectively. They were low in both soil types. This result is similar to those of Mbogne et al. (2015) who recorded a small difference in species richness of squash in Cameroon and Benin. From our studies, the genera Glomus and Gigaspora could be well designated for the production of inocula under the conditions of papaya cultivation in the study sites (Yaounde and NP).
Molecular characterization of AMF
The amplification of the 18 S gene of the rDNA of the small subunit of the soil samples of the papaya rhizosphere was carried out using the pair of primers NS31 (5 'TTG GAG GGC AAG TCT GGT GCC- 3') and AML2 (5'GAA CCC AAA CAC TTT GGT TTC C-3'). The purpose of PCR is to separate the different fragments of nucleic acids in relation to their size. The primers used make it possible to amplify gene portions of size 1100 pb (Simon et al., 1992). Our results show that the soil samples contain AMF sized 250 pb.
CONCLUSION
Investigations on the mycorrhizal status of AMF in the rhizosphere of papaya in two agro-ecological zones of Cameroon (humid forest zone with monomodal rainfall: NP and the humid forest zone with bimodal rainfall: Y) revealed the presence of AMF in the different soils of the two study sites. Papaya harbors arbuscular mycorrhizae with characteristic structures such as hyphae, vesicles and spores. The spores were morphologically different depending on the size, color, shape, presence or absence of the hypha and its mode of attachment to the spore. From these morphological criteria, eleven species of AMF (A. rosa, G. margarita, Acaulospora delicata, Funneliformis mosseae, Glomus sp., R. gregaria, G. manihotis, S. verrucosa, E. colombiana, A. tuberculata, G. etunicatum) belonging to the families Gigasporaceae, Acaulosporaceae, Glomeraceae and Scutellosporaceae were identified. A dominance of small-sized spores (> 45 µm) of the genus Glomus was observed in the different soils of the two study sites. In Njombe-Pendja, which is the papaya cultivation area par excellence, two species S. rosa and R. gregaria are present but absent in Yaounde. The frequency of colonization was high (83.33 and 93.33%) and the intensity of mycorrhization low (12.3 and 27.13%) in the two study sites. Species richness was identical between soil types and differed between study sites. The species identified were inequitably distributed in the different soils and study sites, and biologically diverse between the study sites. The amplification of the 18 S gene of the rDNA of the AMF of the different study soils indicates that these soils harbor AMF with sizes of 250 bp. The choice of sites is a factor to be taken into account to promote the establishment of mycorrhizal symbiosis for profitable papaya cultivation. These AMF would contribute to the regulation and absorption of calcium during the fruit ripening process. Indeed, calcium would reinforce the rigidity of the fruit pulp which would contribute to the reduction of post-harvest losses and increase storage time for exploitation on the international market.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors are grateful for the moral and institutional support from the University of Yaounde I, Regional Laboratory of Biological control and Applied Microbiology of IRAD and Common Microbiology Laboratory (LCM) IRD/ISRA/UCAD, Research Center of Bel-Air, Dakar/Senegal. This project has received financial support from Agence Universitaire de la Francophonie (AUF).
REFERENCES
|
Ahmad S, Thompson AK, Perviez MA, Anwar N, Ahmad F (2006). Effect of fruit size and temperature on the shelf life and quality of ripe banana fruit. Journal of Agriculture Research 44(4):313-324. |
|
|
Azcon-Aguilar C, Palenzuela J, Roldan A, Bautista S, Vallejo R, Barea JM (2003). Analysis of the mycorrhizal potential in the rhizosphere of representative plants species from desertification -threatened Mediterranean shrublands. Applied Soil Ecology 22(1):29-37. |
|
|
Azcón-Aguilar CA, Barea JM (1997). Applying mycorrhiza biotechnology to horticulture : significance and potentials. Scientia Horticulturae, 68(1-4):1-24. |
|
|
Bever JD, Schultz PA, Pingle A, Morton JB (2001). Arbuscular mycorrhizal fungi: more diverse than meets the age, and the ecological tale of why. BioScience 51(11):923-931 |
|
|
B?aszkowski J, Koz?owska A, Niezgoda P, Goto BT, Dalpé Y (2018). A new genus, Oehlia with Oehlia diaphana comb. nov. and an emended description of Rhizoglomus vesiculiferum comb. nov. in the Glomeromycotina. Nova Hedwigia 107(3-4):501-518. |
|
|
Bossou LDR, Houngnandan HB, Adandonon A, Zoundji C, Houngnandan P (2019). Diversité des champignons mycorhiziens arbusculaires associés à la culture du maïs (Zea mays L.) au Bénin. International Journal of Biological and Chemical Sciences 13(2):597-609. |
|
|
Bouazza MK, Ighilhariz Z, de Lajudie P, Duponnois R, Bekki A (2015). Assessing the native arbuscular mycorrhizal symbioses to rehabilitate a degraded coastal sand dune in Algeria. International Journal of Agriculture and Crop Sciences 8:194-202. |
|
|
Brito I, Carvalho M, Goss MJ (2021). Managing the functional diversity of arbuscular mycorrhizal fungi for the sustainable intensification of crop production. Plants people Planet 3:491-505. |
|
|
Brundrett MC, Bougher NB, Grove T, Malajczuk N (1996). Working with mycorrhizas in forestry and agriculture. ACIAR Monographe 32:374. |
|
|
Bruneton J (2009). Pharmacognosie phytochimie, plante medicale, 4ème edition et augmenentée, Tec et Doc- Edition Médicales Internationales P 188. |
|
|
Castillo CG, Oehl F, Sieverding E (2016). Arbuscular mycorrhizal fungal diversity in wheat agro-ecosystems in Southern Chile and effects of seed treatment with natural products. Journal of Soil Science and Plant Nutrition 16(4):967-978. |
|
|
Chebet D, Kariuki W, Wamocho L, Rimberia F (2020). Effect of Arbuscular mycorrhizal inoculation on growth, biochemical characteristics and nutrient uptake of passion fruit seedlings under flooding stress. International Journal of Agronomy and Agricultural Research 16(4):24-31. |
|
|
Cuenca G, Cáceres A, Oirdobro G, Hasmy Z, Urdaneta C (2007). Las micorrizas arbusculares como una alternativa de agricultura sustentable en áreas tropicales. Interciencia 32(1):23-29. |
|
|
Daagema AA, Orafa PN, Igbua FZ (2020) . Nutritional Potentials and Uses of Pawpaw (Carica papaya). European Journal of Nutrition and Food Safety 12(3):52-66. |
|
|
Davison J, Moora M, Öpik M, Adholeya A, Ainsaar L, Bâ A, Burla S, Diedhiou AG, Hiiesalu I, Jairus T, Johnson NC, Kane A, Koorem K, Kochar M, Ndiaye C, Pärtel M, Reier Ü, Saks Ü, Singh R, Vasar M, Zobel M (2015). Global assessment of arbuscular mycorrhizal fungus diversity reveals very low endemism Science 349(6251):970-973. |
|
|
Davison J, Moora M., Öpik M, Ainsaar L, Ducousso M, Hiiesalu I (2018). Microbial island biogeography: isolation shapes the life history characteristics but not diversity of root-symbiotic fungal communities. The ISME Journal 12(9):2211-24. |
|
|
Decaëns T, Jiménez JJ, Gioia C, Lavelle P (2006). The values of soil animals for conservation biology. European Journal of Soil Biology 42:S23-S38. |
|
|
Diop I, Ndoye F, Diédhiou A, Krasova-wade T, Do rego F, Noba K, Ambrosi JP, Kane A (2021). Diversity and spore density of arbuscular mycorrhizal fungi in the rhizosphere of Cowpea (Vigna unguiculata [L.] Walp.) cultivated in different soils in Senegal. Journal of Animal and Plant Sciences 48(1):8552-8565. |
|
|
Elik A, Kocak Y, Istanbullu Y, Guzelsoy NA, Yavuz A, Gogus F (2019). Strategies to Reduce Post-Harvest Losses for Fruits and Vegetables. International Journal of Scientific and Technological Research 5(3):2422-8702. |
|
|
EUROSTAT (2020). Your key to European Statistics: Where are our fruit and veg produced?. |
|
|
FAOSTAT (2019). Food and Agriculture Organization of the United Nations. |
|
|
Greenwald AG, McGhee DE, Schwartz JLk (1998). Meauring individual differences in implicit cognition: The implicit association test. Journal of personality and social Psychology 74(6):1464-1480. |
|
|
Hocking B, Tyerman SD, Burton RA, Gilliham M (2016). Fruit calcium: Tranport and physiology. Frontiers in Plant Science 7:569. |
|
|
Houngnandan P, Yemadje RGH, Kane A, Boeckx P, Van Cleemput O (2009). Les glomales indigènes de la forêt claire à Isoberlinia dokaCraibet Stapf à WariMaro au centre du Bénin. Tropicultura 27(2):83-87. |
|
|
IRAD (2016). Zones Agroecologiques du Cameroun, Centres, Stations et Antennes. Institute of Agricultural Research for Development, Yaoundé P 1. |
|
|
Jaizme-Vega MC, Rodríguez-Romero AS, Barroso Núñez LA (2006). Effect of the combined inoculation of arbuscular mycorrhizal fungi and plant- growth promoting rhizobacteria on papaya (Carica papaya L.) infected with the root-knot nematode Meloidogyne incognita. Fruits 61(3):1-7. |
|
|
Johnson JM, Houngnandan P, Kane A, Sanon K, Neyra M (2013). Diversity patterns of indigenous arbuscular mycorrhizal fungi associated with rhizosphere of cowpea Vigna unguiculata L. Walp. in Benin, West Africa. Pedobiologia 56(3):121-128. |
|
|
Kamda AG, Ponka R, Frazzoli C, Fokou E (2021). Waste of Fresh Fruits in Yaounde, Cameroon: Challenges for Retailers and Impacts on Consumer Health. Agriculture 11:89. |
|
|
Kardol P, Bezemer TM, Van Der Putten WH (2006). Temporal variation in plant-soil feedback controls succession. Ecology Letters 9:1080-1088. |
|
|
Khade SW, Bukhari MJ, Jaiswal V, Gaonkar UC, Rodrigues BF (2002). Arbuscular mycorrhizal status of medicinal plants: A field survey of AM fungal association in shrubs and trees. Journal of Economic and Taxonomic Botany 26(3):571-578 |
|
|
Khade SW, Rodrigues BF (2008a). Spatial variations in arbuscular mycorrhizal (AM) fungi associated with Carica papaya L. in a tropical agro-based ecosystem. Biological Agriculture and Horticulture 26:149-174. |
|
|
Khade SW, Rodrigues BF (2009a). Spatio-temporal variations of arbuscular mycorrhizal (AM) fungi associated with Carica papaya L. in agro-based ecosystem of Goa. India Archives of Agronomy and Soil Science 56(3):237-263. |
|
|
Khade SW, Rodrigues BF (2009b). Ecology of arbuscular mycorrhizal fungi associated with Carica papaya L. in agro-based ecosystem of Goa, India. Tropical and Subtropical Agroecosystems 10: 369-381. |
|
|
Khade SW, Rodrigues BF (2009). Studies on arbuscular mycorrhisation of papaya. African Crop Science Journal 11(3):155-165. |
|
|
Lim TK (2012). Carica papaya. Edible Medicinal and Non-Medicinal Plants 1:693-717. |
|
|
Marcos V, Arévalo-Galarza L, Jaen-Contreras D, Escamilla-García L, Mora-Aguileraa A, Hernández-Castroc E, Cibrián-Tovar, Téliz-Ortiz D (2011). Effect of Glomus mosseae and Entrophospora colombiana on plant growth, production, and fruit quality of 'Maradol' papaya (Carica papaya L.). Scientia Horticulturae 128(3) :255-260. |
|
|
Marcos V, Arévalo-Galarza L, Jaen-Contreras D, Escamilla-García L, Luna-Esquive G (2020). Quality and storage of papaya fruits from plants inoculated with Glomus mosseae, Revista Mexicana Ciencias Agrícolas 11:5. |
|
|
Mbogne JT, Temegne NC, Hougnandan P, Youmbi E, Tonfack LB, Ntsomboh NG (2015). Biodiversity of arbuscular mycorrhizal fungi of pumpkins (Cucurbita spp.) under the influence of fertilizers in ferralitic soils of Cameroon and Benin. Jounal of Applied. Biology and Biotechnology 3(5):1-10. |
|
|
Nakasone HY, Paull RE (1998). Tropical Fruits. CAB International, Wallingford 5:132-148. |
|
|
N'da DH, Adou YCY, N'guessan KEdouard, Moussa Kone et Sagne Yao Charles (2008). Analyse de la diversité floristique du parc national de la Maraboué, Centre-Ouest de la Côte d'Ivoire, Afrique Science 4(3):552-579 |
|
|
Ngonkeu MEL (2009). Tolérance de certaines variétés de maïs aux sols à toxicité aluminique et manganique du Cameroun et diversités moléculaire et fonctionnelle des mycorhizes à arbuscules. Thèse de Doctorat Ph/ D. Université de Yaoundé I. 255. |
|
|
Ngonkeu MEL, Nwaga D, Adamou S, Fokom R, Tchameni NS, Onguene NA, Nana WL, Chaintreuil C, The C, Amougou A, Moulin L, Prin Y, Lebrun M, Dreyfus B (2013). Diversité des Champignons Mycorhiziens Arbusculaires du Cameroun pp. 141-154. |
|
|
Oehl F, Sieverding E, Ineichen K, Mader P, Boller T, Wiemken A (2003). Impact of land use intensity on the species diversity of arbuscular mycorrhizal fungi in agroecosystems of central Europe. Applied and environmental microbiology 69(5) :2816-2824. |
|
|
Phillips JM, Hayman DS (1970). Improved procedures for clearing roots and staining parasitic and vesicular- arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions of the British Mycological Society 55(1):158-161. |
|
|
Pielou EC (1966). The measurement of diversity in different types of biological collections. Journal of Theoretical Biology 13:131-144. |
|
|
Powell JR, Rillig MC (2018). Biodiversity of arbuscular mycorrhizal fungi and ecosystem function. New Phytologist 220(4):1059-1075. |
|
|
Redecker D, Schüßler A, Stockinger H, Stürmer S, Morton J, Walker C (2013). An evidence-based consensus for the classification of arbuscular mycorrhizal fungi (Glomeromycota). Mycorrhiza 23(7):515-531. |
|
|
Sadhana B (2014). Arbuscular Mycorrhizal Fungi (AMF) as a Biofertilizer a Review. International Journal of Current Microbiology and Applied Sciences 3(4):384-400. |
|
|
Samba-Mbaye RT, Anoir CM, Diouf D, Kane A, Diop I, Assigbete K, Tendeng P, Sylla SN (2020). Diversity of arbuscular mycorrhizal fungi (AMF) and soils potential infectivity of Vachellia nilotica (L.) P.J.H. Hurter & Mabb. rhizosphere in Senegalese salt-affected soils. African Journal of Biotechnology 19(7):487-499. |
|
|
Sankaralingam S, Kathiresan D, Harinathan B, Palpperumal S, Shankar T, Ramya B, Prabhu D (2016). Effect of arbuscular mycorrhizae fungi on growth and development of Carica papaya. Scientific Journal of Seoul Sciences 4(1):1-7. |
|
|
Santana FL, Inada CA, Spontoni do Espirito Santo BL, Wander FO, Arnildo A, Alves FM, Guimarães RCA, Freitas KC, Hiane PA (2019). Nutraceutical Potential of Carica papaya in Metabolic Syndrome. National Library of Medicine 11(7):1608. |
|
|
Sas-Paszt L, Sumorok B, Grzyb Z, G?uszek S, Lisek A, Derkowska E, Trzci?ski P, St?pie? W, Przyby? M, Fr?c M, Górnik K (2020). Effect of microbiologically enriched fertilizers on the yielding of strawberry plants in container-based cultivation at different levels of irrigation. Research Institute of Horticulture 1(3):96-100. |
|
|
Schenck NC, Perez Y (1990). Manual for identification of VA Mycorrhizal fungi Gainesville, USA: INVAM, University of Florida 241 p. |
|
|
Schüβler A, Walker C (2010). The Glomeromycota: A species list with new families The Royal Botanic Garden, Edinburgh P. 59. |
|
|
Scullion J, Eason WR, Scott EP (1998). The effectivity of arbuscular mycorrhizal fungi from high input conventional and organic grassland and grass-arable rotations. Plant Soil 204(2):243-254. |
|
|
Sellal Z, Touhami AO, Chliyeh M, Dahmani J, Benkirane R, Douira A (2016). Arbuscular Mycorrhizal fungi species associated with rhizosphere of Argania spinosa (L.) Skeels in Morocco. International Journal of Pure Applied Bioscience 4(1):82-99. |
|
|
Shannon-wiener CE (1948). A mathematical theory for communication. The Bell System Technical Journal 27(4):379-423, 623-656. |
|
|
Sieverding E (1983). Manual de Métodos para la Investigación de la Micorriza Versículo Arbuscular en el Laboratorio. Centro Internacional de Agricultura Tropical, Colombia. |
|
|
Simon L, Lalonde M, Bruns TD (1992). Specific amplification of 18S fungal ribosomal genes from vesicular-arbuscular endomycorrhizal fungi colonizing roots. Applied and Environmental Microbiology 58(1):291-295. |
|
|
Simpson EH (1949). Measurement of diversity. Nature 163:688. |
|
|
Smith SE, Read DJ (2008). Mycorrhizal symbiosis. Amsterdam: Academic Press 787 p. |
|
|
Song J, Chen L, Chen F, Ye J (2019). Edaphic and host plant factors are linked to thecomposition of arbuscular mycorrhizal fungal communities in the root zone of endangered Ulmus chenmoui Cheng in China. Ecology and Evolution 9:900-8910. |
|
|
Spatafora JW, Chang Y, Benny GL, Lazarus K, Smith ME, Berbee ML (2016). A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 108(5):1028-1046. |
|
|
Sukhada M (1989). Response of papaya (Carica papaya L.) to VAM fungal inoculation. In: Mycorrhizae for Green Asia pp. 260-261. |
|
|
Teixeira AF, Kemmelmeier K, Marascalchi MN, Stürmer SL, Carneiro MAC, Moreira FM (2017). Arbuscular mycorrhizal fungal communities in an iron mining area and its surroundings: Inoculum potential, density, and diversity of spores related to soil properties. Ciência e Agrotecnologia 41(5):511-525. |
|
|
Temegne NC, Wakem GA, Taffouo VD, Mbogne TJ, Onguene AN, Youmbi E (2017). Effect of phosphorus fertilization on arbuscular mycorrhizal fungi in the Bambara groundnut rhizosphere. African Journal of Microbiology Research 11(37):1399-1410. |
|
|
Trindade AV, Siqueira JO, Stürmer SL (2006). Arbuscular mycorrhizalfungi in papaya plantations of Espírito Santo and Bahia, Brazil. Brazilian Journal of Microbiology 37:283-289 |
|
|
Trouvelot A, Kough JL, Gianinazzi-Pearson V (1986). Mesure du taux de mycorhization VA d'un système radiculaire. Recherches et méthodes d'estimation ayant une signification fonctionnelle. In. INRA (ed.). Aspects physiologiques et génétiques des mycorhizes. INRA, Paris pp. 217-221. |
|
|
United Nations (2020). United Nations Human Rights Report 2020. |
|
|
Vieira LC, da Silva DKA, Escobar IEC, da Silva JM, de Moura IA, Oehl F, da Silva GA (2020).Changes in an arbuscular mycorrhizal fungi community along an environmental gradient Plants 9:52. |
|
|
Wang JW, Wang GG, Zhang B, Yuan Z, Fu Z, Yuan Y, Zhu L, Ma S, Zhang (2019). Arbuscular mycorrhizal fungi associated with tree species in a planted forest of eastern China. Forests 10(5):424. |
|
|
White TJ, Bruns T, Lee S, Taylor J (1990). Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR protocols, a guide to methods and applications, Academic Press, San Diego pp. 315-322. |
|
|
Wubet TM, Weiss I, Kottke D, Teketay F, Oberwinkler (2004). Molecular diversity of arbuscular mycorrhizal fungi in Prunus africana, an endangered medicinal tree species in dry Afromontane forests of Ethiopia. New Phytologist 161(2):517-528. |
|
|
Zhao ZW, Xia YM, Qin XZ, Li XW, Cheng LZ, Sha T, Wang GH (2001). Arbuscular mycorrhizal status of plants and the spore density of arbuscular mycorrhizal fungi in the tropical rain forest of Xishuangbanna, southwest China. Agriculture Automation and Control 3(11):159-162. |
|
|
Zhu X, Yang W, Song F, Li X (2020). Diversity and composition of arbuscular mycorrhizal fungal communities in the cropland black soils of China. Global Ecology and Conservation 22:e00964. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


