ABSTRACT
Barley ranks third after durum and bread wheat grain production in Algeria. In this study, genetic diversity of twenty nine traditional genotypes of barley was assessed using twenty seven pheno-morphological and agronomic traits in presence of four controls. Very high significant differences were found between genotypes for all quantitative characters statistically analyzed, indicating existence of a great variability within the germplasm. Some traditional genotypes differed from all controls by their better mean values of 1000 grain weight, tiller number per spike, grain number per spike and grain protein content. The long-cycle genotypes had higher values of 1000 grain weight, awn length and plant height than those with short cycle. For quantitative traits, the principle component analysis showed that three components could describe 68.27% of total variances and the cluster analysis divided all six-rowed barley studied into three cluster groups. The following traits: 1000 grain weight, awn length, days to heading and maturity, grain width, grain number per spike, plant height, length of first rachis segment and grain length were those contributing more to variability among the genotypes and also but less strongly the tiller spike number per plant and the spike length. The awn length, the 1000 grain weight, the grain width and the plant height were positively correlated with highly significant correlations between the majority of them. On the qualitative traits, the growth habit, the curvature of the first rachis segment and the lower leaf sheath hairiness were the traits which varied more between all genotypes.
Key words: Qualitative traits, quantitative traits, variability.
Barley is among the most important cereal in the world. It is one of the most ancient crops among the cereals and has played a significant role in the development of agriculture (Ullrich, 2011).
In Algeria and at the beginning of the nineteenth century, barley came at the head of cultures by its importance; it was intended for human consumption and was used as fodder complement. At present, barley ranks third in
Algeria from the point of view growing area and production. It represents currently the main food of sheep.
The introduction during years 1965-1970 of seeds and plantations of varieties with high genetic potential caused the regression of some traditional cultivars. The new varieties of barley adopted in Algeria remain marginal because of their bigger sensibility at the climatic variations.
Jilal (2011) indicates that throughout time, landraces recognized as valuable sources of resistance to pest, diseases and abiotic stresses have been increasingly replaced by commercial cultivars. According to Lasa et al. (2001), specific adaptation traits exist in autochthonous barley and have not been totally incorporated into modern cultivars. Evaluating genetic diversity in cultivated plants for plant breeding programs and heritable resources protection has a vital usage (Khajavi et al., 2014). Determining the level of variation within and among barley populations is an essential step towards conserving genetic resources and developing future strategies for plant improvement (Backes et al., 2009).
Genetic diversity among and within plant species is in danger of being reduced (Eshghi and Akhundova, 2010). Some studies on genetic diversity of barley have focused on phenological and quantitative traits (Assefa and Labuschagne, 2004; Manjunatha et al., 2007; Shakhatrech et al., 2010; Mekonnon et al., 2015). Diversity in barley based on agronomic traits was measured by several authors (Eshghi and Akhundova, 2010; Muhe and Assefa, 2011; Amabile et al., 2013). According to Eshghi and Akhundova (2010), many authors showed that grain yield is an ultimate product of the action and interaction of number of components such as number of tillers, number of grains per spike, 1000-grain weight, plant height, harvest index and etc.
Few works exist on the local resources of cultivated barley in Algeria. On the species Hordeum vulgare, studies concerned generally the local approved varieties "Saïda" and "Tichedrett" and hence the diversity within the local materiel is not known.
This work was done with the objectives to conduct the morphological characterization and to estimate the extent and variability in pheno-morpho-agronomic traits on the barley genotypes of Southern Algeria.
The germoplasm studied on barley consisted of twenty nine genotypes coming from various regions of Algeria. The collection sites were divided into western, central and eastern regions of Algerian Southern. 11 genotypes were recovered from ICARDA (Syria). They concerned the following regions: Biskra with five Genotypes (1, 7, 9, 10 and 16); El Bayadh (8 and 13); Ouargla (11 and 12) and Bechar (14 and 15). Other genotypes were collected by researchers of INRAA within the following regions: Adrar (2, 4, 20, 21, 22 and 24); Touggourt (17, 18, 19, 25, 26 and 27); Ghardaïa (5) and Tamanrasset (28, 29, 30, 31 and 32). All the genotypes were six-rowed barley except the Genotype (14) from Bechar which was barley with 2 rows.
Tamanrasset is located in the Central Sahara and Adrar in South-Western of it. Touggourt, Biskra and Ouargla are regions of the Low Sahara which is a geographic region of north eastern Sahara of Algeria. Bechar is a region of southwest of Algeria. El Bayadh is located on high steppe plains of the southwest of Algeria. Ghardaïa is a part of M’zab which is located between the Low Sahara and the great western erg.
The study was conducted at the Institute of Algerian Agricultural Research of Baraki (Algiers) situated in the plain of Mitidja (with an average rainfall exceeding 500 mm). Planting occurred on 8 December in 2011. Genotypes were evaluated in a sampling random design on three plots in presence of four controls Barberousse or Genotype 3 (France), Pane or Genotype 6 (Spain) and two approved Algerian varieties Saïda (23) and Tichedrett (33). Lines were 4.80 m each, spaced out by 40 cm. With 25 seeds by line, the distance between plants was 20 cm.
Plant height (HPL) (cm), spike length (HEP) (cm), awn length (LBA) (cm), first rachis segment length (LPA) (mm), lower lemma length in sterile spikelet (LGI) (mm), tiller spike number per plant (NTE), grain number per spike (NGE), spikelet number per spike (NEE), days to heading (DEP), days to maturity (DC), 1000 grain weight (PMG) (g), grain length (LOG) (mm), grain width (LAG) (mm) and grain protein content (PRO) (%) were the quantitative traits evaluated.
The qualitative traits were: Growth habit, Lower leaf sheath hairiness, port of flag leaf, shape of spike, spike density, lemma awn barbs, pigmentation of awn tips, glume length compared to grain, lemma type in grain, shape of lemma type in sterile spikelet, curvature of first rachis segment, rachilla hair type and disposition of lodicules in the fertile grain. A random selection of 30 plants was considered to study these following characters: HPL, HEP, LBA, LPA, LGI, NTE, NGE, NEE, LOG and LAG (ten per plot). Also, to determine the degree of fluctuation in the qualitative characters within every genotype, 30 observations by genotype were taken for every character. The test was led without irrigation or fertilization. The texture of the soil was a sandy clay loam texture. The studied characters (quantitative and qualitative) were in their majority, those described by IBPGR (1994) and Niane et al. (1999).
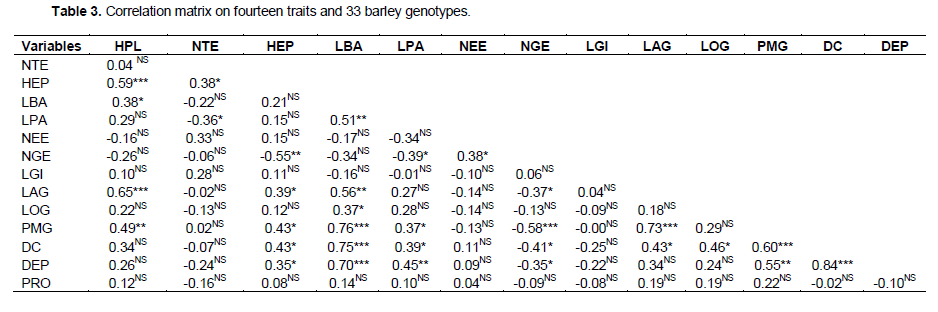
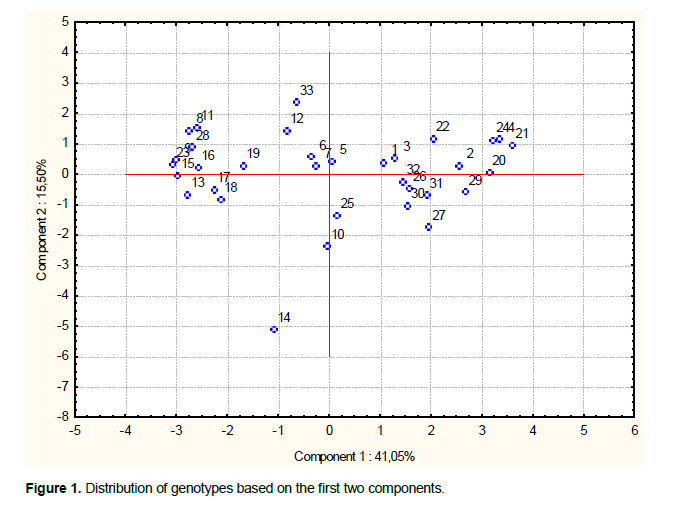
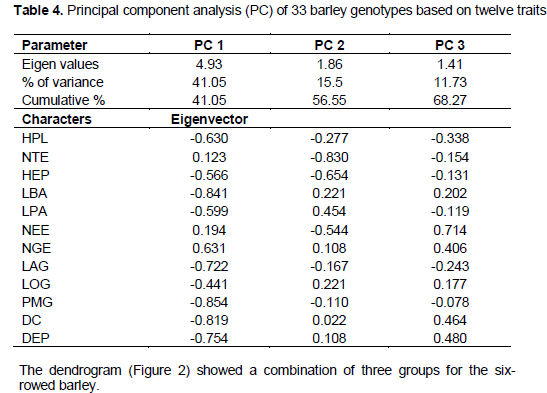
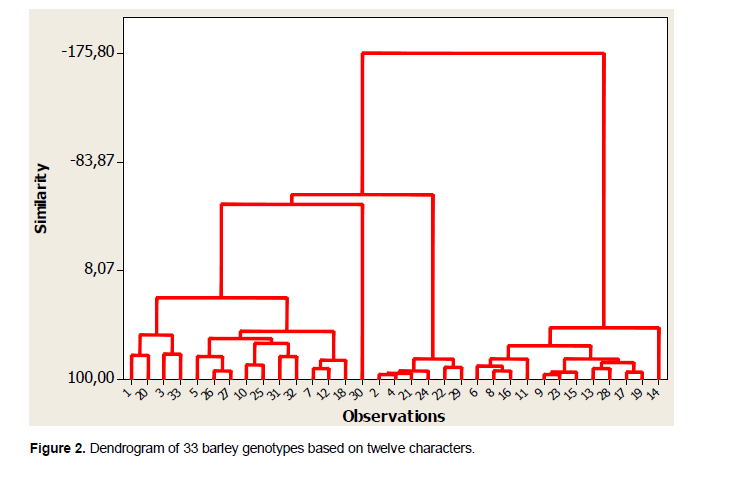
The analysis of variance (ANOVA One-way) was performed by Fisher’s least significant difference (LSD) method to test the significance difference between means. The data was statistically analyzed by the Gen Stat Discovery (Edition 3, Stat Soft Inc.) and concerned only characters with 30 repeated measurements (HPL, NTE, HEP, LBA, LPA, NEE, LGI, NGE, LAG, LOG) (Tables 1 and 2). Correlations (Table 3), principal Component Analysis (Figure 1 and Table 4), cluster analysis (Figure 2) were obtained by STATISTICA (Data analysis Software System, version 8, Stat Soft Inc.). Correlations were performed based on fourteen quantitative characters (HPL, NTE, HEP, LBA, LPA, NEE, LGI, NGE, LAG, LOG, PMG, DEP, DC and PRO). Principal component and cluster analyzes were taken on: HPL, NTE, HEP, LBA, LPA, NEE, NGE, LAG, LOG, PMG, DEP and DC.
The cluster analysis was adopted with the Ward’s method as a clustering algorithm (Ward, 1963). The grain protein content was determined from the nitrogen content, tested by Kjeldahl method (AFNOR, 1985). It is expressed in percentage by weight referred to dry matter.
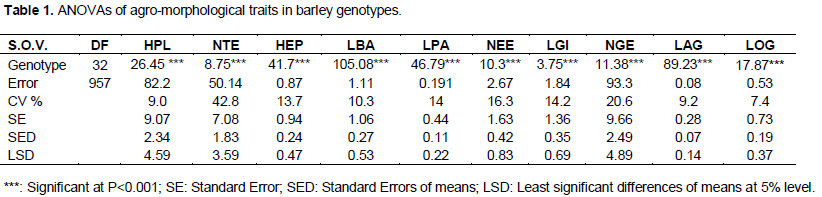
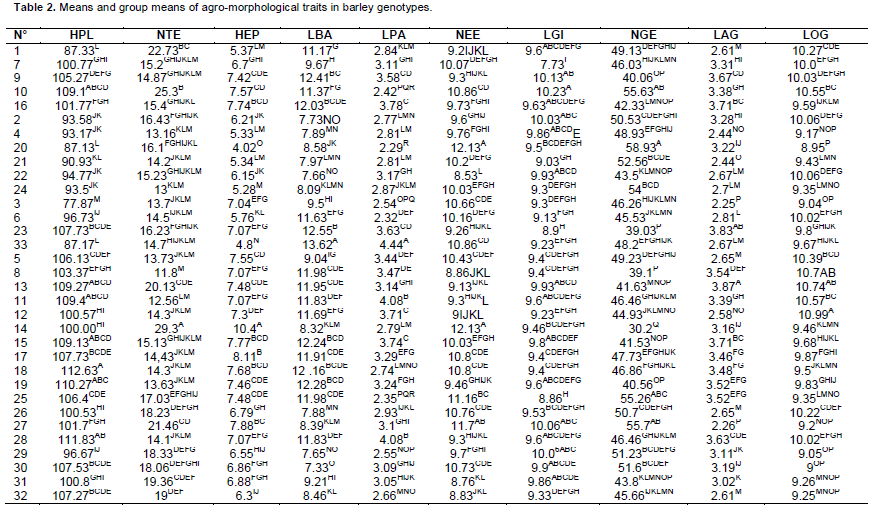
Very highly significant differences were found between genotypes for all quantitative traits statistically analyzed, thus indicating a strong genetic variability within the germplasm (Tables 1 and 2). The distinction between most of the genotypes was marked by at least one character (a separate group). The most distinct genotype is from Adrar (20) at which separate groups are noted on six characters (Table 2). The two-rowed barley (14) also distinguished by its highest values ??(group A) for characters: tillers spike number per plant (NTE), spike length (HEP), spikelet number per spike (NEE) and by the lowest average (group Q) in the grain number per spike (NGE). Seventeen genotypes gave tiller spike number per plant more than fifteen, exceeding “Pane”, “Barberousse” and “Tichedrett (Table 2).
Barley with two rows produced more tillers than all six-rowed genotypes (29.3 tillers). Ten (10) genotypes gave a grain number per spike greater than 50, exceeding the four controls (Table 2). The highest value was registered at the genotype coming from Adrar (20) with 58.93 grains per spike. Seventeen genotypes gave a NGE greater than 40 and the three controls (Tichedrett, Pane and Barberousse). The lowest value of NGE was recorded to barley with two rows (30.2). Nine genotypes gave a 1000 grain weight (PMG) greater than 50 g and also witnesses “Pane” and “Saïda”. With “Tichedrett”, five genotypes gave a PMG more than 40 g.
The better mean value of 1000 grain weight was registered at the Genotype 9 coming from Biskra with 60.9 g. The lowest value concerned 26 from Touggourt with 30 g. Twelve genotypes gave grain protein content exceeding 10% and thus better than controls "Pané" and "Barberousse (9.3 and 8.9% respectively). The highest value was 12.12% and concerned the Genotype 17 (from Touggourt), followed by Genotypes 2 and 21 (from Adrar) with 11.5 and 11.19%, respectively.
The correlation matrix (Table 3) revealed several significant correlations. All the following characters are correlated between themselves: HPL, LBA, PMG and LAG. Days to heading and days to maturity were correlated positively and significantly with LBA, PMG and LPA but significantly and negatively with NGE. The grain width and the grain length were correlated positively with days to maturity. A positive and very highly significant correlation was registered between HPL and HEP. NGE and PMG were highly correlated by negatively. A positive and significant correlation existed between LPA and PMG. There were not significant correlations between the grain protein content and all other characters. It was the same with the lower lemma length.
Principal component analysis was performed based on twelve characters. There were three Eigen values greater than 1, which determined the choice of the three components (Table 4). First component accounted for 41.05%; the second one for 15.5% and the third one for 11.73%. The first component was associated with: PMG, LBA, DC, DEP, LAG, NGE, HPL, LPA and LOG. The second component was associated with NTE and HEP. The NEE contributed to the third component.
In qualitative traits, the most variation among the genotypes concerned the growth habit, the curvature of the first rachis segment and the lower leaf sheath hairiness.
The study showed the existence of a high genetic variability and a great morphological distinction between the various genotypes for all quantitative traits analyzed. The diversity could be explored as a potential source of traits for crop improvement. A wide variation among genotypes of North Africa’s barley was shown for morphological traits (Ben Naceur et al., 2012). Ahmadizadeh et al. (2011) studying genetic diversity of durum wheat landraces from Iran and Azerbaijan reported highly significant differences among genotypes in all of the morphological traits. Compared with four controls, much of traditional genotypes showed better yield components like that the 1000 grain weight, the tiller spike number per plant, the grain number per spike and also for the protein grain content and are therefore interesting for breeding programs.
The most important characters contributing to the variability were: PMG, LBA, DEP, DC, LAG, NGE, HPL, LPA and LOG which explained 41.05% of variation. NTE, HEP and NEE also explain an important part of variability (with cumulative % of components 2 and 3 of 27.23%). These results agree much with those found by Drikvand et al. (2012) were traits contributing to the most variance concerned awn length, plant height, grain yield, grain number per spike, peduncle length, spike length and 1000 grain weight. Many of these traits have also contributed to the genetic differentiation among accessions of barley at Ambo (Setotaw et al., 2010).
The dendrogram of the evaluated barley genotypes divided all six-rowed barley studied into three cluster groups. In a work of Konichi et al. (1993), six-row barley genotypes investigated were also classified into three groups. It is the same for a study by Dimitrova-Doneva et al. (2014).
The first cluster included genotypes: 6, 8, 16, 11, 9, 23, 15, 13, 28, 17 and 19 characterized by long cycle, high values of 1000 grain weight and high means values of awn length. The second cluster grouped the genotypes: 2, 4, 21, 24, 22 and 29 which were characterized by the less large grains, short heading and cycle, short awns, short plants but by the high values of grain number per spike. The third cluster consisted of: 1, 20, 3, 33, 5, 26, 27, 10, 25, 31, 32, 7, 12, 18 and 30 which are characterized by the highest values of late and generally with the higher values of grain number per spike and spikelet number per spike.
The genotypes with long cycle have the greatest weight of 1000 grain and the highest stems. These results are consistent with those found by several authors (Bouzerzour and Monneveux, 1992; Al-Tabbal and Fraihat, 2012). The 1000 grain weight is very highly correlated but negatively with the grain number per spike. A negative significant correlation between these characters was also found by ?áková and Benková (2004) and Babaiy et al. (2011). The spike length was positively correlated (very highly significant) to plant height. Muhammad et al. (2012) found also a significant positive correlation between these two characters. It was also correlated to days to heading and days to maturity (positively and significantly). Mohtashami (2015) found a positive and significant correlation between the spike length and days to heading but positive and not significant correlation with days to maturity. The number of fertile tillers per plant was positively correlated to the spike length. Babaiy et al. (2011) showed a high positive correlation between these traits. The peduncle length was positively correlated with the 1000 grain weight. Babaiy et al. (2011) also found a same result. The awn length was very highly and positively correlated with 1000 grain weight and with days to heading and days to maturity. In fact, late genotypes have longer awns and heavier grains. Indeed, the role played by the awns in the drought resistance and in the grain filling was reported by several authors (Bort et al., 1994; Merah and Monneveux, 2014).
The largest value in grain protein content was 12.12% and concerned the Genotype 17 from Touggourt. The lowest value was 8.53% (Genotype 13 from El Bayadh). The level of protein in barley is highly variable, ranging from 7 to 25% according to a large USDA study involving over 10 000 genotypes (Ullrich, 2002).
On qualitative traits, the homogeneous characters within individuals of the same genotype were: growth habit, lower leaf sheath hairiness, shape of spike, rachilla hair type and shape of lemma type in sterile spikelet. Those with a little fluctuation were: pigmentation of awn tips, lemma type, disposition of lodicules in the fertile grain and glume length compared to grain. A large fluctuation within individuals of the same genotype concerned curvature of first rachis segment and spike density. The port of flag leaf and the lemma awn barb have moderately fluctuated between individuals of the same genotype. These results are consistent with the conclusions drawn by Argüello (1991). The characters which varied more among genotypes were: the growth habit, the curvature of the first rachis segment and the lower leaf sheath hairiness. In a study taken on wheat by Malik et al. (2014), the growth habit was also found as an important phenotypic marker for genetic variability and germplasm evaluation. For the lemma awn barb, proportion of rough awn was greater than the smooth awn. A similar result was found by Derbew et al. (2013).
These descriptors constitute a substantial database on an unknown germplasm and should be considered for breeding programs. The confirmation of these results and the study of other agronomic parameters and those related to the biotic and abiotic stresses will be used for better identifying this germplasm and better directing works of genetic improvement.
The author(s) have not declared any conflict of interests.
The authors’ sincere thanks goes to their colleagues Allam Abdelkader and Derradji Harek for their contribution to the acquisition of germplasm.
REFERENCES
|
Afnor (1985). Aliments des animaux. Méthodes d'analyses françaises et communautaires, 2ème édition. |
|
|
|
Ahmadizadeh M, Shahbazi H, Valizadeh M, Zaefizadeh M (2011). Genetic diversity of durum wheat landraces using multivariate analysis under normal irrigation and rought stress conditions. Afr. J. Agric. Res. 6(10):2294-2302. |
|
|
|
Al-Tabbal JA, Al-Fraihat AH (2012). Genetic variation, heritability, phenotypic and genotypic correlation studies for yield and yield components in promising barley genotypes. J. Agric. Sci. 4(3):193-210. |
|
|
|
Argüello G (1991). Caractères taxonomiques pour la description des variétés d'orge (Hordeum vulgare L. sensu lato). Cours International, Alger du 07 au 25 / 11 / 1992, organisé par : CIHEAM-IAMZ; OAIC; ICARDA; UPOV. |
|
|
|
Babaiy AH, Aharizad S, Mohammadi A, Yarnia M (2011) Survey, correlation of yield and yield components in 40 lines barley (Hordeum vulgare L.) in Region Tabriz. Middle. East. J. Sci. Res. (MEJSR). 10(2):149-152. |
|
|
Backes G, Orabi J, Wolday A, Yahiaoui A, Jahoor A (2009). High genetic diversity revealed in barley collected from small-scale farmer's fields in Eritrea. Genet. Resour. Crop. Ev. 56:85-97.
Crossref |
|
|
Ben Naceur A, Chaabane R, El-Faleh M, Abdelly C, Ramla D, Nada A, Sakr, M, Ben Naceur M (2012). Genetic diversity analysis of North Africa's barley using SSR markers. J. Genet. Eng. Biotechnol. 10:13-21.
Crossref |
|
|
Bort J, Febrero A, Amaro T, Araus JL (1994). Role of awns in ear water-use efficiency and grain weight in barley. Agron. J. 14(2):133-139.
Crossref |
|
|
|
Bouzerzour H, Monneveux P (1992). Analyses des facteurs de stabilité du rendement de l'orge dans les conditions des hauts plateaux algériens. In Séminaire sur la tolérance à la sécheresse, les colloques 64 (Ed INRA France) France pp. 205-215. |
|
|
|
Derbew S, Mohammed H, Urage E (2013). Phenotypic diversity for qualitative characters of barley (Hordeum vulgare (L.)) landrace collections from Southern Ethiopia. Int. J. Sci. Res. 2(9):34-40. |
|
|
|
Dimitrova-Doneva D, Valcheva D, Vulchev D, Dyulgerova B, Gotcheva M, Popova T (2014). Evaluation of grain yield in advanced lines two-rowed winter barley. Agric. Sci. Technol. 6(2):165-169. |
|
|
Drikvand R, Salahvarzi E, Salahvarzi A, Hossinpour T (2012). Study of genetic diversity among rainfed barley genotypes using ISJ markers and morphological traits. J. Agric. Sci. 4(9):137-144.
Crossref |
|
|
|
IBPGR (1994). Descriptors for Barley (Hordeum vulgare L.). International Plant Genetic Resources Institute. Rome, Italy. |
|
|
|
Jilal A (2011). Assessment of genetically diverse international barley germplasm for development of food product applications. PhD Thesis, Southern Cross University, Lismore, NSW. |
|
|
|
Khajavi A, Aharizad S, Ahmadizadeh M (2014). Genetic diversity of promising lines of barley based on pheno-morphological traits in Ardabil area. I. J. Adv. Biol. Biomed. Res. 2(2):456-462. |
|
|
Konichi T, Yano Y, Fukushima Y (1993). Genetic variation in barley landraces from Bhutan. Genet. Resour. Crop. Ev. 40:33-38.
Crossref |
|
|
Lasa JM, Igartua E, Ciudad FJ, Codesal P, Garcia EV, Gracia MP, Medina B, Romagoza I, Motina-Cano JL, Montoya JL (2001). Morphological and agronomical diversity patterns in the Spanish barley core collection. Hereditas 135:217-225.
Crossref |
|
|
|
Malik R, Sharma H, Sharma I, Kundu S, Verma A, Sheoran S, Kumar R, Chatrath R (2014). Genetic diversity of agro-morphological characters in Indian wheat varieties using GT biplot. Aust. J. Crop Sci. 8(9):1266-1271. |
|
|
Merah O, Monneveux P (2014). Contribution of different organs to grain filling in Durum Wheat under Mediterranean conditions I. Contribution of Post-anthesis Photosynthesis and Remobilization. J. Agron. Crop Sci.
Crossref |
|
|
|
Mohtashami R (2015). The correlation study of important barley agronomic traits and grain yield by path analysis. Biol. Forum. Int. J. 7(1):1211-1219. |
|
|
Muhammad RW, Qayyum A, Liaqat S, Hamza A, Yousaf MM, Ahmad B, Hussain M, Ahsan AN, Nagamine T, Yanagisawa T, Minoda T, Shigematsu O, Tokura K, Katou T (2012). Relationships between quality characteristics in the new two-rowed, hull-less barley cultivar "Yumesakiboshi". JARQ – JPN. Agric. Res. Q. 46(2):151-159.
Crossref |
|
|
|
Niane AA, Madarati AW, Abbas A, Turner MR (1999). Manual of morphological variety description for weat and barley with examples from Syria. ICARDA, Syria. |
|
|
Setotaw TA, Dias Lads, Missio RF (2010). Genetic divergence among barley accessions from Ethiopia. Crop. Breed. Appl. Biotechnol. 10:116-123.
Crossref |
|
|
|
Ullrich SE (2002). Genetics and breeding of barley feed quality. in: Barley Science: Recent Advances from Molecular Biology to Agronomy of Yield and quality (Eds Slafer, G.A., Molina-Cano, J.L., Savin, R., Araus, J.L. and Romagoza, I.). Haworth press, Binghamton, NY. pp. 115-142. |
|
|
|
Ullrich SE (2011). Significance adaptation, production and trade of barley. In Barley: Production, Improvement and Uses (Ed UllrichJohn Wiley & Sons Inc., Ames, IA, USA) pp. 3-13. |
|
|
Ward JH (1963). Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 58:236-244.
Crossref |
|
|
|
Žáková M, Benková M (2004). Genetic diversity of genetic resources of winter barley mainted in the genebank in Slovakia. J. Genet. Plant Breed. (CJGPB), 40:118-126. |
|
|



