Full Length Research Paper

ABSTRACT
Filamentous fungi of the genus Trichoderma are known for their activity as plant growth promoters. The objective of this work was to evaluate the efficiency of the Trichoderma asperellum UFT 201 strain as a plant growth promoter in soybean. The ability of phosphate solubilization, indole acetic acid synthesis and the effect on soybean plant growth under greenhouse conditions were evaluated in four experiments. In vitro and in vivo analyzes were performed. Plant biomass characteristics such as shoot dry mass (SDM), root dry mass (RDM) and total dry mass (TDM) were evaluated. T. asperellum UFT 201 showed a significantly higher concentration (p <0.05) with a phosphate solubilization capacity of 67.8% higher than the positive control treatment (Trichoderma harzianum). The production of indole acetic acid (IAA) by T. asperellum UFT 201 was 26.7% higher than the positive control. The effect on the biomass treated by T. asperellum UFT 201 was higher (p <0.05) than the positive and absolute controls. Thus, inoculation by T. asperellum UFT 201 showed high efficiency as a growth promoter in soybean.
Key words: Phosphate solubilization, indole acetic acid, growth promoter, Glycine max.
INTRODUCTION
Soy (Glycine max L.) is one of the most cultivated grains in the world. For the cultivation of soybean and any other crop, some factors must be considered, being the choice of healthy seeds and the performance of the seed treatment, which aims at maintaining the quality of the plants, that could allow the expression of the maximum potential of the crop (Cunha et al., 2015).
The plant growth promotion by soil microorganisms can be performed by direct and indirect mechanisms. The direct mechanisms can be the production of hormones or another substance analogous to them, which influence the growth or development of the plant (Machado et al., 2011; Zeilinger et al., 2016; Gonçalves et al., 2018), or the supply of their nutritional needs by solubilization of phosphates (Contreras-Cornejo et al., 2016). On the other hand, the indirect mechanisms can be by the action of microorganisms through the suppression of pathogens (Silva et al., 2011; Gava and Menezes, 2012; Saravanakumar et al., 2016).
Various soil microorganisms such as fungi can solubilize different forms of inorganic phosphates. Fungi have been reported as phosphate solubilizers in various works (Kapri and Tewari, 2010). Filamentous fungi of the genus Trichoderma have been the most commonly studied microorganisms as biological control agents of plant diseases and they also have activity as plant growth promoters (Santos et al., 2010; Machado et al., 2011). The influence of Trichoderma species on plant development is broad, which includes beneficial effects on seed germination, seedling emergence, grain growth and yield (Chagas et al., 2016). Kapri and Tewari (2010) highlighted the potential of phosphate solubilization by Trichoderma spp. isolates in culture medium. They evaluated the concentrations of soluble phosphate (μg mL-1) and the significant increase in growth parameters of chickpea (Cicer arietinum) in greenhouse trials. Similar studies were done for crops such as cowpea according to Chagas et al. (2015).
Nowadays, the use of growth-promoting micro-organisms in plants to increase agricultural production will probably be one of the most important tactics in the world. This is due to the emerging demand to reduce dependence on chemical fertilizers and the need to develop sustainable agriculture. The production of low cost bioinoculants formulated with plant growth promoting microorganisms such as Trichoderma species is an alternative to reduce the environmental risks caused by inadequate and sometimes excessive use of inputs and pesticides.
Thus, the objective of this work was to evaluate the efficiency of the Trichoderma asperellum UFT 201 in phosphate solubilization, indole acetic acid synthesis and as a soybean growth promoter.
MATERIALS AND METHODS
Location of experiments
The experiments were conducted in a laboratory and greenhouse at the experimental station of the Federal University of Tocantins, Campus Gurupi, located in the southern region of Tocantins State, Brazil. The geographical coordinates of the experimental station correspond to 11°43'45 "S and 49°04'07''W, with an average altitude of 280 m.
Trichoderma strains and characterization
The T. asperellum UFT 201 strain was obtained from the Microbiology Laboratory of the Federal University of Tocantins, Campus Gurupi. This strain was isolated from Cerrado soil in Tocantins. A preliminary identification was made according to the morphological characteristics based on specialized bibliography and with the aid of an optical microscope. Then, the genetic characterization was performed by sequencing the TEF (translation elongation factor) region and identified by the access codes in GenBank (Table 1). This identification was carried out by Instituto Biológico de São Paulo (Brazil).

Trichoderma harzianum CIB T23 strain was used as a positive control in laboratory experiments and in the second greenhouse experiment (access GenBank EU279989) (Hoyos-Carvajal et al., 2009). This strain was used considering that it is the most commonly used strain in commercial products with active microorganism based on Trichoderma. Also, the commercial bioinoculant ICB Nutrisolo (BioAgritec Ltda), based on T. harzianum, T. asperellum and T. koningiopsis (concentration of 1 x 1011 CFU mL-1, liquid formulation, recommended dose of 300 mL ha-1) was used in experiments three and four under greenhouse conditions. This bioinoculant is registered as a plant growth promoter (MAP Register in RS 12734/10000-4) for lettuce (Lactuca sativa L.).
Phosphate solubilization ability
The strains of T. asperellum UFT 201 and T. harzianum CIB T23 were first cultured in petri plates containing PDA medium (HiMedia: 200 g potato, 20 g dextrose, 15 g agar, 1 L distilled water) at 25±2°C 12 h photoperiod for seven days. Discs of approximately 8.0 mm diameter containing mycelium and spores were removed from the colonies and transferred to Erlenmeyer flasks (250 ml) to evaluate their phosphate solubilization potential in vitro in modified NBRIP medium containing the following ingredients (g L-1): glucose, 10.0; MgCl2.6H2O, 5.0; MgSO4.7H2O, 0.25; KCl, 0.2; (NH4)2SO4, 0.1. 50 ml of K2HPO4 (10%) and 100 ml of CaCl2 (10%) were added to the medium to form an insoluble precipitate of calcium phosphate (CaHPO4).
The quantitative estimation of phosphate solubilization was performed in triplicate in a completely randomized design. On the seventh day of microbial growth, the determination of the soluble phosphorus (P) concentration was performed by the colorimetric method described by Murphy and Riley (1962), by subtracting the soluble P contained in the treatments by the contained in the control sample (phosphate culture medium and without inoculum). 1.0 ml of the reagent, 0.5 ml of the filtered sample and 3.5 ml of ultrapure water (Biosystem) were used for evaluations of each sample. After 20 minutes of reaction, the soluble P was quantified in a spectrophotometer at the wavelength of 725 nm by measuring the absorbance. The standard curve was made from monobasic potassium phosphate and the concentrations calculated in µg mL-1.
Synthesis of indole acetic acid (IAA)
The Trichoderma colonies were transferred to Erlenmeyer flasks (150 ml) containing 25 ml of potato dextrose broth with addition of L-tryptophan (100 mg L-1). The indole acetic acid (IAA) production was performed for 7 days on a rotary shaker (100 rpm) at 26 ± 2°C. The fungal biomass was separated by centrifugation (12.000 rpm for 15 min). Colorimetric analysis of IAA was performed using a portion of the Salkowski reagent (0.5 M L-1 FeCl3 + 35% HClO4), two parts (2 ml) of the fungal supernatant and filling the final volume to 5 ml with ultrapure water (Biosystem). After qualitative verification of IAA in the supernatant (pink staining after 25 minutes of reaction at 26±2°C in the dark), the phytohormone was quantified spectrophotometrically at 530 nm. Concentrations were calculated from a standard curve of IAA (0 to 10 μg mL-1) (Cattelan, 1999).
Greenhouse experiments
After the phosphate solubilization and IAA synthesis tests, the ability of Trichoderma strains as promoters of soybean plant growth under greenhouse conditions was evaluated in four experiments. Positive controls (commercial Trichoderma bioinoculants) were used. The experiments were carried out in black plastic pots of 1.7 L. The pots were filled with soil collected in crop areas at the experimental station of the Federal University of Tocantins, Campus Gurupi.
Soil analysis and fertilization
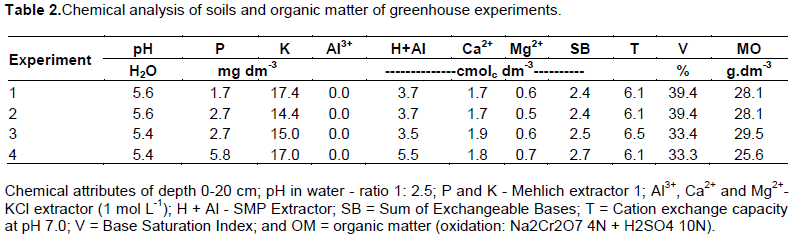
A sample composed of each soil in the different experiments was collected before planting and the chemical characterization of organic matter (OM) was carried out at the Soil Laboratory of the Federal University of Tocantins (Table 2). Soil samples were classified as Dystrophic Red-Yellow Latosol (sieved) and medium texture. It presented the following granulometric characteristics: experiment 1 (72.3% sand, 8.2% silt and 19.5% clay), experiment 2 (70.1% sand, 7.2% silt and 22.7% clay), experiment 3 (72.3% sand, 8.2% silt and 19.5% clay) and experiment 4 (70.5% sand, 7.2% silt and 22.3% clay) (EMBRAPA, 2009). The soils were fertilized using a minimum dose rate of 250 kg ha-1 of formulation 05-25-15 in all experiments.
Preparation of inoculants and soil inoculation
The strains of Trichoderma asperellum UFT 201 and T. harzianum CIB T23 were grown and multiplied on petri dishes containing PDA culture medium. Then, the inoculants were transferred to sterilized millet grains. The millet was weighed, and distilled water was added. Afterwards the grains were transferred to polypropylene bags (300 g of millet / bag with 300 ml of distilled water) and autoclaved at 121°C for 1 h. After cooling, six 5 mm diameter discs containing mycelia, Trichoderma spores and PDA medium were inoculated into the millet with the aid of sterilized forceps. The polypropylene bag was opened in laminar flow camera.
The bag was incubated in camera type Biochemical oxygen demand (B.O.D.) with controlled temperature and light at 25°C and 12 h photoperiod for 14 days. Every two days the millet was stirred to facilitate the gas exchange, the breakdown of the mycelial aggregates and the increase of sporulation. Some millet bags were incubated without Trichoderma inoculation to be used in vessels with the absolute control treatment. After 14 days of incubation, the millet colonized with the Trichoderma strains were weighed at a ratio of 30 g per 1.7 L pot. Thus, the millet was mixed with the sifted soil and placed in the pot. The colonization was carried out for 7 days for subsequent planting of the soybean. Millet without inoculation by Trichoderma was added to the soil in the absolute control.
Samples of 1 g of millet colonized by Trichoderma asperellum UFT 201 and T. harzianum CIB T23 were collected for evaluation of spore counts in Neubauer chamber. Data analysis was done by means of the Trichoderma spore counts, where they had a concentration of 2 × 108 CFU g-1for the test strain T. asperellum UFT 201 and 1.8 × 108 CFU g-1 for T. harzianum CIB T23 as standard strain (positive control).
Design of experiments
The experimental design was completely randomized with four replicates for the four experiments. The experiments are described as follows: (1) Two treatments, one with the inoculation of Trichoderma asperellum UFT 201 and one control without inoculation (absolute control); (2) Three treatments, one with inoculation of Trichoderma asperellum UFT 201, one treatment with inoculation of the standard Trichoderma harzianum strain CIB T23 (positive control) and one treatment without inoculation (absolute control); (3) and (4) Three treatments, one with inoculation of Trichoderma asperellum UFT 201, one with commercial inoculant as positive control and one treatment without inoculation (absolute control). The liquid commercial inoculant was composed by T. harzianum, T. asperellum and T. koningiopsis (concentration of 1 × 1011 CFU mL-1 and recommend dose of 300 mL ha-1).
Treatment of soybean seeds and inoculation
The soybean cultivar M 8644 ipro was used in the first experiment. The soybean cultivar M 8349 ipro was used in the second experiment. The soybean cultivar M 9056 RR was used in the third and fourth experiments. Soybean seeds were initially inoculated with rhizobium (Bradyrhizobium japonicum) in all experiments. SEMIA 5079 and SEMIA 5080 strains for soybean (3 × 109 CFU mL-1) were used. The seeds were sown at a ratio of 10 seeds per pot. The thinning was done between five and seven days after germination and two plants per pot were maintained. The plants were irrigated daily as needed for field capacity maintenance.
Biomass evaluations
The plants were harvested by separating the root system from the aerial part of the plants and the roots were washed in running water to remove the adhered soil. The nodules were removed from the roots and counted. Then, the material was placed for oven drying with forced aeration at 75°C until constant weight.
In the first experiment, two evaluations were made at two separate times, the first at 10 days after plant emergence (DAPE) and the second at 20 DAPE. In the second and third experiment, the evaluations were carried out at 40 days after planting (DAP). The shoot dry mass (SDM), the root dry mass (RDM), the total dry mass (TDM) and the relative efficiency (RE) for the SDM, RDM and TDM were determined. The RE was calculated according to the equation:
RE = (Dry mass inoculated by Trichoderma/Dry mass without inoculation) × 100.
In the fourth experiment, the evaluations were also carried out at 40 days after planting (DAP). The SDM, RDM, TDM, number of nodules (NN), nodules dry mass (NDM) and RE were determined. In addition, the SDM was ground in a knife mill and a sample was taken to evaluate the nutritional status of the plants. The nitrogen (N) contents were determined by the Kjeldahl method. The nitrogen accumulation in shoots (NAS) was calculated by multiplying the mass by the N content.
Statistical analysis
The data were submitted for analysis of variance and test F. The means of the treatments were grouped by the Duncan test at 5% of probability. The statistical software used was Assistant.
RESULTS AND DISCUSSION
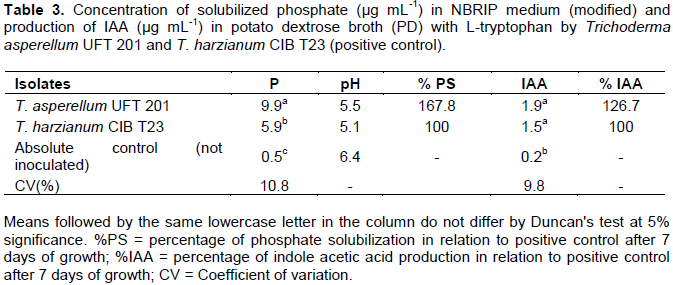
In the laboratory experiments to determine the capacity of phosphate solubilization and synthesis of indole acetic acid (AIA), the treatment with T. asperellum UFT 201 had a phosphorus concentration and a percentage of phosphate solubilization of 67.8% significantly higher (p <0.05) than the positive control treatment (T. harzianum CIB T23) (Table 3). There was a reduction of pH in the culture media with T. asperellum UFT 201 and the positive control in the liquid medium with calcium phosphate. In addition, the Trichoderma isolates used were able to produce indole acetic acid (IAA) in potato dextrose broth supplemented with L-tryptophan, but without significant difference with the positive control. However, treatment with T. asperellum UFT 201 showed an increase in IAA production of 26.7% higher than the positive control T. harzianum CIB T23 (Table 3).
The increase observed in SDM, RDM and TDM variables for the treatment inoculated by T. asperellum UFT 201 in relation to the positive control and absolute control treatments for the soybean plants was probably due to the ability of phosphate solubilization and indole acetic acid synthesis observed for experiment 1 in a greenhouse (Table 3). These results agree with those observed in other agricultural cultures inoculated by specific strains of Trichoderma. According to Hoyos-Carvajal et al. (2009), the increase in biomass production due to inoculation with Trichoderma spp. may be related to the production of growth hormones or analogs. Several species of fungi have been reported to produce auxins, which are fundamental hormones produced by fungi in symbiotic interactions with plants that affect growth and development (Chagas et al., 2015; Contreras-Cornejo et al., 2016).
The inoculant T. asperellum UFT 201 presented IAA production and promoted an increase in the biomass production of soybean plants, indicating a relation between the production of hormones and biomass. The accumulation of biomass in soybean plants may be related to the production of hormones or growth factors, greater efficiency in the use of some nutrients such as phosphorus and the increase of the availability and absorption of this nutrient by plants. Thus, the fungus T. asperellum UFT 201 together with the synthesis or stimulation of phytohormones can acidify the environment where it is established by the secretion of organic acids such as gluconic, citric or fumaric acids (Gómez-Alarcón and Torre, 1994). According to Harman et al. (2004), these acids are the result of the metabolism of the carbon source, mainly glucose, and they can solubilize phosphates, micronutrients and cations including iron, manganese and magnesium. Therefore, the addition of Trichoderma in soils with scarcity of these cations could result in biofertilization by the solubilization of the available metals or the addition of poorly soluble natural phosphates as an alternative of P supplies in the soil. This would result in increased biomass production and crop yields (Contreras-Cornejo et al., 2016).
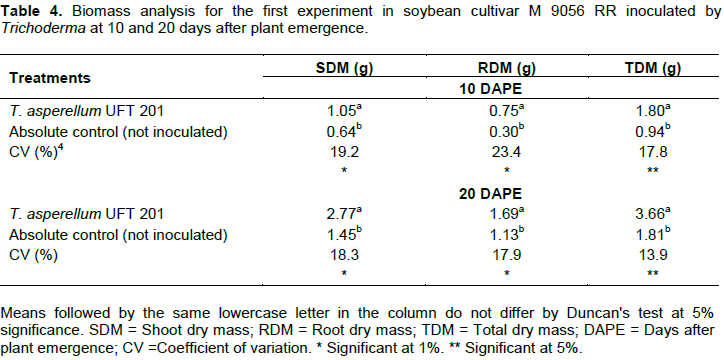
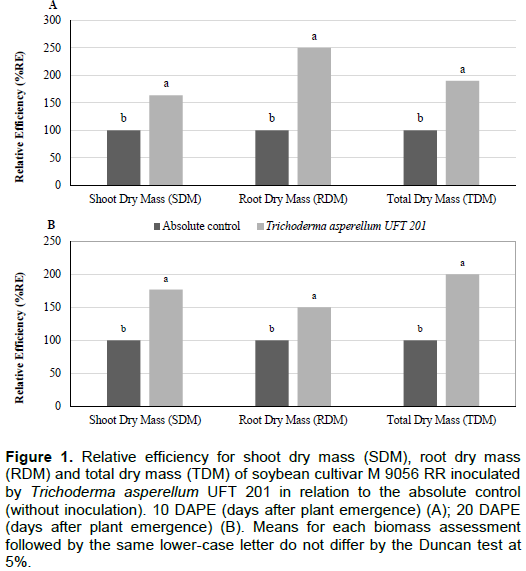
In the greenhouse experiments, for the first experiment, the treatment with the inoculation of T. asperellum UFT 201 was superior than the absolute control (without inoculation) for shoot dry mass (SDM), root dry mass (RDM) and total dry mass (TDM) at 10 and 20 DAPE (Table 4). On the other hand, increased values (p<0.05) of relative efficiency (RE) of 64% in SDM, 150% in RDM and 90% in TDM were observed for the treatment inoculated by T. asperellum UFT 201 at 10 DAPE (Figure 1A). Likewise, Relative efficiency (ER) values for the treatment inoculated by T. asperellum UFT 201 with 77% in SDM, 50% in RDM and 100% in TDM higher than the absolute control at 20 DAPE (Figure 1B).
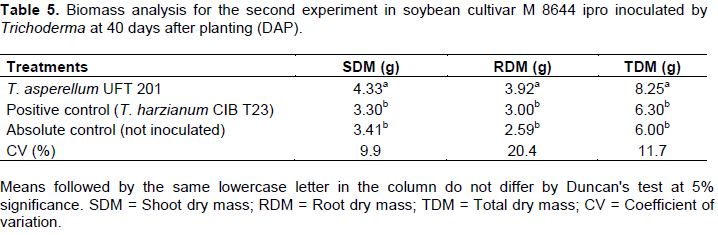
In the second greenhouse experiment, the capacity of plant growth promotion by Trichoderma asperellum UFT 201 was evidenced by the biomass values significantly higher (p <0.05) of SDM, RDM and TDM than the positive control and absolute control at 40 days after planting (DAP) (Table 5). The relative efficiency (ER) of experiment 2 showed higher values (p <0.05) for treatment with T. asperellum UFT 201 with means above 25% than the absolute control and positive control for SDM. Values above 30% and 50% relative efficiency in RDM were detected for T. asperellum UFT 201 compared to the positive and absolute control respectively. Finally, values above 30% relative efficiency in TDM were found for T. asperellum UFT 201 in relation to positive and absolute control (Figure 2).
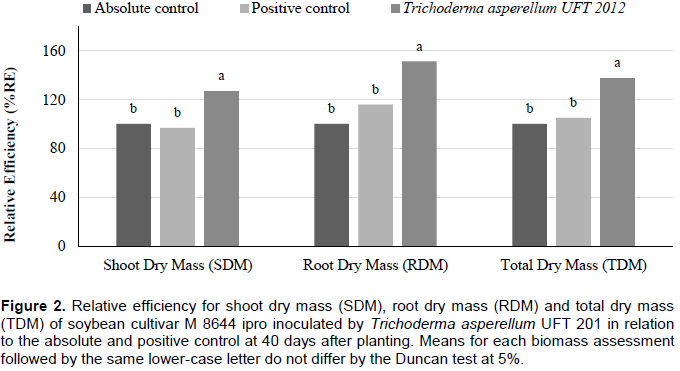
For the third greenhouse experiment, the evaluation of the biomass of treatment inoculated by T. asperellum UFT 201 was significantly higher (p <0.05) than the positive control and absolute control treatments for SDM, RDM and TDM (Table 6).

The relative efficiency (RE) for the biomass evaluations had superior results (p <0.05) for the treatment inoculated by T. asperellum UFT 201 with means above 152, 73 and 102% for SDM, RDM and TDM, respectively; compared to the absolute control. With 67, 22 and 38 for SDM, RDM and TDM, respectively; compared to the positive control (commercial inoculant ICB) (Figure 3). For the fourth greenhouse experiment, the shoot dry mass (SDM), root dry mass (RDM), total dry mass (TDM), nitrogen content (N content) and nitrogen accumulation in shoots (NAS) were significantly higher (p <0.05) for treatment with the inoculation of T. asperellum UFT 201. The number of nodules (NN) and nodules dry mass (NDM) were higher (p <0.05) for treatments inoculated by T. asperellum UFT 201 and positive control than the absolute control without inoculation (Table 7). The relative efficiency evaluations in the treatment inoculated by T. asperellum UFT 201 were highlighted (p <0.05) in relation to the positive control and absolute control with means above 30, 20, and 27% for SDM, RDM and TDM, respectively (Figure 4).
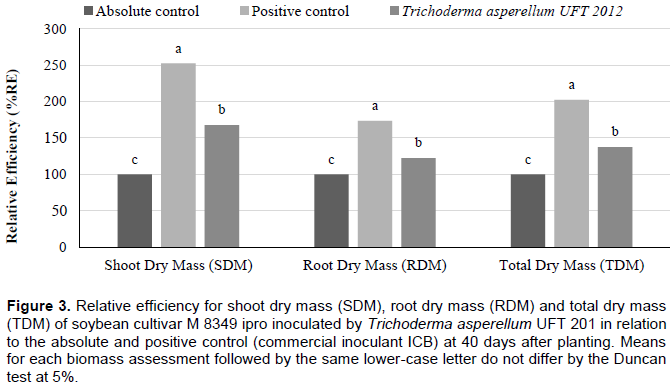
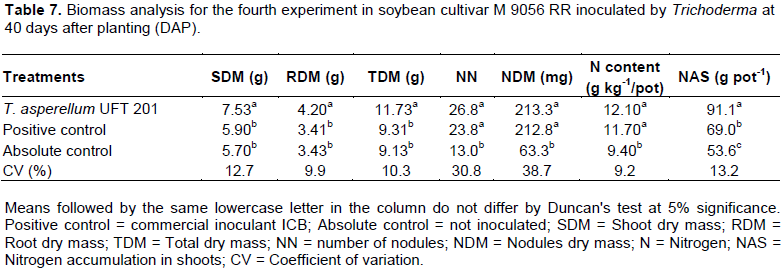
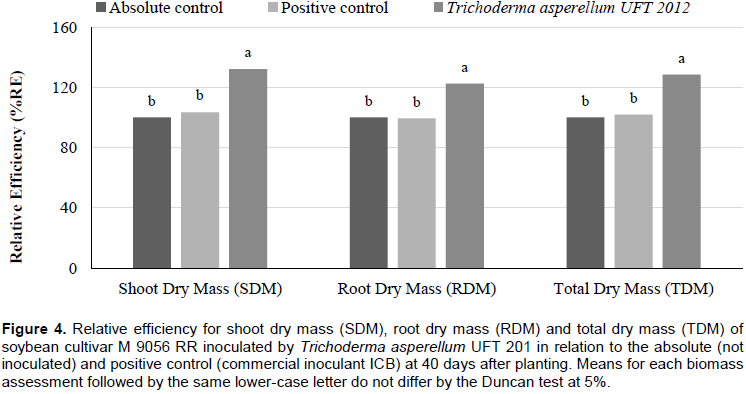
Hence, species of Trichoderma spp. can promote increases of up to 300% in plant growth according to Brotman et al. (2010). The results obtained in greenhouse experiments confirmed the results of Silva et al. (2011),
where they evaluated the effect of Trichoderma on cucumber growth and observed a significant increase in relation to the control without inoculation of Trichoderma. Researchers have been carried out with fungi of the genus Trichoderma spp. Some strains of Trichoderma spp. promote plant growth by increasing nutrient availability and production of growth hormones (Louzada et al., 2009; Chagas et al., 2015). Likewise, the present work could verify the positive results of inoculation by T. asperellum UFT 201.
In all the experiments there was an increase in the biomass of the root system and this increase in the root is related to the sanity of the plant provided by the microorganisms. Plants containing this microorganism associated with their roots or in the rhizosphere tend to have a better ability to survive and absorb nutrients in adverse situations. Consequently, these plants have a productive advantage over those with absence of Trichoderma in their roots (Contreras-Cornejo et al., 2015; 2016).
Rudresh et al. (2005) observed that inoculation with Trichoderma promoted an increase in the growth rate and production parameters of chickpea grown in soil with phosphorus deficiency and fertilized with insoluble rock phosphate under greenhouse and field conditions. These results in the different experiments agree with other works with different agricultural cultures inoculated with different Trichoderma isolates. Santos et al. (2010) concluded that the use of Trichoderma spp. provided positive results in the increase of fresh and dry mass of passion fruit plants from cuttings. Carvalho et al. (2011) evaluated the inoculation of Trichoderma isolates in the promotion of initial common bean growth. Jesus et al. (2011) emphasized the potential of T. asperellum as a substrate conditioner to produce coffee seedlings; they showed a positive effect on the increase of root, shoot and total biomass as well as the increase of the phosphorus absorption efficiency.
Silva et al. (2012) demonstrated that Trichoderma isolates from Amazonian soils increased the biomass of rice plants in greenhouse as well as potential as growth promoters. Machado et al. (2012) also emphasized that
research has proved that Trichoderma spp. is efficient, practical and safe in terms of application methods, biocontrol and plant growth promotion. The results demonstrated the ability of the T. asperellum UFT 201 strain as an active microorganism in granulated formulation (millet) and its efficiency in promoting the initial growth of soybean plants.
CONCLUSION
The isolate T. asperellum UFT 201 solubilized calcium phosphate and synthesized indole acetic acid (IAA). This isolate had a positive effect on the increase of biomass in relation to the absolute control treatment as observed in experiment 1. In addition, the inoculation of T. asperellum UFT 201 presented positive results in the accumulation of biomass (Experiments 2, 3 and 4), nitrogen content and nodulation (Experiment 4). Finally, this isolate showed its potential as an inoculant plant growth promoter in soybean.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Brotman Y, Gupta KJ, Viterbo A (2010). Trichoderma. Current Biology 20:390-391. |
|
|
Carvalho DDC, Mello SCM, Lobo Junior M, Silva MC (2011). Control of Fusarium oxysporum f. sp. phaseoli in vitro and on seeds and growth promotion of common bean in early stages by Trichoderma harzianum. Tropical Plant Pathology 36(1):28-34. |
|
|
Cattelan AJ (1999). Métodos quantitativos para determinação de características bioquímicas e fisiológicas associadas com bactérias promotoras do crescimento vegetal. Londrina: Embrapa Soja 36 p. |
|
|
Chagas LFB, Castro HG, Colonia BSO, Carvalho Filho MR, Miller LO, Chagas Junior AF (2016). Efficiency of the inoculation of Trichoderma asperellum UFT-201 in cowpea production components under growth conditions in field. Revista de Ciências Agrárias 39(3):413-421. |
|
|
Chagas LFB, Castro HG, Colonia BSO, Carvalho Filho MR, Miller LO, Chagas Junior AF (2015). Efficiency of Trichoderma spp. as a growth promoter of cowpea (Vigna unguiculata) and analysis of phosphate solubilization and indole acetic acid synthesis. Brazilian Journal of Botany 38(4):1-11. |
|
|
Contreras-Cornejo HA, Macías-Rodríquez L, Del-Val E, Larsen J (2016). Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: interactions with plants. FEMS Microbiology Ecology 92(4):1-17. |
|
|
Contreras-Cornejo HA, López-Bucio JS, Méndez-Bravo A, Macías-Rodriguéz L, Ramos-Vega M, Guevara-García AA, Lópes-Bucio J (2015). Mitogen-activated protein kinase 6 and ethylene and auxin signaling pathways are involved in Arabidopsis root-system architecture alterations by Trichoderma atroviride. Mol Plant Microbe Interact 28(6):701-710. |
|
|
Cunha RP, Corrêa MF, Schuch LOB, Oliveira RC, Abreu Junior JS, Silva JD, Almeida TL (2015). Diferentes tratamentos de sementes sobre o desenvolvimento de plantas de soja. Revista Ciência Rural 45(10):1761-1767. |
|
|
EMBRAPA (2009). Manual de análises químicas de solos, plantas e fertilizantes. EMBRAPA 2° edição 627 p |
|
|
Gava CAT, Menezes MEL (2012). Eficiência de isolados de Trichoderma spp no controle de patógenos de solo em meloeiro amarelo. Revista Ciência Agronômica 43(4):633-640. |
|
|
Gómez-Alarcón G, De La Torre MA (1994). Mecanismos de corrosión microbiana sobre los materiales pétreos. Microbiología 10(1-2):111-120. |
|
|
Gonçalves AH, Chagas LFB, Santos GR, Fidelis RR, Carvalho Filho MR, Miller LO, Chagas Junior AF (2018). Trichoderme efficiency in the maintenance and productivity of soybean plants in producing savanna regions, Tocantins, Brazil. Revista de Ciências Agrárias 41:175-181. |
|
|
Harman GE, Howell CR, Viterbo A, Chet I, Lorito M (2004). Trichoderma species-opportunistic, avirulent plant symbionts. Nature Reviews Microbiology 2(1):43-56.. |
|
|
Hoyos-Carvajal L, Orduz S, Bissett J (2009). Genetic and metabolic biodiversity of Trichoderma from Colombia and adjacent neotropic regions. Fungal Genetics and Biology 46(9):615-631. |
|
|
Jesus EP, Souza CHE, Pomella AWV, Costa RL, Seixas L, Silva RB (2011). Avaliação do potencial de Trichoderma asperellum como condicionador de substrato para a produção de mudas de café. |
|
|
Kapri A, Tewari L (2010). Phosphate solubilization potential and phosphatase activity of rhizospheric Trichoderma spp. Brazilian Journal of Microbiology 41(3):787-795. |
|
|
Louzada GAS, Carvalho DDC, Mello SCM, Lobo Júnior M, Martins I, Braúna LM (2009). Antagonist potential of Trichoderma spp. from distinct agricultural ecosystems against Sclerotinia sclerotiorum and Fusarium solani. Biota Neotropica 9(3):145-149. |
|
|
Machado DFM, Parzianello FR, Silva ACF da, Antoniolli ZI (2012). Trichoderma no Brasil: O fungo e o bioagente. Revista de Ciências Agrárias 35(1):274-288. |
|
|
Machado RG, De As ELS, Damasceno RG, Hahn L, Almeida D, Morais T, Camargo FAO, Reartes DS (2011). Promoção de crescimento de Lotus corniculatus L. e Avena strigosa Schreb pela inoculação conjunta de Trichoderma harzianum e rizóbio. Ciência e Natura 33(2):111-126. |
|
|
Murphy J, Riley JP (1962). A modified single solution method for determination of phosphate in natural waters. Analytical Chemistry Acta 27:31-36. |
|
|
Rudresh DL, Shivaprakash MK, Prasad RD (2005). Tricalcium phosphate solubilizing abilities of Trichoderma spp. in relation to P uptake and growth and yield parameters of chickpea (Cicer arietinum L.). Canadian Journal Microbiology 51(3):217-222. |
|
|
Samuels GJ, Ismaiel A, Bon MC, De Respinis S, Petrini O (2010). Trichoderma asperellum sensu lato consists of two cryptic species. Mycologia 102(4):944-966. |
|
|
Santos HA, Mello SCM, Peixoto JR (2010). Associação de isolados de Trichoderma spp. e ácido indol-3-butírico (AIB) na promoção de enraizamento de estacas e crescimento de maracujazeiro. Bioscience Journal 26(6):966-972. |
|
|
Saravanakumar K, Yu C, Dou K, Wang M, LI Y, Chen J (2016). Synergistic effect of Trichoderma-derived antifungal metabolites and cell wall degrading enzymes on enhanced biocontrol of Fusarium oxysporum f. sp. Cucumerinum. Biological Control 94:37-46. |
|
|
Silva JC, Torres DB, Lustosa DC, Filippi MCC, Silva GB (2012). Rice sheath blight biocontrol and growth promotion by Trichoderma isolates from the Amazon. Amazonian Journal of Agricultural and Environmental Sciences 55(4):243-250. |
|
|
Silva VN, Guzzo SD, Lucon CMM, Harakava R (2011). Promoção de crescimento e indução de resistência à antracnose por Trichoderma spp. em pepineiro. Pesquisa Agropecuária Brasileira 46(12):1609-1618. |
|
|
Zeilinger S, Gruber S, Bansal R (2016). Secondary metabolism in Trichoderma – Chemistry meets genomics. Fungal Biology Reviews 30(2):74-90. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


