ABSTRACT
Salinity is one of the most common abiotic factors in the world and which strongly limits crop yields. Germination under salt stress could be a quick test to look for salt-tolerant plants. Our study was carried out on the seeds of okra (Abelmoschus esculentus L.), a mallow native to East Africa, known for its fruit used as a vegetable, rich in vitamin C and calcium. The objective of this experiment is to study the effect of salt stress on germination using NaCl at different concentrations. The seeds are germinated at a temperature of 25°C. The results obtained show that the first sprouts appear from the second day after planting seeds for the controls as for stressed seeds salinity; changing germination accelerates with decreasing salt concentration. By cons, salinity does not affect the final rate of germination but rather influences the duration of the germination by lengthening when the medium of the salt concentration increases.
Key words: Abelmoschus esculentus L., tolerance, germination, salt stress, glycophytes.
Climate changes made since ten years are constraints to the growth and development of plants in all regions of the world in general (Higazy et al., 1995) and especially the Mediterranean countries where drought has long been observed, has clearly led to soil salinization process (Gaucher and Burdin, 1974), consequently the first stage of the life of plants namely seed germination conditions its future.
Indeed germination becomes a determining factor for the success of plant growth in saline environments (Khane and Rizvi, 1994). Being a very complex biological phenomenon, germination requires a good understanding of the factors of precocious germination of the seeds that would classify genotypes (Obendorf and Wettanfer, 1984) The duration of germination which is also often cited as a criterion possible effect and the gap between the first and last germinated seeds that affects the success of the plant (Ungar and Badger, 1989).
In order to enhance agricultural production Abelmoschus esculentus L. in Mediterranean regions, it is necessary to better understand the species. Originally from Africa, (Macleod and Ames, 1990), okra is a Mallow, known since the year 1216 B.C. for its fruit used as a vegetable (MACLEOD and AMES, 1990) and all its organs which are of great interests (Grieve, 1984).
To evaluate the response of seeds of this glycophyte, action salinity, we propose initially to study how early germination, then a seed germination kinetics, germination rate and finally the final rate seed germination.
Plant material
The seeds used in germination tests were collected during the month of July 2010 in the region of Nechmaya east of Algeria. Once harvested, the seeds are carefully dusted off and stored in the dark at 5°C for dormancy. After 6 months, the seeds are ready for manipulation. The seeds are disinfected with sodium hypochlorite 1% by soaking them for 3 minutes, then rinsed with distilled water several times to remove traces of chlorine. The seeds used for the germination tests are divided into lots of 10 seeds placed in sterile petri dishes of 10 cm diameter filled with two layers of filter paper.
The Petri dishes are placed in an oven with a thermostat ensuring adequate thermal stability (± 1°C). In each, Petri dishes are paid 7 ml distilled water to control seeds and the same volume of saline for other seeds. Was used three treatments NaCl: to 50 meq.l-1 (2.72 g.l-1), 100 meq.l-1 (5.45 g.l-1) and 200 meq.l-1 (10.9 g.l-1).
In this study, the authors considered that a seed germinated when the radicle had pierced the shell and became visible to the naked eye as defined Come (1970). During the observations, the authors have taken care to imbibe the culture medium, basting as needed. The atmosphere of the oven is kept moist by placing in the bottom of it full of water tray.
At the onset of the tip of the radicle through casings, we proceeded steadily counting sprouts. When the germination rate stabilizes, we completed our observations.
Early germination
In general, each species has a specific germination early to his nature, because even placed in the same experimental conditions the early emergence of the radicle through the membrane will not take place simultaneously in all the seeds (Renard and Quillec, 1975).
This is determined when we see the first sprouts. In this case, early germination is expressed by the rate of the first sprouts corresponding to the time interval between the planting of seeds and raw seeds germinated (Belkhodja, 1996).
Estimated germination
Based on the total number of seeds used (Nt), we calculate the percentage of germinating seeds (Ni) according to the relation: Tg = 100 × Ni / Nt (Tg: germination rate).
Germination time
When the seeds placed in the optimum conditions do not germinate immediately after sowing, are said to represent a germination period (Come, 1970). And germination time is the time (in days) between the first time sprouts and end of germination.
Germination rate
It characterizes the variation in time of the germination rate at the onset of the first peak of the radicle of a seed to the stability of the germination. The authors used the formula of Kotowski (1926) to calculate the coefficient of velocity (Cv) and germination average time (Tm):
Cv = (N1 + N2 + N3 + .... + Nn / N1T1 N2T2 + + + ... N3T3. NnTn
+) x 100
N1T1 Tm = + + N2T2 N3T3 + .... + NnTn / N1 + N2 + N3 + .... + Nn
N1: Number of seeds germinated at time T1; N2: Number of seeds germinated at time T2; N3: Number of seeds germinated at time T3 and Nn: Number of germinated seeds at time Tn.
Germination daily rate
The daily germination rate is calculated based on the number of newly germinated seeds at each observation.
Final germination
This rate is obtained by adding the daily rates of sprouts from the beginning to the end of germination.
Statistical treatment
For the control group and for each of the treatment applied concentrations were taken five repetitions, and obtained experimental data were statistically analyzed by Student's test at the 5% significance level (Snedecor and Cochran, 1984).
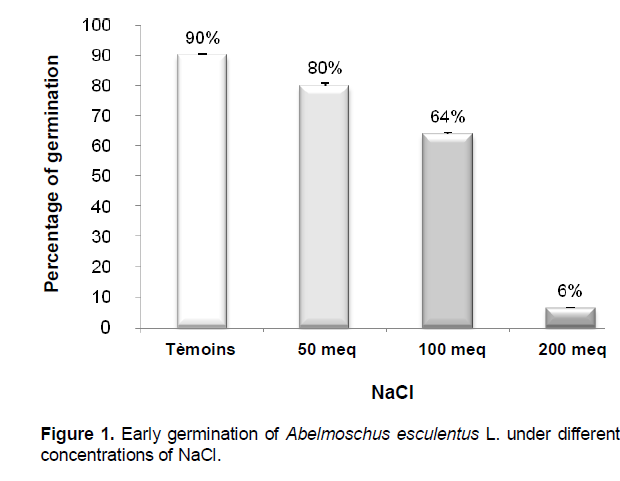
Early germination
Seed germination decreases as the salinity increases in concentration (Figure 1). Indeed, the earliest seeds for sprouting are those soaked in distilled water because the highest rates are achieved (90%) from the 2nd day after sowing.
When the NaCl solution is applied to 50 meq.l-1, the response of the first start their seeds with a germination rate of 80% during the same time. When the concentration of saline twice, the response of seeds is always manifested from the 2nd day but with more reduced rate of 64% for the treatment at 100 meq.l-1 and 6% for the treatment 200 meq.l -1 NaCl.
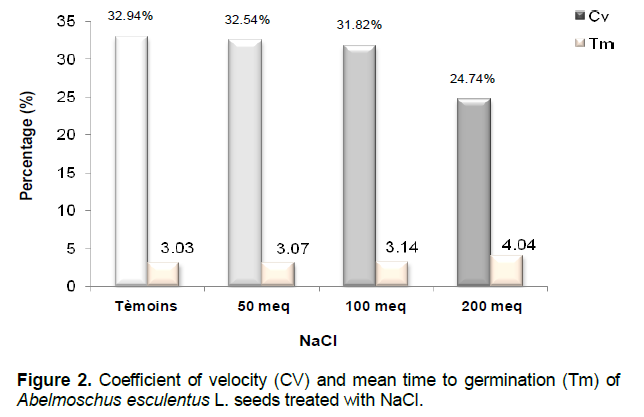
Germination rate
The highest velocity factor is that of control seeds (32.94%). For saline treatments 50 meq.1-1 and 100 meq.l-1 Cv does not very much, but as soon as the double salt concentration, Cv drops to 24.74% (Figure 2).
In terms of the average germination time (Tm), it does not vary much between the different treatments is for control seeds (3.03 days) or treated with 50meq.l-1 (3.07 days) and 100 meq.l-1 NaCl (3.17 days) where it does not exceed three days.When the medium is 200meq.l-1 NaCl; the mean time to germination increases to 4.04 days.
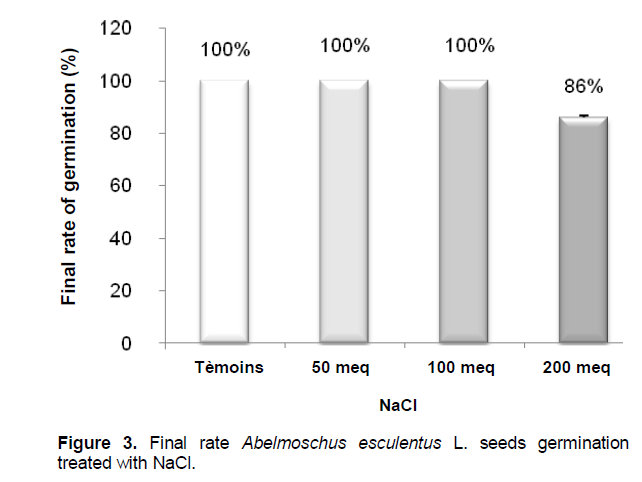
Final rate of germination
The final rate of germination is 100% for control seeds watered with distilled water and sheaths treated with saline 50 and 100 meq.l-1 NaCl (Figure 3). According to the Student test, unlike the germination rate is not significant compared to the control for the seeds watered with saline NaCl 50 and 100 meq.l-1 (100%). However, this difference was highly significant in stressed seeds NaCl 200 meq.l-1 (86% ± 0.6).
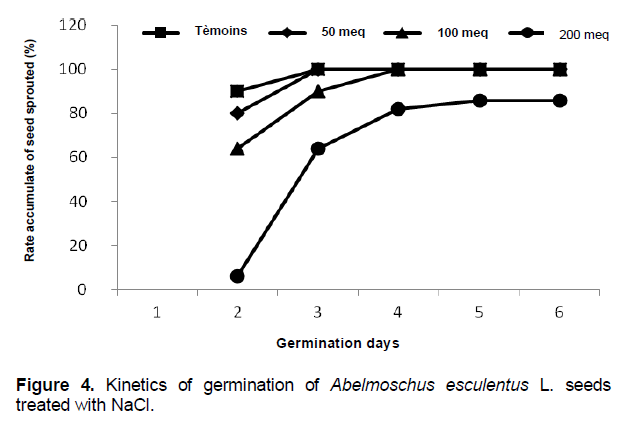
Kinetic of germination
For the control seeds and seeds watered NaCl 50 meq.l-1, the germination starts the second day after sowing, with respective rates of 90% and 80 to 96% for both batches after the 3rd day shown in Figure 4. For the seeds watered 100 meq.l-1 NaCl, germination also starts the second day after seeding, but with a rate of 64% of sprouts Changes in cumulative rates of sprouts three batches overlap and germination rate stabilizes in the 4th with 100% sprouts.
For the saline treatment at 200 meq.l-1 the cumulative rate of germination is progressing very slowly compared to the control batch and that receiving 50 and 100 meq.l-1 NaCl. Germination was completed after 5 days after sowing with a rate of 86% of all the tested seeds.
Changes in the germination of seeds A . esculentus L. recorded in the experimental conditions as described above involve a number of reflection. The final rate of seed germination under different salt treatments did not change much but the speed and germination time varies depending on the treatment applied. Indeed similar results have been observed in many plant species (Demir et al., 2003; Khajeh-Hosseini et al., 2003; Okcu et al., 2005; Kayam et al., 2006). These authors have demonstrated that osmotic potential due to NaCl affects the absorption of water and the mean time to germination but not the final germination. On the other hand, according Askri et al. (2007), the delay caused by the salt is not binding for the final crop yield, agronomic point of view, but rather the germination capacity which is the more decisive. In fact, the germination is not regulated by genotypic characteristics but also by the environmental conditions and in particular, the availability of water in the soil and the presence of salt (Gutterman, 1993).
In halophytes as in glycophytes, salinity reduced germination capacity and delays the initiation process for germination (Bayuelo-Jiménez et al., 2002). However, responses are variable and specific for each species (Ungar, 1991) indeed glycophytes species where most species of agronomic interest its rows has diminished growth in the presence of excessive salinity in the soil generally above 100 meq.l-1 NaCl (Marouf and Reynaud, 2007).
High concentrations of salts, particularly sodium chloride (NaCl), can inhibit the germination of seeds due to osmotic and toxic effects (Perez and Tambelini, 1995). The low external potential can inhibit the enzymatic activity of seeds and delay the release and development of the radicle (Perez and Tambelini, 1995). The Na+ absorption has toxic effects on seed germination, primarily by the disruption of the movement of Ca+2 Na+ In the cell wall which could disrupt its synthesis and, therefore, prevent the growth of rootlets (Zidane et al., 991; Xue et al., 2004; Martinez- Ballesta et al., 2004).
NaCl also affects the permeability of the plasma membrane by increasing the influx of external ions and efflux solutions cytosol (Allen et al., 1995). Furthermore, it increases the rigidity parietal (Nabil and Coudret, 1995) and decreases the fluid conductance of the plasma membrane (Cramer, 1993).
During germination of most seeds, reserves are degraded by amylases, phosphorylases and glucosidase (Bewley and Black, 1994) and much of the hydrolysis products are transported to the embryo for growth (Sun and Henson, 1991).
The slowdown in the mobilization of reserves is due either to delay the activation or synthesis hydrolases or to transfer inhibition hydrolysis products of the endosperm to the embryo. This slowdown in the mobilization of reserves is mainly due to the salinity effect (De-Oliveira et al., 1998).
The results on the influence of salinity on seed germination of A. esculentus L. shows that if the seeds germinate better in the absence of salt (100%) with an average time most current (3.07 days), seeds germinate even at a concentration equal to or less than 100 meq.l-1 NaCl. At high NaCl concentrations to 200 meq.l-1 is slightly inhibited germination (86%) with an average longest time (4.04 days). The average time of germination decreases with the decrease of the NaCl concentration against the germination speed, expressed as velocity coefficient (Cv) decreases with increasing salt concentration. These results suggest that during the seed germination, the species A. esculentus L. behaves as salt tolerant glycophytes.
The authors have not declared any conflict of interest.
REFERENCES
Allen J, Muir SR, Sanders D (1995). Release of Ca2 + from individual plant vacuoles by Both InsP3 and cyclic ADP- ribose. Science 268:735-737.
CrossRef |
|
|
|
Askri H, Rejeb S, Jebari H, Nahdi RH (2007). Effect of sodium chloride on seed germination of three varieties of watermelon (Citrullus lanatus L.). Drought 18:51-55. |
|
|
Bayuelo-Jiménez JS, Craig R, Lynch JP (2002). Salinity tolerance of Phaseolus species falling on germination and early seedling growth. Crop Sci. 42:2184-2192.
CrossRef |
|
|
|
Belkhodja M (1996). Action of salinity on the physiological behavior in the metabolic field beans (Vicia faba L.). Ph.D. thesis, University of Oran. |
|
|
Bewley JD, Black M (1994). Seeds: Physiology of development and germination. New York: Plenum Press.
CrossRef |
|
|
|
Come D (1970). Barriers to germination. Editions Masson. |
|
|
|
Cramer GR (1993). Response of maize (Zea mays L.) to salinity. In: Handbook of Plant and Crop Stress. Mr. Ed Pessarakli, Marcel Dekker, New York pp. 449-459. |
|
|
Demir I, Mavic K, Ozcoban M, Okcu G (2003). Effect of salt stress on germination and seedling growth in serially harvested aubergine (Solanum melongena L.) seeds falling on development . Israel. J. Plant Sci. 51:125-131.
CrossRef |
|
|
|
De Oliveira FA, De- Campos TGS, Oliviera (1998). Effect of saline Subtract on germination, vigor and growth of herbaceous cotton. Engenharia Agricola 18:1-10. |
|
|
|
Gaucher F, Burdin S (1974). Géologie, géomorphologie et hydrologie des terrains salés. Paris: PUF. |
|
|
|
Grieve A (1984). Compared to a perennial and an annual Salicornia Salicornia ecological requirements: Germination and growth of young stages. Oecologia plantarum 8:367-374. |
|
|
Gutterman Y (1993). Strategies of dispersal and germination in plants inhabiting deserts. Bot. Rev. 60:373-425.
CrossRef |
|
|
|
Higazy M, Shehata M, Allam A (1995). Proline relation to salinity of three sugar varieties. Egypt Agric. Res. J. 73:175-89. |
|
|
|
Khane MA, Rizvi Y (1994). Effect of salinity, temperature and growth regulators on the germination on the germination and early seedling growth of Atriplex griffithi var. stocksii. Bot. J. 72:475-479. |
|
|
Khajeh -Hosseini MAA, Bingham Powell IJ (2003). The interaction between salinity stress falling on germination and seed vigor of soybean seeds. Seed Sci. Technol. 31:715-725.
CrossRef |
|
|
Kayam D, Okçu G, Atak M, Çikili Y, Kolsarici Ö (2006). Seed treatments to overcome salt and drought stress falling on germination in sunflower (Helianthus annuus L.). European J. Agron. 24:291-295.
CrossRef |
|
|
|
Kotowski F (1926). temperature relationship to germination of vegetable seeds. Am. Soc. Hortic. Sci. Proc. 23:176-184. |
|
|
|
Macleod G, Ames JM (1990). Volatile components of starfruit. Phytochemistry. Oxford: Pergamon Press. |
|
|
|
Marouf A, Reynaud J (2007). Botany from A to Z. 1662 definitions. Dunod Edition. |
|
|
Martinez-Ballesta MC, Martinez V, Carvajal (2004). Osmotic adjustment, water connections and gas exchange in pepper plants grown under NaCl or KCl. Bot. Environ. 52:161-174.
CrossRef |
|
|
Nabil M, Coudret A (1995). Effects of sodium chloride on growth, tissue elasticity and solute adjustment in two Acacia nilotica subspecies. Physiol. Plantarum 93:217-224.
CrossRef |
|
|
Obendorf RL, Wettanfer HS (1984). Precocious germination falling on in vitro growth of Soybean seeds. Plant Physiol. P. 76.
CrossRef |
|
|
|
Perez SCJGA, Tambelini M (1995). Efeito back Estresses salinos e hydric edo envelhecimento precocious na germinação of algarobeira. Pesquisa Agropec. Bras. Brasília 11:1289-1295. |
|
|
|
Okcu G, Kaya MD, Atak M (2005). Effects of salt and drought stresses on germination and seedling growth of pea (Pisum sativum L.). Turk. J. Agric. 29:237-242. |
|
|
|
Renard JL, Quillec G (1975). The blight of coconut. Preliminary studies. Oilseeds 30:209-213. |
|
|
|
Snedecor GW, Cochran WG (1984). Statistical methods. Paris. Association of Agricultural Technical Coordination edition. |
|
|
Sun Z, Henson CA (1991). A quantitative assessment of the importance of barleyseed a- amylase, P- amylase, debranching enzyme, and a- glucosidase in starch degradation. Arch. Biochem. Biophys. 284:298-305.
CrossRef |
|
|
|
Ungar A (1991). Ecophysiology if vascular halophytes. Boca Raton (Florida): CRC. |
|
|
|
Ungar IA, Badger KS (1989). The effect of salinity and temperature on the germination Inland halophyte Hordeum jubatum. Bot. J. 67:1420-1425. |
|
|
Xue Z, Zhid D, Xue L, Zhang H, Zhao Y, Xia G (2004). Enhanced salt tolerance of transgenic wheat (Triticum aestivum L.) expressing a vacuolar Na+ / H+ antiporter gene with Improved Grain Yields in saline soils in the year has fioed Reduced level of leaf Na+. Plant Sci. 167:849-859.
CrossRef |
|
|
Zidane I, Azaizeh H, Neumann PM (1991). Does salinity REDUCE growth of maize root epidermal cells by inhibiting their capacity for cell wall acidification. Plant Physiol. 93:7-11.
CrossRef |