Full Length Research Paper

ABSTRACT
Plants are frequently exposed to various biotic and abiotic stresses that impair their growth, development and limit the productivity. In the present work the objective was to evaluate the effect of saline stress on the germination and biochemical alterations by the expression of enzymatic systems in corn seeds treated with different concentrations of NaCl. Hybrid maize seeds (H1, H2, H3) were used during 2013 cropping season and solutions of sodium chloride (NaCl) to simulate the saline stress. The respective concentrations of NaCl (mol.m-3) were zero (0.0 g/L); 25 (1.46 g/L); 50 (2.93 g/L); 75 (4.39 g/L) and 100 mol.m-3 (5.85 g/L), diluted into water. In order to evaluate the physiological potential of these seeds, we realized the germination test with four replications of 50 seeds, distributed between three germitest papers moistened with NaCl solutions, described above. The evaluation was performed on the seventh day, with the first count realized determining the percentage of normal seedlings on the fourth day after planting. The biochemical evaluation was performed by the expression of isoenzymes superoxide dismutase (SOD), esterase (EST), catalase (CAT), alcohol dehydrogenase (ADH), malate dehydrogenase (MDH) and alpha amylase. Seeds of the three tested hybrids have lower germination when placed in conditions of greater concentration of salinity. The hybrid H3 that presents higher tolerance to salinity and hybrid H1 are most susceptible during the germination process. The hybrid with higher vigor, H3 had higher activity of alpha amylase and catalase enzyme groups. Esterase and alcoholic dehydrogenase were not effective in measuring the quality of maize grains under salt stress conditions.
Key words: Zea mays, physiological quality enzyme.
INTRODUCTION
Nowadays, maize is the cereal with higher volume of production in the world with approximately 990 million of tons. Brazil is the third higher world producer and the second in number of exportations, with a cultivated area around 17 million of hectares (CONAB, 2014).
Despite the expressive growth in area, production and productivity, the conditions for cultivation of this species, are not always appropriate to the agronomic performance as, the limiting nutrient availability and the occurrence of hydric deficit in the field.
In situations where the integrated fertilization management and irrigation are not done correctly, increases occur in the concentration of salts in the soil making impossible the cultivation of most part of plants (Tôrres et al., 2004). The occurrence of saline soils in arid and semi-arid regions is due mainly to the low precipitation and high evapotranspiration. In these conditions, the salts are not leachate, accumulating in the soil or in the water, being harmful to the normal development of the plants. The high concentration of salt is a factor of stress for the plants, because it reduces the osmotic potential and provides the action of ions on the protoplasm. Besides this, it compromises the germination of seeds and leads to the ionic imbalance on cells and consequently, toxicity and stress (Deuner et al., 2011).
Salt stress affects the germination, as well as the vigor, metabolism and the seeds chemical constitution, being these effects dependent of other factors like; species, cultivar, phenological stages, intensity and the duration of the stress (Tester and Davénport, 2003). Most commonly used method for determining the adaptation and tolerance of different species to these stresses is the percentage of germination in salt substrate. The reduction in the percentage of germination in relation to the control is used as indicative of the level of tolerance of the species to the salinity (Góis et al., 2008).
Internal quality control programs beyond the germination and biochemical tests have been used (Catão et al., 2010). Besides this, researches have associated the electrophoreses technique to the physiological quality of maize seeds, being the molecular markers used like tools in the evaluation of this physiological quality. The selection of genotypes through the previous evaluation of physiological quality of seeds and their enzymatic system can provide parameters capable to help in the evaluation of new maize cultivars tolerant to different stresses (Abreu et al., 2014).
Therefore, the objective of the present work was to evaluate the effect of salt stress on germination and biochemical alterations by the expression of isoenzymatic systems in maize seeds submitted to different concentrations of NaCl.
MATERIALS AND METHODS
This study was conducted at Central Laboratory of Seeds, Department of Agriculture of Universidade Federal de Lavras (UFLA), in Lavras-MG. Maize hybrid seeds described as hybrids 1, 2 and 3 (H1, H2 and H3) were used during the 2013 cropping season. The materials were obtained from the same field under the same irrigation system and the three cited materials have the same male genitor. After the harvest, these seeds were stored in cold chamber at 10°C in multiwall paper bags until realization of the following tests and determinations.
Water content was determined by the oven method at 105°C for 24 h using five replicates of 50 seeds from each treatment (Brasil, 2009). The results were expressed in percentage.
In order to simulate the salt stress, sodium chloride solution (NaCl) was used. The concentrations of NaCl (mol.m-3) were: zero (0.0 g/L); 25 (1.46 g/L); 50 (2.93/L); 75 (4.39 g/L) and 100 mol.m-3 (5.85 g/L) diluted in distilled water. At level zero, distilled water was used to moisten the substrate. The value of electric conductivity of these solutions was verified with aid of a conductivimeter; while Van't Hoff formula according to Salisbury and Ross (1991) was used to calculate the amount of NaCl to be added for the obtainment of each treatment.
In the germination test, for each treatment four replications of 50 seeds were used. Seeds were distributed between three germitest papers moistened with the NaCl solutions previously described. The amount of solution equivalent to 2.5 times the weight of dry paper was used. Following that, the rolls were kept in germinator type B.O.D regulated to the constant temperature of 25ºC. The evaluation was realized at seventh day after the installation of the test by determining the percentage of normal seedlings (Brasil, 2009). The first count was realized together with the germination test, by determining the percentage of normal seedlings at fourth day after the test installation.
The hypocotyl and root length were also determined. In this regard, seeds from each treatment, in five replications of 15, were sown equidistant from each other, according methodologies described to germination test. The measurement of hypocotyls and root length of seedlings was classified with an aid of a graduated ruler. The results were expressed in cm.seedling-1.
In the biochemical evaluation, samples of 10 seeds of each treatment were collected and macerated in presence of PVP (polyvinylpyrrolidone) and liquid nitrogen in small container and afterwards stored at -86ºC temperature. The seeds were collected 48 h after the germination test installation.
For enzymes extraction, the extraction buffer (tris HCl 0.2 M pH 8 + 0.1% of β-mercaptoethanol) was added in the proportion of 250 Ï»L for 100 mg of seeds powder. The material was homogenized in vortex and kept in refrigerator during 12 h followed by the centrifugation at 14000 rpm for 30 min at 4ºC and then applied in polyacrilamide gel. The electrophoretic run was realized in a discontinuous polyacrylamide gel system at 7.5% (separating gel) and 4.5% (concentrating gel) using tris-glycine pH 8.9 as standard buffer in the gel electrode system. In each gel channel, was applied 50 µL of the sample supernatant and the running was performed at 150 V for 5 h. At the end of running, the gels were revealed for the enzymes superoxide dismutase (SOD- EC.1.15.1.1.), catalase (CAT- EC.1.11.1.6.), esterase (EST- EC 3.1.1.1.), malate dehydrogenase (MDH- EC 1.1.1.37.) and alpha amylase (α-AMI- EC 3.2.1.1.), according to the protocols established by Alfenas (2006). Evaluation of the gels was realized on transilluminator, being considered the variation of intensity of bands.
Factorial experiment in randomized complete design of (3x5) was used, including three maize hybrids (H1, H2, H3) and five different salinity levels (0, 25, 50, 75, 100 mols.m-3). The data, previously submitted to the normality tests and homocedasticity of variances, were submitted to analysis of variances and the averages were compared by the Scott-Knott test at 5% of probability. The statistical analyzes were realized with aid of SISVAR® statistical program (Ferreira, 2011). Evaluation of the enzymatic patterns was made according to the intensity of the bands.
RESULTS AND DISCUSSION
The medium water content of seeds in the moment of the test was of 12.3% with maximum variation of 1%. This uniformity between the tested materials is important once accentuated variations accelerates the deterioration process and the formation of products which entail immediate damages, like the free radicals, masking the final result (Marcos Filho, 2005).
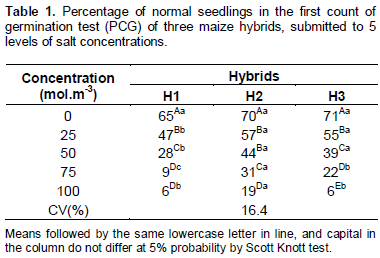
The decrease in water availability caused by the reduction of osmotic potential of saline solutions reduced gradually the percentage of normal seedlings at fourth day of evaluation for all the hybrid seeds accordingly verifying the first count of germination (PGG) (Table 1). For the H1, the vigor reduction is more pronounced in the concentration of 75 mol.m-3, not differing statistically from the 100 mol.m3 concentration. For the H2, the concentrations of 25 and 50 mol.m-3 did not present significant differences. The hybrid seeds H3 had a continous and proportional decline with increasing salt concentration, statistically differing from each concentration at which they were submitted.
In analyzing each salt concentration in different maize hybrids function, it was observed that when the concentration of 0 mol.m-3 was used, the three hybrids did not present significant diferences between them (Table 1). Already in the concentrations of 25 and 50 mol.m-3, the hybrids H2 and H3 had presented higher percentage of normal seedlings in the first count in comparison to the H1. In the concentraion of 75 mol.m-3 all maize hybrids were significantly different between them, with the better performance of the H2, followed by the H3 and H1. In the higher concentration of 100 mol.m-3, the H2 had higher percentage of normal seedlings, while the hybrids H1 and N3 did not differ between them.
The reduction in the osmotic potential induced by treatment with NaCl, caused alterations in the germinative parameters of maize seeds in different intensities. The percentage of germination decreased proportionally to the increase in the osmotic potential with NaCl for all the hybrid seeds (Table 2). In the absence of salt (0 mol.m-3), the percentage of seeds germination achieve levels superior than 95%. Seeds subjected in the 25 mol.m-3 solution recorded percentage of germination equal or superior than 90% and, however, did not differ statistically in the control and between them. The reduction in the percentage of H1 germination, comparing the difference between the control treatment and the higher osmotic potential, was of 56% points, more drastic than the reduction of H2 (36% points) and of H3 (28% points). In the concentration of 50 mol.m-3, H3 presented 86% of germination value above the considered minimum pattern for the commercialization of maize seeds, according to the Ministry of Agriculture, Cattle and Supply (MAPA). This minimum value for the remaining concentrations of H2 and H1 was not verified.
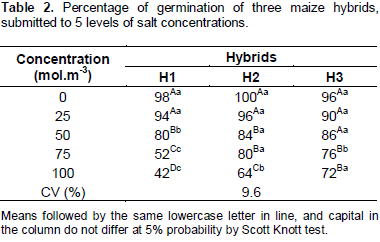
The decrease in the percentage of germination with increasing of the salt stress may be due to the osmotic effect, as well as the effect known as physiological drought; like the toxic effect resulting from the acumula-tion of ions in the protoplasm and the nutritional imbalance occasioned by the inhibition of absorption and transport of nutrients (Tôrres et al., 2004). Similar results were found by Moraes and Menezes (2003) on soybean seeds, confirming the decrease in germination with the reduction of osmotic potential of solutions in the substrate. In rice, Lima et al. (2005) verified decrease in the germination of all the cultivars, in function of the increase in salt concentration, and suggested that the salinity affects the development of normal seedlings and decreases the viability and vigor of seeds.
The root length was less compromised by the water restriction occasioned by the presence of salts, than the development of the shoot part (Tables 3 and 4). These results can be explained by the fact that the seedlings submitted to higher levels of water stress, in general tend to invest higher biomass and to develop higher radicular system as strategy of survival. The distribution of the root system in depth/length on grounds of the water failure is considered as indicative parameter of drought tolerance and can confer adaptation in some species.
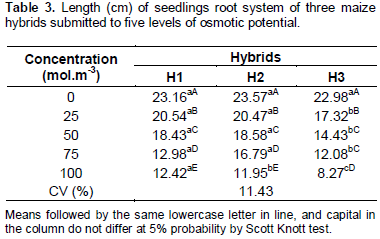
There was no variation between the three hybrids analyzed in the treatment of 0 mol.m-3. In the concentrations of 25, 50, and 75 mol.m-3, the hybrids H1 and H2 presented results statistically superior. In the treatment of 100 mol.m-3, the hybrid H1 had higher length. Beside this, comparing the variations inside each hybrid, it is noted that in all of them occurs reduction of the root length with increase in water stress (Table 3).
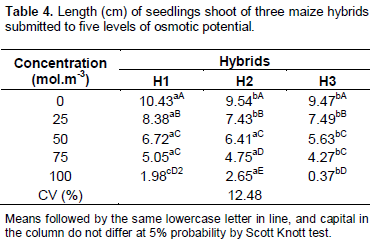
In relation to the data about shoot length (Table 4), it was observed that in the concentration of 25 mol.m-3, the hybrid H1 presented the higher length of shoot in relation to the other hybrids. However, in the concentration of 50 and 75 mol.m-3, H3 had lower shoot length. In the concentration of 100 mol.m-3, the higher length was observed with H2 hybrid. All maize hybrids recorded a reduction in the shoot length according the osmotic potential decreased, mainly due to the reduction of the metabolism occasioned by the lack of water, which harms the plant development.
Generally, the water stress caused damage in the radicular and shoot growth of maize seedlings. Similar results were found in other studies reported by Kaya et al. (2006), Ávila et al. (2007) working with canola, and Conus (2009) on maize, where they observed that salt stress decreased shoot and roots lengths.
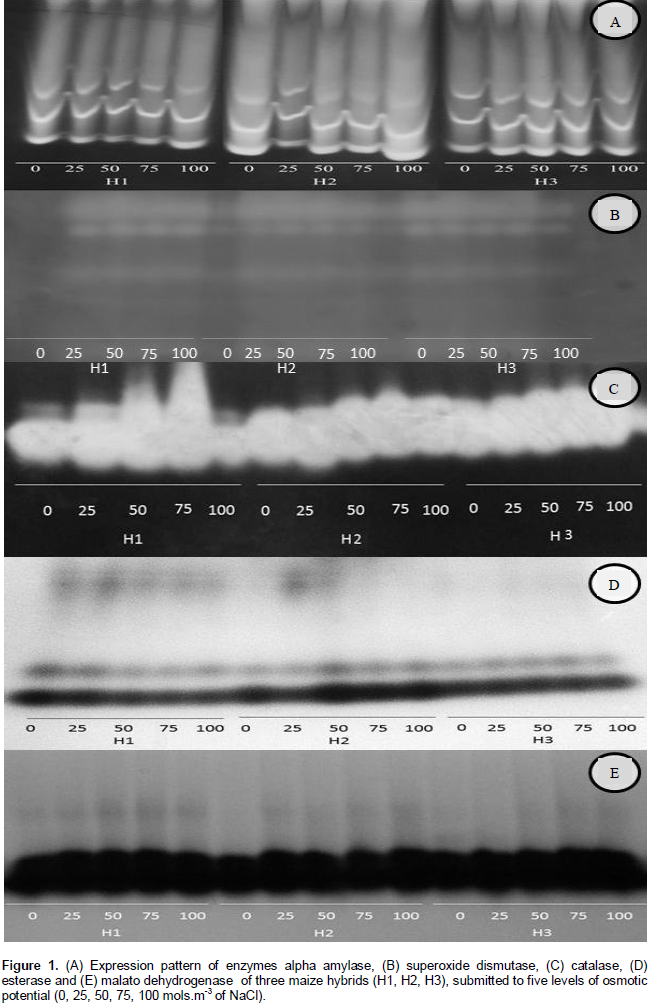
Figure 1 illustrates the expression pattern of enzymes alpha amylase, superoxide dismutase, catalase, esterase and malato desidrogenase in seeds of three maize hybrids submitted to imbibition. The activity of the enzyme alpha amylase can be evidenced by achromatic bands in blue background due to the reaction of the iodine with amylase. In the locals where the enzymes were present, the starch was hydrolyzed. This enzyme has high activity in the beginning of the germinative process and, over time, the activity decreases. This behavior is due to the reduction of starch content while the germination is occurring. Being an enzyme responsible for starch hydrolyses, genotypes which present high levels of alpha amylase expression have higher capacity in providing starch for the embryos, justifying the better performance in the final germination (Franco et al., 2002). However, Abreu et al. (2014) working on maize seeds, claims that water restriction to the seeds (lower availability of water) results in delay of imbibition process. Once the enzymes responsible for starch degradation in maize seeds are “synthesized again” at the moment of imbibition, there will be higher acumulation of alpha amylase enzyme as soon as there is an increase of this water restriction.
In general, it is observed from this study, that H1 presented lower activity of alpha amylase in all saline concentrations, except in the higher concentration of 100 mol.m-3, when compared to the hybrids H2 and H3 (Figure 1A). The same result was observed in relation to the first count of germination and germination (Tables 1 and 2). However, comparing the electrophoretic profile of hybrids H2 and H3, it is observed that the H3 presents lower activity of this enzyme, differently from what was presented in Tables 1 and 2. Besides this, it was observed that in the concentration of 100 mol.m-3, the activity of enzymes is higher for the three hybrids.
The enzyme SOD is directly involved in the celular defense against the free radicals being responsible by the convertion of superoxide radical into hydrogen peroxide. However, it was observed from Figure 1B that there was no increase in the expression of this enzyme with increasing osmotic potential, suggesting that the stress occasioned by the water restriction for the three maize hybrids was not enough for increasing activity of this enzyme.
Another enzyme involved in the removal of hydrogen peroxide, formed from the activity of SOD is the enzyme CAT. The compound hydrogen peroxide is toxic for the seeds but not for the cells, the catalases enzymes act on hydrogen peroxide neutralizing it. The higher activity of CAT for the three hybrids in higher concentrations of stresses (50, 75 and 100 mol.m-3) can be atributed to the activity of this enzyme for removing the hydrogen peroxide accumulated in occasion of saline stress. As verified in Table 2 for H1, the reduction in germination decreased significantly from the potential of 75 mol.m-3 and suggests higher deterioration level of these seeds.
In relation to the enzyme EST, it was observed that in the control treatment (0 mol.m-3) for hybrids H1 and H2, the activity of this enzyme was higher when compared to H3 (Figure 1C). In other concentrations, high activity was observed only for H2. The high activity of this enzyme is related to the higher efficiency of the membrane system, and according to the Tables 1 and 2, the H2 can be observed to present higher quality when compared to H1.
The enzyme MDH is an important molecular marker of seeds respiration. The accumulation of salt in solution and consequently water restriction, take seeds to a condition of lack of oxygen. In Figure 1E, higher expression of this enzyme with increasing osmotic potential was observed; there was also decrease in seeds respiration and increase in the deterioration process.
However, this enzyme is codified for five loci and it is found in great abundance in different cellular organelles, in mitochondria’s and cytoplasm’s. Due to this alteration in your expression, it is only observed when it occurs in seeds, a very large deteriorating process, which makes this enzyme an inefficient marker of physiological quality.
CONCLUSIONS
Seeds of the three maize hybrid tested presented low percentage of germination when placed in conditions of higher salt concentration.
The hybrid with higher vigor, H3, had lower activity of the enzymatic groups alpha amylase and esterase. Superoxide dismutase and malato desidrogenase were not efficient in measuring the quality of maize seeds under salt stress.
The hybrid H3 is the one which presents higher tolerance to the salinity and the hybrid H1 is the most susceptible during the germinative process
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors thank the Foundation for Research Support of the State of Minas Gerais (FAPEMIG), the Coordination for the Improvement of Higher Education Personnel (CAPES), and The National Council for Scientific and Technological Development (CNPq), for financial support and scholarships.
REFERENCES
|
Abreu VM, Von Pinho EVR, Von Pinho RGV, Naves GMF, Neta ICS, Guimarães RM, Carvalho MR (2014). Physiological performance and expression of isozymes in maize seeds subjected to water stress. J. Seed Sci. 36:40-17. |
|
|
Alfenas AC (2006). Eletroforese e marcadores bioquímicos em plantas e microrganismos. Viçosa: UFV. p. 627. |
|
|
Ávila MR, Braccini AL, Scapim CA, Fagliar JR, Santos JL (2007). Influência do estresse hídrico simulado com manitol na germinação de sementes e crescimento de plântulas de canola. Rev. Bras. Sementes 29:98-106. |
|
|
Brasil (2009). Ministério da Agricultura, Pecuária e Abastecimento. Regras para análise de sementes. Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Defesa Agropecuária. Brasília, DF: Mapa/ACS. p. 395. |
|
|
Catão HCRM, Costa FM, Valadares SV, Dourado ER, Brandão Junior DS, Sales NLP (2010). Qualidade física, fisiológica e sanitária de sementes de milho crioula produzidas no norte de Minas Gerais. Ciência Rural 40:2060-2066. |
|
|
Companhia Nacional De Abastecimento- CONAB (2014). Perspectivas para a agropecuária. Brasília 2:1-155. |
|
|
Conus LA, Cardoso PC, Venturoso LR, Scalon SPQ (2009). Germinação de Sementes e Vigor de plântulas de milho submetidas ao estresse salino induzido por diferentes sais. Rev. Bras. Sementes 31:67-74. |
|
|
Deuner C, Maia MS, Deuner S, Almeida AS, Meneghello GE (2011). Viabilidade e atividade antioxidante de sementes de genótipos de feijão-miúdo submetidos ao estresse salino. Rev. Bras. Sementes 33:711-720. |
|
|
Ferreira DF (2011). SISVAR: a computer statistical analysis system. Ciência Agrotecnologia 35:1039-1042. |
|
|
Franco OL, Rigden DJ, Melo FR, Grossi-de-Sá M (2002). Plant alfa amylase inhibitors and their interaction with insect alfa amylase. Eur. J. Biochem. 269:397-412. |
|
|
Góis VA, Torres SB, Pereira RA (2008). Germinação de sementes de maxixe submetidas a estresse salino. Caatinga 21:64-67. http://www.redalyc.org/articulo.oa?id=237117689010 |
|
|
Kaya MD, Okçu G, Mehmet A, Çilkili Y, Kolsarici O (2006). Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Eur. J. Agron. 24:291-295. |
|
|
Lima MGS, Lopes NF, Moraes DM, Abreu CM (2005). Qualidade fisiológica de sementes de arroz submetidas a estresse salino. Rev. Bras. Sementes 27:54-61. |
|
|
Marcos Filho J (2005). Fisiologia de sementes de plantas cultivadas. Piracicaba: Fealq. P 495. |
|
|
Moraes GAF, Menezes NL (2003). Desempenho de sementes de soja sob condições diferentes de potencial osmótico. Ciên. Rural 33:219-226. |
|
|
Salisbury FB, Ross CW (1991). Plant Physiology. 4th ed. California: Wadsworth Publishing Company, Inc. p. 682. |
|
|
Tester M, Davénport R (2003). Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 19:503-507. |
|
|
Tôrres ANL, Pereira PRG, Tôrres JT, Gallotti GJM, Pilati JA, Rebelo JA, Henkels H (2004). A salinidade e suas implicações no cultivo de plantas. Florianópolis: Epagri. p.54 |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


