Full Length Research Paper

ABSTRACT
Rubber tree clones are used for their high latex production, resistance to the diseases and to the climatic conditions. The aim of this study was to characterize the parasitism of Phragmanthera capitata on three rubber clones. The study was carried out in the rubber plantations of the Cameroon Development Corporation located in 4 villages around Muyuka and Tiko, two locality of the Fako Department, South-West Region of Cameroon. The method used was cluster sampling and the parcels of 100 m × 100 m were measured in each bloc prospected. Parameter such as circumference of trunk, the row number, and the number of tufts of P. capitata were noted for each parasitized tree. Clone PB 217 (61.73%) recorded the highest parasitism rate, and PR 107 the highest average of number of tufts (5±3.12 tufts/tree). Parasitized trees of the clone PR 107 were located closer to the border than the others clones, which an average depth of 16.4 m. An efficiency struggle method against P. capitata must be found to preserve the yield of rubber tree plantations.
Key words: Heveaculture, parasitism, spatial dissemination, susceptibility.
INTRODUCTION
Along with palm oil, natural rubber is one of the few agricultural products from the Third World that has resisted the collapse of world prices. As a result of globalisation, the demand for rubber is constantly increasing. Rubber is used in the manufacture of tyres, but also in everyday life (manufacture of condoms, surgical gloves and syringes). It is extracted from the latex produced by the rubber tree, which is the only exploited source of natural rubber in the world. Its exploitation is spread over three continents, namely South-East Asia, America and Africa. Asia, with 93% of world production, is the main supplier of natural rubber. Next, comes Africa (5%) and America (3%) (IRGSM, 2007). In Cameroon, rubber trees are mainly exploited in the Littoral, South-West and South regions.
Mistletoes are plants that occur in tufts and are linked to their host only by a structural and physiological bridge that constitutes an absorption or suction system, allowing the parasite to take water, minerals and additional organic matter that it needs (Bannister et al., 2002). In addition to wild trees, mistletoes attack cultivated species such as cocoa, avocado, citrus and rubber trees, reducing their yield (Sonké et al., 2000). The Loranthaceae family is widespread throughout the world and includes 77 genera and 950 species. In Cameroon, the Loranthaceae are represented by 26 species belonging to 7 genera (Polhill and Wiens, 1998).
The action of Loranthaceae is harmful in sub-Saharan Africa. Indeed, they are reported in many countries, notably in Benin, 4 genera (Agelanthus, Globimetula, Phragmanthera and Tapinanthus), in Burkina Faso, with 3 genera (Agelanthus, Globimetula and Tapinanthus) (Boussim et al., 2012; Houenon et al., 2012). In Cameroon, home gardens as well as orchards and industrial plantations containing species such as cocoa (Theobroma cacao), avocado (Persea americana), orangutan (Citrus sinensis), safflower (Dacryodes edulis), and rubber (Hevea brasiliensis) are particularly susceptible to the genus Phragmantera in the coastal region (Dibong et al., 2009; Ondoua et al., 2015; Massako et al., 2014).
It is therefore interesting to study Phragmantera capitata, the main species of the genus found in Cameroon on rubber trees, supplier of natural rubber, one of the few agricultural products of the Third World, along with palm oil, to have resisted the collapse of world prices. The aim of this study was to characterize the parasitism of P. capitata on three cultivated rubber clones.
MATERIALS AND METHODS
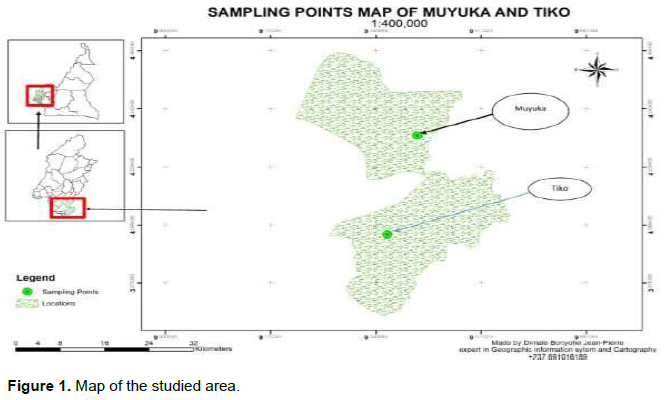
The study was conducted around of Muyuka and Tiko, two subdivisions in the Fako Department, located, respectively to the north and southeast of the city of Buea, the regional capital (Figure 1). This region is characterized by a coastal humid equatorial climate of the Cameroonian maritime domain with a long rainy season and a short dry season (December-February). Rainfall is abundant and regular, averaging about 2975 mm per year. The average temperature is 25.6°C, with March being the hottest month of the year (26.7°C) and July the least hot month (24.2°C). The monthly relative humidity remains consistently high at around 80%.
Sampling of plant groups
The work took place in four exploited plantations of the Cameroon Development Corporation (CDC), these plantations are located in the villages Big Ikange, Likomba, Meanja, Missellele, and Muyuka. The method used was cluster sampling, and the work was carried out on rubber trees between 20 and 25 years old, and the clones were selected on the basis of their representativeness in this age group. Thus, clones PB 217, PB 260, and PR 107 were selected for this study.
Delimitation of the parcels
The work took place in the period from August 2014 to September 2016. The clones were distributed in different blocks of variable dimensions. Three plots of 100 m × 100 m dimensions, so 1 ha each were delimited for each clone, for a total area of 9 ha. The trees were planted in rows, and the distance between rows was either 2.8 m or 7 m, depending on the orientation of the plot in relation to the road.
Data collection
Loranthaceae species were observed in the canopy either by naked eyes or by using a pair of binoculars. A delimber was used when the foliages of many trees were crossed, making observation difficult. Plots were prospected line by line, and the number of tufts of P. capitata was recorded for each parasitized tree. According to the number of tufts found on the trees, a scale of parasitism intensity was established with four levels: lowly parasitized (1 to 2 tufts); moderately parasitized (3 to 5 tufts); heavily parasitized (5 to 10 tufts); very heavily parasitized (> 10 tufts). Other parameters such as tree circumference and the row number were recorded. The following indices were calculated from the data collected.
The density of parasitism was calculated as the ratio of the number of parasitized trees (Np) per unit area (S).
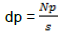
The parasitism rate was calculated as the ratio of the number of parasitized trees (Np) to the total number of trees in a block (N).
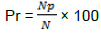
The average number of clumps is the ratio of the number of tufts of P. capitata (n) to the number of parasitized trees in a block (Np).
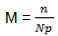
Finding P. capitata outbreaks
Outbreaks are considered as trees with large diameters and a high number of parasite clumps. In this study, foci correspond to trees of circumference class X, and which are very heavily parasitized. The class X has to do with trees which have a circumference of 90 cm and above.
Statistical analysis
Statistical analyses were performed with XLSTAT 20.6 software. Chi-square test was used to compare the parasitism rates of the clones, Kruskal-Wallis test was used to make multiple comparisons between the average number of tufts, and the means of the circumferences. Pearson correlation test was used to analyze the degree of relationship between the number of tufts of parasite, the circumference of the trees, and the position of parasitized trees in the block.
RESULTS
A total of 2111 parasitized trees were recorded for all clones, with a relative density of 235 trees/ha. Among the clones, the number of parasitized trees was 873 individuals in PB 260, 650 individuals in PB 217, and 588 individuals in PR 107, for relative densities of 219 individuals/ha, 218 individuals/ha, and 156 individuals/ha, respectively. Table 1 shows that parasitism rate in the study area was 56.35%, the highest being obtained in clone PB 217 with 61.73%. It was significantly higher than clones PR 107 and PB 217 with parasitism rates of 59.04 and 51.44%, respectively (K = 31.94; p < 0.0001). The number of P. capitata tufts encountered was 7002 in the study area, for an average of 4 tufts/tree. Among clones, the highest number of tufts was found in PR 107 with 2689 tufts, followed by 2293 tufts in PB 217, and 2020 clumps in PB 260. The average number of P. capitata tufts among clones was significantly higher in PR 107, with 5 ± 3.12 tufts/tree (K = 458.94; p < 0.0001). 4 ± 2.9 tufts/tree were found in PB 217 and 3 ± 1.57 tufts/tree in PB 260. These results show that the horizontal spread of P. capitata is higher in clone PB 217, and its vertical spread is higher in PR 107.
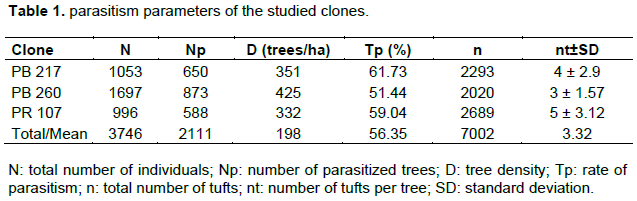
The study of parasitism intensity showed that 64.03% of individuals in clone PB 260 are weakly parasitized, and only 0.23% of individuals in the same clone are very heavily parasitized. In PB 217 and PR 107, the rate of very heavily parasitized trees was higher than the average value, these rates being 4.31 and 7.31%, respectively. Clone PR 107 is the only one of the three clones whose number of weakly parasitized trees is lower than the number of moderately and strongly parasitized trees (Figure 2).
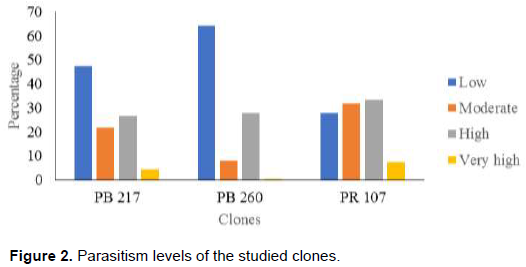
Figure 3 shows the average circumferences of parasitized trees in the different clones. It appears that the circumferences of clone PB 217 with 101.57±23.91 cm, was higher than those of PR 107 and PB 260 with 96.77±16.65 cm in clone PR 107, and 93.18±14.39 cm in PB 260 (K = 35.22; p < 0.0001). The study of the classes of circumference showed that class X was the most represented in the clones PB 217 and PR 107 with the proportions of 39.54 and 38.16%, respectively, and the class VIII was the highest represented in the clone PB 210 (Figure 4).
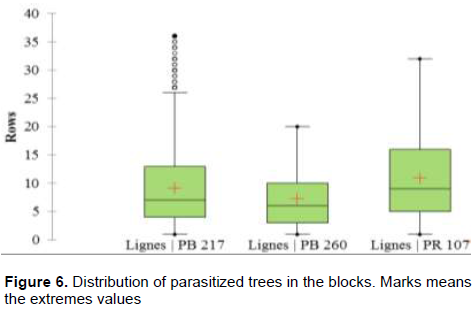
Figure 6 show that the mean position of parasitized trees was 19.6 m to the border, the median was 16.8 m, and the 3rd quartile was 28 m. Parasitized trees of the clone PB 260 were the closest to the border of all of the studied clones. The average position of them was on the row 9, that is, at 25.2 m to the border. Median was found in row 6, and the third quartile was in the row 9. For the clone PB 217, parasitized trees have an average position on the row 9, the median was found in the 7th row, and the 3rd quartile was in the 13th row. For PR 107, the average position was on the 10th row, the median was at row 9, and the 3rd quartile was at row 16.
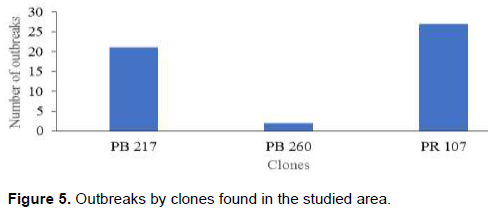
Level of parasitism of the class 10 of each clone was permitted to determine 51 outbreaks of P. capitata. The clone PR 107 is the main focus of P. capitata spread in the study area with 27 outbreaks, followed by clone PB 217 with 22 outbreaks. Clone PB 260 contains only 2 outbreaks (Figure 5).
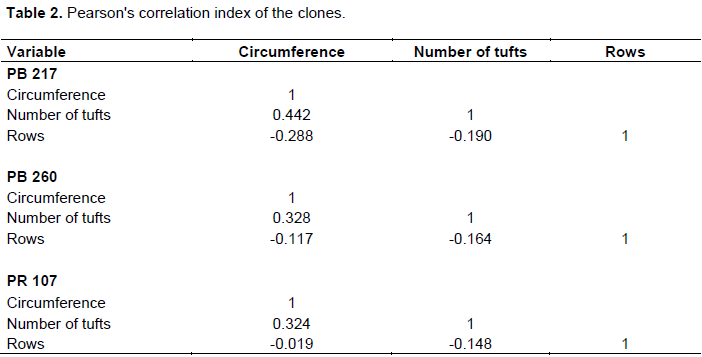
The study of the correlation between morphometric parameters of the clones shows that circumference and number of tufts correlated positively, but have correlated negatively with depth in all of the studied clones (Table 2).
DISCUSSION
The results showed that all the clones studied were parasitized. This shows that there are no clones resistant to Loranthaceae parasitism. This was also demonstrated in Nigeria where Loranthaceae attacked both native and exotic clones (Orumwense et al., 2012). Clone PB 260 showed the lowest rate of parasitism among the 3 clones studied. Similar results were found in Gabon, where clones AVROS 2037, PB 217, PB 235, and GT 1 were characterised as highly parasitised, and clones PB 261, PB 260 and RRIM 600 were characterised as low parasitism (Engone, 2006). This could be explained by the fact that clone PB 260 has dense foliage, allowing little light to pass through, thus making it difficult for P. capitata, which is known to be a heliophilic species, to establish itself.
The gradient of tree susceptibility to parasitism was explained by the presence of anatomical parameters (thickness of the suber and collenchyma) and biochemical parameters (phenolic compounds) in poplar. These characteristics, which allow plants to defend themselves against external aggressions, were found to be strongly correlated with resistant clones, which were characterised by a high thickness of suber and cortical parenchyma, as well as a higher density of tannins (Hariri et al., 1992). However, in African Loranthaceae, phenolic and tannin contents did not show significant differences (Engone, 2006). On the other hand, low tree density in a plot increases the amount of incident light, thus favouring the establishment of P. capitata. This observation was confirmed in clone PR 107, which had a lower tree density and a higher rate of parasitism. The study of the row parasitized trees showed that clone PB 260 are located more closest to the edge of the plantations, and those of clone PR 107 more inside the plantations. It means that light is a factor greatly in parasitism in clone PB 260, but would have less effect in PR 107.
The high rate of parasitism in all the clones studied reflects the high implementation of P. capitata in the region studied. Similar results were found in rubber plantations in southern Cameroon (63.4%), in citrus plantations in Benin (59.87%), or on tree legumes in Côte d'Ivoire (56%) (Dibong et al., 2009; Houenon et al., 2012; Soro, 2005). However, this infestation rate is much lower than the 96% observed on shea in northern Côte d'Ivoire (Traoré et al., 2003), and much higher than the 10.4% found in rubber plantations in southern Côte d'Ivoire (Soro et al., 2011). The difference observed with shea is explained by the fact that the shea plantations were located in a semi-arid zone. In this type of environment, the trees are spaced out and strongly branched, and would thus constitute favourable sites for the installation of Loranthaceae. In Côte d'Ivoire, where the rate of parasitism was 10.4%, the explanation could be found in the fact that the studies were carried out on trees aged 15 to 20 years, whereas the trees in this study belonged to the 20 to 25 years age group, thus confirming that parasitism increases with the age of the host plant (Engone, 2002).
The difference between the parasitism rates of rubber trees in Tiko and Nyete can be explained by the fact that parasitism depends on geographical location (Raynal-Roques and Paré, 1998). On the other hand, the Muyuka and Tiko areas are characterised by a Cameroonian type climate with one dry season, while the Nyete area has a climate with two dry seasons, thus showing that P. capitata develops optimally in areas with high rainfall.
CONCLUSION
P. capitata shows a high rate of parasitism in all clones. However, genotypic and phenotypic variability allows us to characterise parasitism at the level of each clone. Indeed, P. capitata is localised at the edge of the plantation in PB 260, whereas it extends much further inland in clones PB 217 and PR 107. Since the main foci of propagation are the individuals of clones PR 107 and PB 217, it is important to set up a monitoring system to follow the evolution of the spread of this species in rubber plantations, in order to avoid a drastic drop in production.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
The authors thank the Cameroon Development Corporation (CDC) for giving their experimental parcels for this study.
REFERENCES
|
Bannister P, Graham L, Strong, Inge A (2002). Differential accumulation of nutrient elements in some new mistletoes and their hosts. Functional Plant Biology 29(11):1309-1318. |
|
|
Boussim IJ, Yonli D, Medah NM, Guinko S, Sallé G (2012). Prospects for an integrated control of Loranthaceae species parasitizing Vitellaria paradoxa C. F. Gaerth in Burkina Faso. International Journal of Biological and Chemical Sciences 6(1):355-364. |
|
|
Dibong SD, Engone Obiang NL, Din N, Priso RJ, Taffouo VD, Fankem H, Amougou A (2009). Niveau d'infestation des arbres fruitiers des groupements végétaux par Phragmanthera capitata (Sprengel) S. Balle (Loranthaceae) dans la région littorale du Cameroun. International Journal of Biological and Chemical Sciences 3(2):347-454. |
|
|
Engone ONL (2002). Les Loranthaceae au Gabon. Master dissertation, Université Pierre et Marie Curie. Paris, France. |
|
|
Engone ONL (2006). Biologie et écologie des Loranthaceae parasites du Gabon. Impact de Phragmanthera capitata sur la productivité des hévéas. Ph.D thesis, University Pierre et Marie Cury, Paris, France. 189 p. |
|
|
Houenon JG, Yedomonhan H, Adomo AC, Tossou MG, Omorou M, Akoegninou A (2012). Les Loranthaceae des zones guinéenne et soudanoguinéenne au Bénin et leurs hôtes. International Journal of Biological and Chemical Sciences 6(4):1669-1686. |
|
|
Massako F, Mony R, Tchata M, Dibong SD (2014). Inventaire et évaluation de l'infestation des Loranthaceae sur les essences du safoutier (Dacryodes edulis (G. Don) H. J. Lam au Nord-Est de Douala, Cameroun. Sciences, Technologies et Développement 15:87-92. |
|
|
Ondoua JM, Dibong SD, Taffouo VD, Ngotta Biyon JB (2015). Parasitisme des champs semenciers de cacaoyers par les Loranthaceae dans la localité de Nkoemvone (sud Cameroun). Journal of Applied Biosciences 85:7794-7803. |
|
|
Polhill R, Wiens D (1998). Mistletoes of Africa. Royal Botanic Gardens, Kew. 370 p. |
|
|
Raynal-Roques A, Paré J (1998). Biodiversité des Phanérogames parasites: leur place dans la classification systématique. Adansonia 20(2):313-322. |
|
|
Sonké B, Kenfack D, Tindo M (2000). Parasitisme de l'avocatier (Persea americana, Lauraceae) par les Loranthacées dans la région de Yaoundé (Cameroun). Fruits 55(5):325-331. |
|
|
Soro K (2005). Les Loranthaceae des Légumineuses arborées introduites dans la zone forestière d'Oumé : espèces rencontrées, spectre d'hôtes et taux d'infestie. Master dissertation, University of Cocody, Abidjan, Côte d'Ivoire. 51 p. |
|
|
Soro K, Soro D, N'Guéssan K, Da KP, Traoré D (2011). Parasitisme des Loranthaceae sur les cacaoyers et les caféiers en zone forestière : cas de la Région d'Oumé en Côte d'Ivoire. Annales Botaniques d'Afrique de l'Ouest 7:1-6. |
|
|
Traoré D, Da KP, Soro D (2003). Lutte contre les plantes vasculaires parasites du karité, dans le Nord de la Côte d'Ivoire : Cas du parc naturel à karité de Tengrela. PEPAISA Report. Abidjan : University of Cocody. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


