Full Length Research Paper

ABSTRACT
The aim of this study was to evaluate the agronomics, physiological and biochemical characteristics of different peanut seeds and plant genotypes. The procedures have been performed during the secondary crop in the agricultural year 2013 to 2014, after the peanut harvest in Barbalha, Ceará – Brazil. The experiment was laid out in split-plot in randomized complete block design with four replicates, constituting scheme 4×5 factorial analysis, providing four levels of replacement of ET0 (40, 70, 100 and 130%) and five genotypes of peanut (BRS Perola Branca, L7 Beje, Runner, BRS Havana, BR 1), which were measured: number of leaves, total phytomass, and total chlorophyll content was performed, as well as hundred-seeds weight, pods weight per plant, number of pods per plant and seed yield per plant oil yield and fatty acid composition. It is shown herein that the interaction between peanut genotypes and water restriction levels promote a decrease in plant growth, physiological factors, and biochemical production of oil, possibly due to a decrease in the photosynthetic metabolism, whereas the plants exposed to water stress conditions modulated all ecophysiological responses in favor of a better performance in the harsh environment. The significant differences exhibited by different cultivars in this study for oil content, some productive characteristics and fatty acids compositions could be attributed to the genetic make-up of a particular cultivar (BR 1), its place of the environmental to reach high oil quality. Knowing better combination effects of climatic fluctuations and botanical types on fatty acid composition would be useful in designing management practices to obtain a specific oil quality and improving predictions of crop models in northeastern Brazil.
Key words: Arachis hypogaea L., water deficiency, adaptability, tolerance.
INTRODUCTION
Peanut (Arachis hypogaea L.) is grown in many arid and semi-arid regions of Brazil and, therefore, needs to be irrigated for economical yield. However, the vegetative preflowering growth stage and the late stage of pod maturation have been shown to be non responsive to water stress (Gohari and Bazkiyaei, 2012).
Peanut is tolerant to water stress conditions but drought conditions have adverse effects on the pod yield and seed quality (Nageswara Rao et al., 1989; Sulc and Franzluebbers, 2014). The effect of drought on the chemical composition of the Peanut seeds has been reported to be limited in the mid-season drought but significant in end-season drought (Sun et al., 2014).
Furthermore, increased worldwide demand for water due to rapid population growth and non sustainable irrigation practices have resulted aquifer depreciation, limiting the availability of water for irrigation. To meet future food supply demands, crop production has to be increased, but it must do so under the constraints of less water and, most likely, less farm land. EMBRAPA - National Center for Research on Cotton, Campina Grande-PB, Brazil, and The State University of Paraíba are working in cooperators to help peanut farmers to maintain and improve production in a water-limited changing environment.
Drought-stressed plants lose moisture from pods which lead to the reduction in the seeds physiological activity, thereby increasing the susceptibility to fungal invasion. Besides affecting food quality, drought stress is also known to alter nutritional quality of peanut seed proteins (Holbrook et al., 2000).
Peanut is cultivated in different regions of the world, under diverse climate conditions and in Brazil it grows primarily under rain fed conditions in the northeastern region (Santos et al., 2006). EMBRAPA - National Center for Research on Cotton has been conducting research on peanut cultivars BR 1, BRS 151 L-7 and BRS Havana, among others (Santos et al., 2006). Faced with scarcity of water resources, drought is the single most critical threat to world food security. It was the catalyst of great famines in the past. Because the world’s water supply is limiting, future food demand for rapidly increasing population pressures is likely to further aggravate the effects of drought (Somerville and Briscoe, 2001). The severity of drought is unpredictable as it depends on many factors such as occurrence and distribution of rainfall, evaporative demands and moisture storing capacity of soils (Wery et al., 1994).
Reddy et al. (2003) observed the application of the different irrigation rates amounts has induced different responses on the protein content of the seeds; the plants with adequate water irrigation not only give more kernels, but also higher levels of total proteins and oil content. The major fatty acid components of peanut oil are oleic, linoleic, palmitic and stearic acids, but others are found in different proportions: 0.13 to 0.33% myristic, 8.70 to 13.03% palmitic, 0.23 to 0.47% palmitoleic, 3.77 to 4.53% stearic, 43.13 to 55.10% oleic, 25.13 to 35.20% linoleic, 0.20 to 0.30% linolenic, 1.53 to 1.93% arachidic, 0.40 to 1.37% gadoleic and 2.40 to 3.47% behenic acids (Özcan and Seven, 2003).
The objective of the present study was to determine the effect of differentiated irrigation on the agronomic performance, productivity and fatty acid composition of peanut oils of several genotypes subjected under drought stress conditions.
MATERIALS AND METHODS
Field experimentation
This experiment was conducted in Barbalha-CE where is the Northeast area of Brazil with an altitude of 414 m above sea level and average temperature of 25.2°C. Annual precipitation varies from 1001.4 to 1054.1 mm distributed into two distinct rain seasons. Weather data were obtained from the meteorological office in Barbalha-CE. Five genotypes of peanut (BRS Perola Branca, L7 Beje, Runner, BRS Havana, BR 1) were grown under four irrigation levels L1, L2, L3 and L4. The value of accumulated total evaporation four different levels (L1=40%, L2=70%, L3=100%, and L4=130%) were measured using Class A Pan evaporation containers during 130 days after plant emergence. The experiment was conducted in the trial area of EMBRAPA - National Center for Research on Cotton. The experiment was laid out in split-plot in randomized complete block design with four replications on the entisol soil with an initial characterization of soil fertility are presented for the respective soil depths of 0.00 to 0.20 and 0.20 to 0.40 m as follows: OM (12.3; 8 mg dm-3), pH (7.65; 7.8), lime content of 8.67%, P (8; 3 mg dm-3); K (0.2; 0.1 mmolc dm-3); Ca (1; 0 mmolc dm-3), Mg (1; 0 mmolc dm-3), H+Al (28; 23 mmolc dm-3), Al (0; 6 mmolc dm-3), BS (19; 9 mmolc dm-3), CEC (47; 32 mmolc dm-3), V (40; 28%), and m (32; 42%). In initial characterization of physical properties, we verify that soil resistance to penetration at the depths of 0.00 to 0.10, 0.10 to 0.20, 0.20 to 0.30, and 0.30 to 0.40 m were 1.054, 1.445, 1.558, and 1.513 MPa and soil moisture at those depths were 0.035, 0.030, 0.030, and 0.040 kg kg-1, respectively. Sowing was performed on irrigated seedbeds. Plot area was 2.8 × 7.0 m. The seeds were sown by using sewing machine at a spacing of 0.38 m distance between seeds and 0.70 m distance between the rows (Figure 1).

Variables measured
At the 130th days after sowing (DAS), plants were collected to determine the initial growth characteristics: number of leaves, total phytomass (g plant-1) and total chlorophyll content (mg g-1 FW). The harvest was performed taking five random plants from the center line of each treatment, in each block. The plants were separated by treatment and placed in screened greenhouse for three days in order to dry the pods. The productive components evaluated in this study was 100-seeds weight (g), pods weight per plant (g), number of pods per plant, seed yield per plant (g), oil content and fatty acid composition.
Chlorophyll determination
Extraction and determination of chlorophyll were performed according to the method of Arnon (1949). Five hundred milligrams of fresh leaf material was ground with 10 ml of 80% acetone at 4°C and centrifuged at 2500 rpm for 10 min at 4°C. This procedure was repeated until the residue became colorless. The extract was transferred to a graduated tube and made up to 10 ml with 80% acetone and assayed immediately. Three milliliters aliquots of the extract were transferred to a cuvette and the absorbance was read at 645 nm with a spectrophotometer (UV-1800 UV-VIS - SHIMADZU) using 80% acetone as blank. Chlorophyll content was expressed in mg g-1 fresh weight (FW) using the formula: (mg ml-1) = (0.0202) × (A.645) + (0.00802) × (A.663).
Oil content
Oil content was determined using a commercial nuclear magnetic resonance spectrometer as described by Jambunathan et al. (1985). All readings were taken on oven-dried (110°C, 16 h) samples and the values were expressed on a uniform 5% seed moisture content basis.
1H NMR Spectra
1H NMR spectra were recorded on a Bruker AMX500 spectrometer operating at 500 MHz for the proton nucleus at room temperature. The phosphitylated oil samples used in 31P NMR experiments were used to obtain 1H NMR spectra with the following acquisition parameters: time domain, 32K; 90° pulse width, 9.3 µs; spectral width, 12 ppm; relaxation delay, 2 s. Sixteen scans and four dummy scans were accumulated for each free induction decay. Baseline correction was performed carefully by applying a polynomial fourth order function in order to achieve a quantitative evaluation of all signals of interest. The spectra were acquired without spinning the NMR tube in order to avoid artificial signals, such as spinning sidebands of the first or higher order.
Preparation of fatty acid methyl esters (FAMEs) and gas chromatography
Seed samples were taken for total fatty acid analyses. Total fatty acid content was analyzed by using a method modified by Wu et al. (1994). In this method seed samples were soaked in 2 ml of 2% sulphuric acid in dry methanol for 16 h at room temperature, followed by 80 min of heating at 90 °C to convert the fatty acids into methyl derivatives (FAMEs). Methyl-heptadecanoate (17:0-ME) was added as an internal standard. The FAMEs were extracted in 2 ml water and 3 ml hexane and then determined by gas liquid chromotography (GLC). The fatty acid methyl ester composition was analyzed by using a Varian 3400 gas chromotography equipped with a Supelcovax-10 fused slica capillary column (30 m × 0.25 μm film thickness). The column’s initial temperature was kept at 160°C for 15 min so that in this temperature an increase could be occurred at the rate of 5°C min-1. The temperatures of the injector and the detector (FID) were 240 and 280°C, respectively. The carrier gas was nitrogen with a flow rate of 1 to 2 ml min-1. Split ratio was adjusted to 30 ml min-1. The injected volume of the sample was 1 μl. Fatty acids were identified by retention time relative to that of an internal standard. FAMEs were identified by comparing the retention times with those of the standards. Fatty acid content was computed as weight percentage of the total fatty acids by using the GC area counts for various FAMEs.
Statistical analysis
Statistical evaluation was carried out by using Sigma Plot 11.0. An analysis of variance was performed on the data, and means were separated using Tukey’s multiple comparison procedure (probability of 5%).
RESULTS AND DISCUSSION
According to the analysis of variance (data not shown), the irrigation levels × genotypes interaction effects were significant (P<0.01) for all characters. The effect of irrigation levels and genotypes was also significant (P<0.01) for all characters.
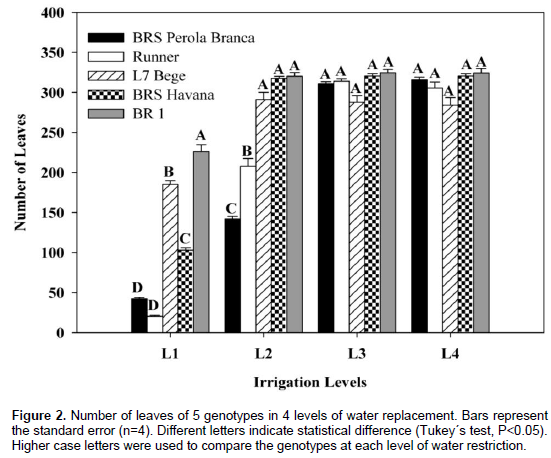
The number of leaves of all peanut genotypes significantly was decreased with increasing water deficit levels (Figure 2).
BR 1 showed significantly higher number of leaves values than the other genotypes at different levels of water. BR 1, BRS Havana and L7 Bege showed significantly higher leaves number than BRS Perola Branca and Runner in irrigation level L2, except for the L1 treatment. Non-significant differences were observed in number of leaves of L7 Bege, BRS Havana and BR 1 for all irrigation treatments (L2, L3 and L4). In water treatment L1, genotype BR 1 showed higher leaves number compared to the other genotypes. Water deficit induces several physiological and biochemical changes in plants depending on intensity and duration of stress (Pattangual and Madore, 1999). General effect of water stress in plants is manifested in terms of osmotic stress which results in lower leaf water potential, turgor potential and stomatal conductance (Dzotsi et al., 2015). This study demonstrated that water stress led to differential responses in peanut genotypes. The reduction in the number of leaves is presumably a response to the decrease in leaf water content and changes in membrane stability under water stress (Chakraborty et al., 2015).
Plant weight of all genotypes was decreased significantly by applying different water stress levels as compared to control treatment (Figure 3). At L1, BR 1 showed significantly plant weight than the other peanut genotypes.
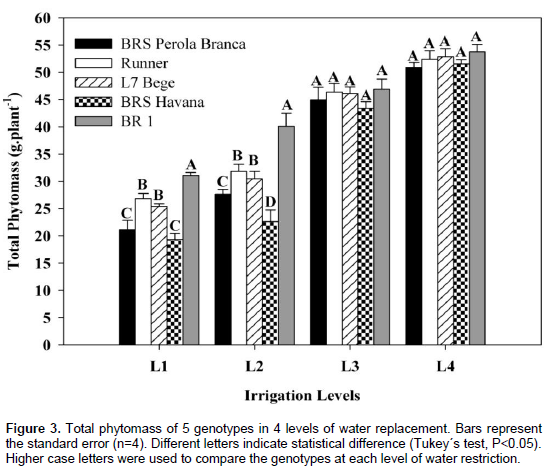
At L1 and L2, BRS Perola Branca, Runner, L7 Bege and BRS Havana showed low plant weight, whereas BR 1 showed significantly highest plant weight. It is reported that a positive association is observed between total phytomass accumulation and pod yield under moisture stress conditions (Reddy et al., 2003). Total phytomass accumulation of peanut was reduced due to stress (Richards et al., 2014). Sulc and Franzluebbers (2014) reported a decrease in total phytomass production due to drought in peanut. Reddy et al. (2003) reported that there was a close relationship between total phytomass production and transpiration with an average production of 3.0 mg total phytomass g-1 water. They also noted that the amount of total phytomass accumulated by a crop was closely related to the water transpired. Reddy et al. (2003) reported that there was no recovery of total phytomass in peanut when stress was imposed at of the beginning of emergence, while recovery of total phytomass was found when stress was imposed from emergence to the begining of flowering and from emergence to start of pegging.
The total chlorophyll content of all peanut genotypes was decreased significantly with increasing water deficit levels as compared to control treatment (Figure 4). At L1, BR 1 showed significantly higher total chlorophyll content than the other genotypes.
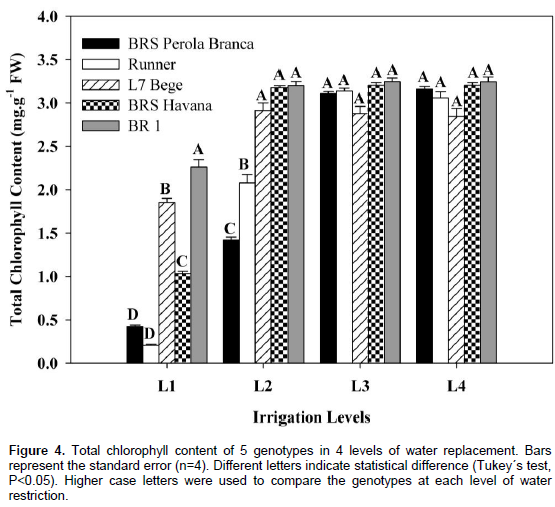
At L1 and L2, phenotype L7 Bege showed significantly higher chlorophyll content compared to the other genotypes, except for BRS Havana and BR 1 in L2. Non-significant differences were observed in total chlorophyll content of BRS Perola Branca and Runner. In the present study, the total chlorophyll content of the leaves was decreased in most peanut cultivars when exposed to drought stress. Water deficit degrades chlorophyll and impairs its biosynthesis as well (Reddy et al., 2003), which was reported by other researchers in many crops, including peanut (Chakraborty et al., 2015). The ability to maintain a stable content of chlorophyll under water deficit conditions is a good measure of the peanut genotypes capability to cope with drought stress during initial stages of growth (Arunyanark et al., 2008). Here, the tolerant cultivar BR 1 showed higher adaptation to maintain stable contents of chlorophyll at final growth stage indicating more tolerance to water stress conditions. The 100-seeds weight, pods weight per plant, number of pods per plant and seed yield per plant of all genotypes significantly decreased with increasing water deficit levels (Figure 5A, B, C and D). At L1, highest 100-seeds weight, pods weight per plant, number of pods per plant and seed yield per plant were observed for BR 1, being significantly higher than the other peanut genotypes.
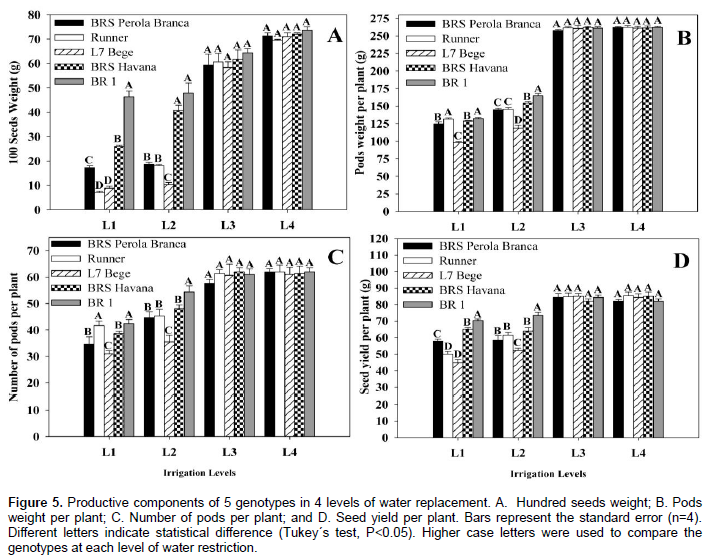
Non-significant differences were observed in 100-seeds weight of genotypes in L3 and L4. Whereas, BRS Perola Branca, L7 Bege and Runner showed consistently the lowest 100-seeds weight, pods weight per plant, number of pods per plant and seed yield per plant values. At L2 and L3, more pronounced differences in the seeds weight, pods weight per plant, number of pods per plant and seed yield per plant were observed compared to L1 and L2. BR 1 showed significantly higher 100-seeds weight, pods weight per plant, number of pods per plant and seed yield per plant than the other genotypes.
For the variable weight of a hundred seeds (100SW, shown in Figure 5A), the statistical analysis that best significant. From this result, it was calculated that 100SW reached a maximum (77.50 g) at the irrigation level of L3 and L4. Corroborating with this study, Araújo and Ferreira (1997) showed a 100SW of 75.25 g for the peanut crop grown without water stress. On the other hand, Silva et al. (1998) obtained opposite results for this study. According to the authors, the irrigation depths applied measuring (300, 500 to 700 mm) by furrow irrigation, did not significantly influence this variable in the peanut crop. Du et al. (2015) reported that the pods weight of peanut variety TMV-2 was decreased under water stress conditions. Arunachalam and Kannan (2013) reported that seed weight was deceased significantly due to moisture stress during pod development stage in peanut. Junjittakarn et al. (2014) reported that moisture stress at flowering stage in peanut increased 100-seeds weight, while at pegging reduced weight compared to control. Water deficit during kernel or seed development stages reduced the weight of kernel. Srinivasan et al. (1987) reported that pod weight of peanut decreased due to water stress. A reduction of 22% in 100-seed weight was observed during flowering stage compared with control under drought stress conditions (Pathak et al., 1988). Nautiyal et al. (1991) reported that peanut cultivars exposed to soil moisture stress at different growth stages especially during early vegetative phase resulted in an increase in seed weight. Hundred-seeds weight was reduced greatly due to the moisture stress occurred at pod development stage (Reddy et al., 2003). Pathak et al. (1988) reported that plants subjected to drought stress during flowering stage resulted in reduction (29%) in 100-pods weight compared with control. Arunachalam and Kannan (2013) stated that there was decrease in 100-kernels weight and 100-pods weight under moisture stress conditions in simulated drought treatments compared to adequately irrigated control.
It can be seen in Figure 5B there was a significant response of pods weight per plant with increased irrigation levels. The model estimates an optimum pods weight per plant of 271.86 g per plant for the irrigation level of L3 and L4, in other words, the pods weight per plant would increase from the treatment in 44.64% for an irrigation depth L3 and L4. In pepper crop, Azevedo et al. (2005), testing different irrigation levels, although based on water evaporation in Class “A” tank, they observed that increasing irrigation levels influence significantly the average pod weight.
The statistical analysis showed that the number of pods per plant, relatively to the different irrigation levels, between genotypes, except for L3 and L4 (Figure 5C). The increasing of irrigation amount resulted in a maximum number of pods per plant equal to 65, obtained with irrigation level of L3 and L4. Távora and Melo (2001) working with the peanut crop in greenhouse conditions, observed similar results to this study for this variable. It should be noted that the occurrence of a water deficit in a peanut crop, during the growth and development phases of the gynophores and pods, causes a decrease in the number of pods (Távora and Melo, 2001).
The productivity of grain in the peanut crop was influenced by the increase in the amount of water applied (Figure 5D). This data estimates a maximum grain yield value of 88.97 g per plant which corresponds to an irrigation level of L3 and L4. The yield obtained with this irrigation level is above the average grain yield values of 1,151.6 kg ha-1 in Ceará (IPCE, 2010) and below the average productivity values of 2,225 kg ha-1 in Brazil (IBGE, 2011). Under field conditions, the irrigation depth of 700 mm provided a peanut yield of 2,026 kg ha-1 (Silva et al., 1998). On the other hand, Silva and Beltrão (2000) found a grain yield of 1,671 kg ha-1 with the peanut crop being irrigated with a 500 mm of irrigation depth.
The oil content of all peanut genotypes was decreased significantly with increasing water deficit levels as compared to the control treatment (Figure 6). At L1, BR 1 maintained significantly higher oil content than the other peanut genotypes in L1 and L2, with the exception of BRS Havana in L2.
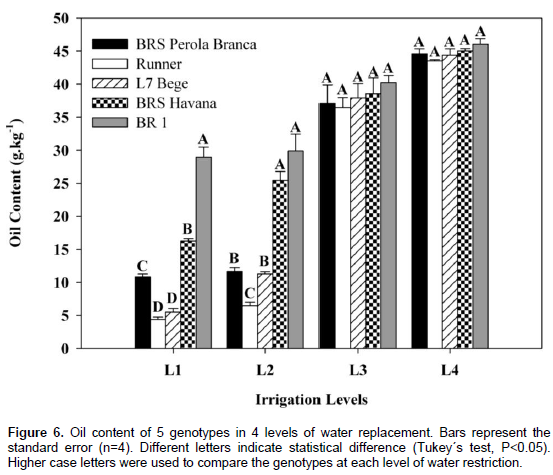
Phenotype Runner significantly maintained lowest oil content compared to the other genotypes under all irrigation treatments. At L2 and L3, genotypes were ordered in terms of oil content as: BR 1 > BRS Havana > L7 Bege > Runner. The chemical composition of peanut seed is influenced, among other factors, by its stage of maturity (Akhtar et al., 2014). The present study therefore included only sound mature seeds to eliminate maturity-related differences in seed chemical composition. Total oil was affected by water deficit levels. This decrease was progressive, corresponding to the intensity of water deficit. Differences in oil content became significant only under moderate to intense water deficit. Our results, however, are in agreement with previous reports that late-season drought had great effect on total oil content (Sun et al., 2014), or that mid-season drought reduced total oil content (Bhalani and Parameswaran, 1992). This could be due to the differences in genotypes and in the timing and intensity of drought in these studies. Further, growing conditions and crop management practices (foliar diseases control) also influence total oil, total protein and fatty acids in peanut (Akhtar et al., 2014). In the present study, genotypes interacted with water deficit levels for total oil.
Palmitic, stearic, oleic, linoleic, arachidic and behenic acids are the principal fatty acids that these constitute whole peanut seed fatty acid composition's of 98%. The interaction effect of irrigation levels x genotypes was found highly significant (P<0.01) for all tested fatty acids (Figure 7). The fatty acid composition of all peanut genotypes changed significantly with increasing water deficit levels as compared to the control treatment. Non-significant differences were observed in fatty acid composition of genotypes at L3 and L4.
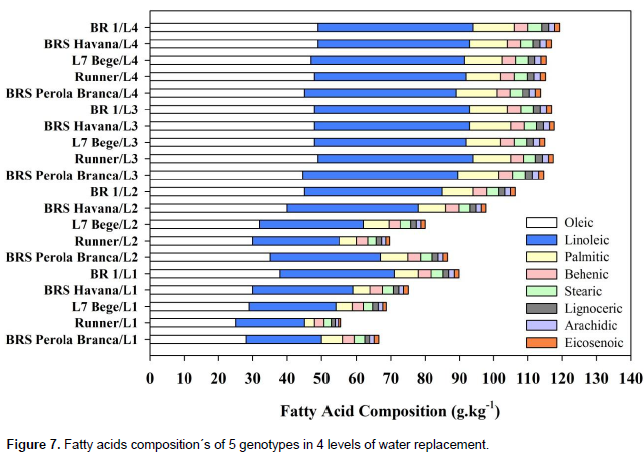
In L1 and L2 the highest Oleic acid content was obtained from BR 1 and the lowest was obtained from Runner, the highest Linoleic acid was obtained from BR 1 and BRS Havana, while the lowest was obtained from Runner. The highest Palmitic acid content was obtained from BR 1, while the lowest was obtained from L7 Bege and Runner, the highest Behenic acid content was obtained from BR 1, while the lowest was obtained from Runner. The highest Stearic acid content was obtained from BR 1 and the lowest was obtained from Runner interaction. The highest Lignoceric, arachidic and Eicosenoic acids content were obtained from BR 1 and the lowest were obtained from Runner, respectively. Fatty acids are synthesized by consecutive desaturation from stearic (C18:0) to oleic (C18:1) to linoleic (C18:2) fatty acid (Akhtar et al., 2014). In the present study, water deficit levels had significant effect on the major fatty acids. Stearic and oleic acids decreased, just like linoleic and behenic acids decreased under water deficit levels. The decrease was progressive with increasing water deficit. Differences in oleic, stearic, linoleic and behenic acids became significant at a low level of moisture deficit. These results differ from those of Hashim et al. (1993), who reported an increase in palmitic and linoleic acids and a decrease in stearic, oleic and eicosenoic acids when Runner was exposed to drought stressed for 30 days at maturation. When water stress was occurred at the pre-flowering and pod formation periods, an increase in behenic and lignoceric acids was observed. These differences between the two studies could be ascribed to the larger number of genotypes in the present study.
CONCLUSIONS
Water deficit levels affect the chemical composition of peanut seed in a significant manner, and total oil and fatty acids decreased in seeds, due to water deficit levels. This decrease was progressive and associated with increasing degree of water deficit. The plants irrigated from 100% and 130% had better response. Among all genotypes analyzed, cultivar BR 1 showed better agronomic performance, being more adapted to water deficiency, therefore showing a better economic viability, and would be the ideal for family farming in semi-arid regions of northeastern Brazil.
CONFLICT OF INTEREST
The authors have not declared any conflict of interest.
ACKNOWLEDGEMENTS
We are grateful to Dr. Napoleão Esberard de Macêdo Beltrão, in memoriam, by contribution to the Graduate Program in Agricultural Sciences of UEPB. The first author thanks CAPES (Higher Education Coordination Agency, linked to the Ministry of Education, Brazil) for the fellowship. This work was supported by the project EMBRAPA Cotton.
REFERENCES
| Araújo WF, Ferreira LGR (1997) Efeito do déficit hídrico durante diferentes estádios do amendoim. Pesq. Agropec. Bras. 32(5):481-484. | ||||
|
Akhtar S, Khalid N, Ahmed I, Shahzad A, Suleria HAR (2014). Physicochemical characteristics, functional properties, and nutritional benefits of peanut oil: a review. Crit. Rev. Food Sci. Nutr. 54(12):1562-1575. Crossref |
||||
|
Arnon DI (1949). Copper enzymes in isolated chloroplasts: polyphenoloxidase in Beta vulgaris. Plant Physiol. Maryland 24(1):1-15. Crossref |
||||
|
Arunachalam P, Kannan P (2013). Screening for Drought Tolerant Groundnut (Arachis hypogaea L.) Lines Suitable for Rainfed Alfisol. Asian J. Agric. Res. 7:35-42. Crossref |
||||
|
Arunyanark A, Jogloy S, Akkasaeng C, Vorasoot N, Kesmala T, Rao RCN, Wright GC, Patanothai A (2008). Chlorophyll stability is an indicator of drought tolerance in peanut. J. Agron. Crop. Sci. 194:113-125. Crossref |
||||
| Azevedo BM, Chaves SWP, Medeiros JF, Aquino BF, Bezerra FML, Viana TVA (2005). Rendimento da pimenteira em função de lâminas de irrigação. Rev. Ciênc. Agron. 36(3):268-273. | ||||
| Bhalani GK, Parameswaran M (1992). Influence of differential irrigation on kernel lipid profile in groundnut. Plant Physiol. Biochem. New Delhi. 19:11-14. | ||||
|
Chakraborty K, Singh A, Kalariya K, Goswami N, Zala P (2015). Physiological responses of peanut (Arachis hypogaea L.) cultivars to water deficit stress: status of oxidative stress and antioxidant enzyme activities. Acta Bot. Croat. 74(1):123-142. Crossref |
||||
|
Du T, Kang S, Zhang J, Davies WJ (2015). Deficit irrigation and sustainable water-resource strategies in agriculture for China's food security. J. Exp. Bot. 66(8):2253-2269. Crossref |
||||
|
Dzotsi KA, Basso B, Jones JW (2015) Parameter and uncertainty estimation for maize, peanut and cotton using the SALUS crop model. Agric. Sys. 135:31-47. Crossref |
||||
| Gohari A, Bazkiyaei ZB (2012). The optimum amount of water use and nitrogen fertilizer in the peanut (Arachis hypogaea L.) with the aim of achieving net profit. Int. J. Agric. Crop Sci. 9:518-524. | ||||
|
Hashim IB, Koehler PE, Eitenmiller RR, Kvien CK (1993). Fatty acid composition and tocopherol content of drought stressed florunner peanuts. Peanut Sci. 20:21-24. Crossref |
||||
|
Holbrook CC, Kvien CK, Rucker KS, Wilson DM, Hook JE, Matheron ME (2000). Preharvest aflatoxin contamination in drought-tolerant and drought-intolerant peanut genotypes, Peanut Sci. 27:45-48. Crossref |
||||
| IBGE - Instituto Brasileiro de Geografia e Estatística (2011). Produção Agrícola. | ||||
|
Jambunathan R, Raju SM, Barde SP (1985). Analysis of oil content of groundnuts by nuclear magnetic resonance spectrometry. J. Sci. Food Agric. 36:162-166. Crossref |
||||
|
Junjittakarn J, Girdthai T, Jogloy S, Vorasoot N, Patanothai A (2014). Response of root characteristics and yield in peanut under terminal drought condition. Chil. J. Agric. Res. 74(3):249-256. Crossref |
||||
|
Nageswara Rao RC, Williams JH, Singh M (1989). Genotypic sensitivity to drought and yield potential of peanut. Agron. J. 81:887-893. Crossref |
||||
| Nautiyal PC, Ravindra S, Joshi YC (1991). Moisture stress and subsequent seed viability physiological and biochemical basis for viability differences in Spanish groundnut in response to soil moisture stress. (En) oleagineux. 46:153-158. | ||||
| Özcan M, Seven S (2003). Physical and chemical analysis and fatty acid composition of peanut. peanut oil and peanut butter from ÇOM and NC-7 cultivars. Grasas Aceites. 54:12-18. | ||||
| Pathak SR, Patel MS, Qureshi AU, Ghodasara GV (1988). Effect of water stress on yield and diurnal changes of biophysical parameters of groundnut. Legume Res. 11:193-195. | ||||
|
Pattangual W, Madore M (1999). Water deficit effects on raffinose family oligosaccharide metabolism in Coleus. Plant Physiol. 121:993-998. Crossref |
||||
|
Reddy TY, Reddy VR, Anbumozhi V (2003). Physiological responses of groundnut (Arachis hypogea L.) to drought stress and its amelioration: a critical review. Plant Growth Regul. 41(1): 75-88. Crossref |
||||
| Richards RA, Hunt JR, Kirkegaard JA, Passioura JB (2014). Yield improvement and adaptation of wheat to water-limited environments in Australia-a case study. Crop Pasture Sci. 65:676–689. | ||||
|
Santos RC, Freire RMM, Suassuna TMF, Rego GM (2006). BRS Havana: nova cultivar de amendoim de pele clara. Pesq. Agropec. Bras. 41:1337‑1339. Crossref |
||||
| Silva LC, Beltrão NEM (2000) Incremento de fitomassa e produtividade do amendoinzeiro em função de lâmina e intervalos de irrigação. Rev. Bras. Oleag. Fibr. 4(2):11-121. | ||||
| Silva LC, Beltrão NEM, Rao TVR, Fideles Filho J (1998) Efeito do manejo da irrigação na qualidade da produção e na produtividade do amendoim cv. BR 1. Rev. Bras. Eng. Agríc. Ambient. 2(2):175-178. | ||||
| Somerville C, Briscoe J (2001). Genetic Eng. Water Sci. 292:2217-2231. | ||||
| Srinivasan PS, Sathasivam R, Anjuman A, Ramalingam RS, Bhat MV (1987). Effect of water stress on partitioning of dry matter and crop growth rate in relation to productivity in groundnut cultivars. J. Oilseeds Res. 4:89-96. | ||||
|
Sulc RM, Franzluebbers AJ (2014). Exploring integrated crop–livestock systems in different ecoregions of the United States. Eur. J. Agron. 57:21-30. Crossref |
||||
|
Sun MJ, Spears F, Isleib TG, Jordan DL, Penny B, Johnson D, Copeland S (2014). Effect of Production Environment on Seed Quality of Normal and High-Oleate Large Seeded Virginia-Type Peanut (Arachis hypogaea L.). Peanut Sci. 41:90-99. Crossref |
||||
| Távora JAF, Melo FIO (2001) Resposta de cultivares de amendoim a ciclos de deficiência hídrica: crescimento vegetativo, reprodutivo e relação hídricas. Rev. Ciênc. Agron. 22(2):47-60. | ||||
|
Wery J, Silim SN, Knights EJ, Malhotra RS, Cousin R (1994). Screening techniques and sources and tolerance to extremes of moisture and air temperature in cool season food legumes. Euphytica 73:73–83. Crossref |
||||
| Wu J, James Jr DW, Dooner J, Browse F (1994). A mutant of arabidopsis deficient in the elongation of palmitic acid. Plant Physiol. 29:143-150. | ||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


