ABSTRACT
The objective of this study was to assess yield loss of weed-infested cassava and the degree of interference of weeds on the crop productivity in response to the fertilization of NPK. The study was conducted at the Universidade Estadual do Sudoeste da Bahia, in the municipality of Vitória da Conquista, BA, with two different treatment groups that were evaluated in parcels with and without fertilizer. The treatments of the first group consisted of initial coexistence periods in which the cassava plants and weeds were put to live together: 35, 70, 105, 140 and 175 days after planting (DAP); in the second group, the cassava plants, initially, remain free from weeds during the same periods. Weeds were assessed every 35 days, from the 35 to 525 days after planting of cassava, determining the fresh mass of the identified species in the evaluated treatments with and without fertilizers. The characteristics of root yield, shoot weight, harvest index, dry mass of roots, starch content and flour production were evaluated 18 months after plantation. The predominant weeds in the experimental area were: Panicum maximum, Brachiaria plantaginea, Sida rhombifolia, Pavonia cancellata, Portulaca oleracea, Cynodon dactylon and Setaria parviflora. The interference of the weeds in the cassava yield was bigger when the crop was subjected to fertilization, in coexistence periods from the 35 days after planting; however, when cassava plants were kept in the absence or in coexistence with weeds up to 35 DAP, fertilization provided increase in crop productivity factors. The competition with weeds resulted in high losses in root yield of the cassava plant, thus, being necessary the control of the invading plants in the period between 35 and 175 DAP.
Key words: Coexistence period, competition, control, fertilizer, Manihot esculenta.
Cassava plant (Manihot esculenta Crantz) grows under varied conditions of climate and soils, even the ones with low fertility (Carvalho et al., 2007b). Such characteristics enable the plant to be of expressive economic and social importance in regions considered mostly unsuitable for farming, making the cassava plant an alternative of subsistence and generation of jobs and income in places where occur the least levels of human development index (HDI), in Brazil and in the world (Silva et al., 2014).
Most of the cassava crops are concentrated in small farms, under rustic management, which is characterized as a low-input system, resulting in low yields of roots in Brazil (Alves et al., 2012), with low national average of 14.6 t ha-1 (IBGE, 2014), when it is compared to the productive potential of the crop, which can reach about 90 t ha-1 of tuberous roots (Cock et al., 1979). Cardoso et al. (2013) also add that among the main reasons for low productive index of the crop is the poor technology adoption in the farming system, low yielding varieties and, mainly, the competition with weeds.
Weed in cassava cultivation has been reported as one of the main factors affecting crop yield. According to Albuquerque et al. (2008), root yield can be reduced by more than 90% in absence of weed control. This is mainly due to a slow initial growth of cassava plants, which facilitates weed species development, favoring the competition for water, light, nutrients, carbon dioxide and physical space (Azevêdo et al., 2000). In addition, cassava harvest can occur up to two years after planting, when roots are delivered to processing industry (Silva et al., 2012). Because of long cultivation and the soil partial covering by the plant, several weed infestations can occur within the planting area, what might increase crop yield losses (Johanns and Contiero, 2006).
Although, the degree of interference of weeds in the crops depends on factors related to the crop such as, the weed community, the environment and the period in which they coexist (Silva et al., 2007). In this process, one of the factors more easily controllable, in practice, is the length of the coexistence period between crop and weeds (Pitelli, 1985). According to Carvalho (2000), the degree of this competition depends on the species, the population density and, mainly, the period in which they remain growing together. Therefore, identification of the most frequent species of weed is necessary because each one – according to the potential to establish in the area and the aggressiveness – can interfere differently with the cassava plant.
Among resources liable to weed and crop competition, nutrient extraction and accumulation appear to be a crucial feature when studying the entire weed community in competition with intermediate cycle crops, such as cassava (Albuquerque et al., 2012).
Fertilizers can be used to alter competitive relationships between crop and weed, favoring crop plants by changing weed community composition and density, since the species have different responses to nutrient inputs (Armstrong et al., 1993). Even though a large amount of knowledge and technological advances on crop mineral nutrition are available, there is a lack of that regarding infesting communities, what impairs the understanding of interfering factors in competition for nutrients between weed and crop plants (Procópio et al., 2005). Based on the above, this study aimed to identify the main species of weed found during cassava cultivation, and to evaluate its effect on crop productivity in response to NPK fertilization.
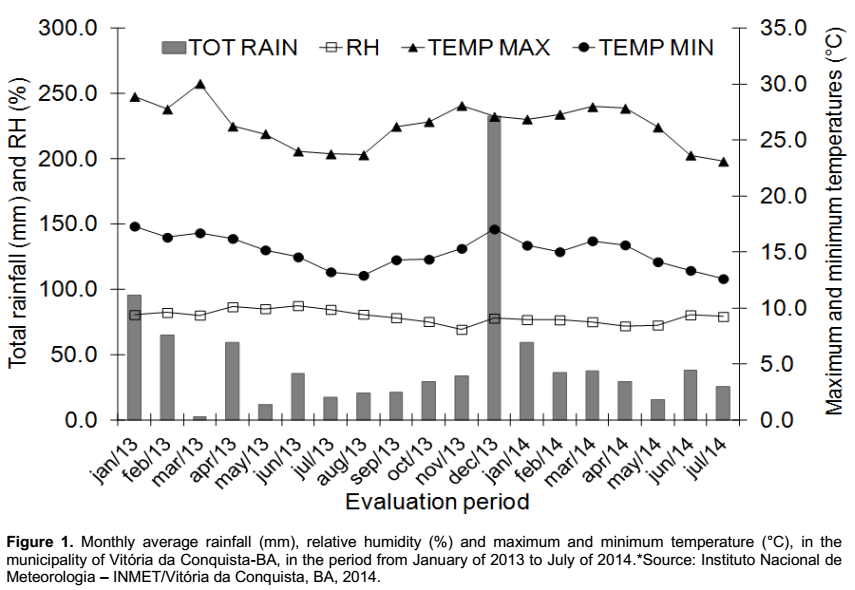
The study was developed between January of 2013 and July of 2014, in the experimental area of the Universidade Estadual do Sudoeste da Bahia, campus Vitória da Conquista, BA. The geographic coordinates are 14°51’ South and 40°50’ West, 941 m of average altitude. The climate, in accordance with the Köppen classification is Cwa (Humid subtropical climate), with annual average rainfall of 741 mm. The soil of the experimental area was classified as typical Dystrophic Yellow Latosol (Oxisol) (EMBRAPA, 2006), of which main physicochemical characteristics are shown in the Table 1. Figure 1 depicts the climatic data that were obtained during the experiment, regarding the rainfall, relative humidity, maximum and minimum temperature.
Soil tilling consisted of plowing, harrowing and grooving. Fertilization of treatments was based on soil analysis and recommendation for cassava crop proposed by Nogueira and Gomes (1999). It was applied 40 kg ha-1 P2O5, directly into planting groove; and 70 kg ha-1 N and 30 kg ha-1 K2O as top dressing, sixty days after planting. In the second year, 60 kg ha-1 N and 60 kg ha-1 K2O were applied as topdressing at the beginning of rainy season (December, 2013). For treatments without fertilizer application, it was considered soil natural fertility (Table 1).
The planting was manually conducted in January, 2013, where variety of Caitité was taken, which is a bitter variety, with approximately from 2 to 3 cm of diameter, 20 cm of length and seven buds. The spacing was 1.0 m between rows and 0.6 m between plants; each plot consisted of four lines of 8.4 m of length and 4.0 m of width, totaling 33.6 m2. The usable area of the plot was represented by two central lines, leaving out 0.6 m in each
extremity with frontal borders, with a usable area of 14.4 m2.
The experiment consisted of two treatment groups, which both were evaluated in plots with and without fertilizers, and four repetitions. In the first group, it was assessed with the different periods of coexistence of weeds and cassava plants, as specified in Table 2. After each period of coexistence, the crop remained free from competition with weeds due to manual weeding.
In the second group, the treatments consisted of different periods of weed control that were performed by manual weeding, maintaining the crop free from interference of weeds after the planting, as shown in Table 3.
The assessments of weeds were performed on the 35, 70, 105, 140, 175, 210, 280, 315, 350, 385, 420, 455, 490 and 525 days after the planting (DAP) of cassava. During these evaluations, weeds were collected through sampling. For that, a metallic square measuring 0.5 m x 0.5 m (0.25 m2) was randomly thrown on the usable area of the plot with and without fertilization. Weeds situated within the sampled areas were cut at ground level and, afterwards, they were taken to the laboratory, where the identification, counting and weighing of the fresh mass of the species were conducted.
During the harvest, manually performed 18 months (June, 2014) after the planting, were evaluated the following characteristics: root yield (t ha-1), shoot weight (t ha-1), harvest index, dry mass of the roots (%), starch content (%) and flour production (%). In order to measure the root yield, roots were collected in the usable area; then, they were cleaned and weighed on a precision scale at 0.1 g. The shoot (leaves and stem) was separated by sectioning at a height of approximately 15 cm from the soil; after, the green mass was quantified. The harvest index was evaluated by using the formula: HI = root weight ÷ (root weight + weight of the shoot). The characteristics of dry mass of the roots and starch content were determined by the hydrostatic weighing (Grossmann and Freitas, 1950), considering zero for values below three kilograms of root. The root yield was determined through the method proposed by Fukuda and Caldas (1987).
A randomized block design with four replications was used in this experiment, with the treatments of each plot arranged in a 7 x 2 factorial scheme, in which there were seven initial periods of coexistence or weed control, evaluated with and without fertilization. The obtained data related to the assessment of the crop was subjected to variance analysis, by the F-test at 5% of probability and the means were compared to each other through the Tukey’s test at 5% of probability.
Regarding the data from the assessment of the weed population, the treatments were distributed in a 15 x 2 factorial scheme (fifteen periods of evaluation and two fertilization conditions), using the F-test to compare the quadratic means and, and the means that had been compared by the Tukey’s test at 5% of probability. In order to quantify the results, a regression analysis was performed, selecting the significant models (F ≤ 0.05), with the coefficient of determination being above 60%. The statistical analysis of the data was carried out by using the software ASSISTAT, beta version 7.7.
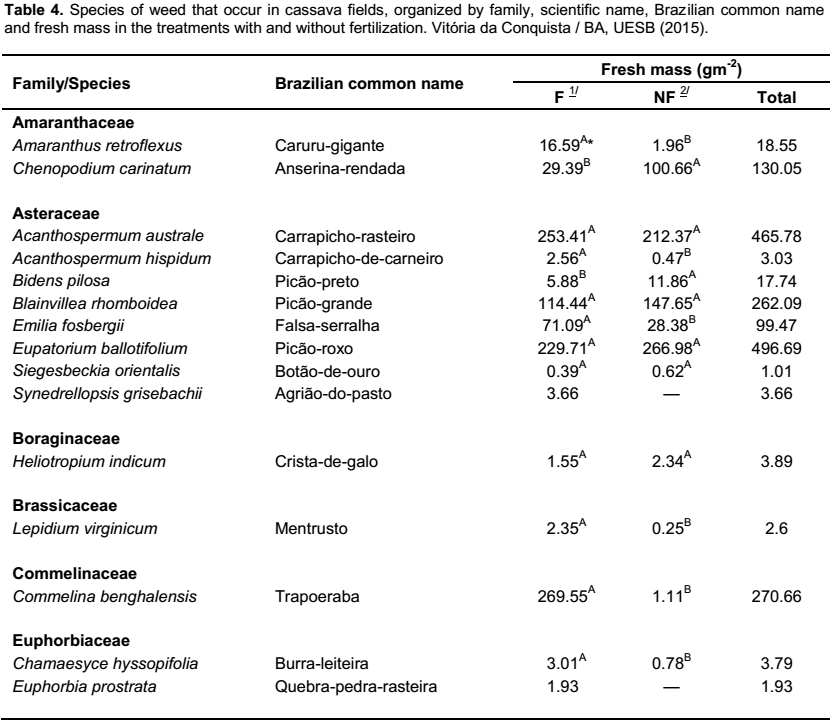
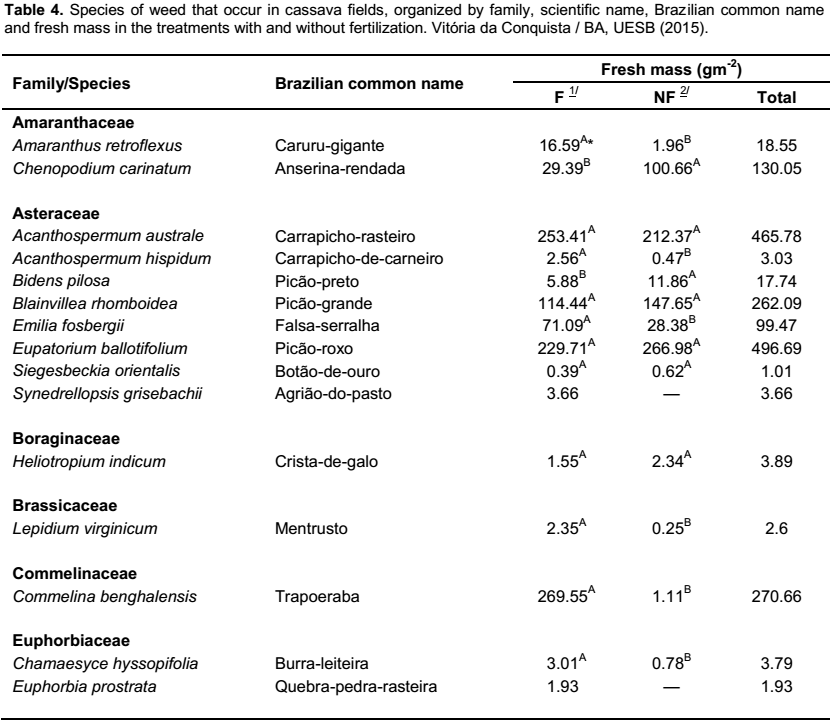
By surveying the weed population, 50 species were identified, which were classified in 39 genera and 15 botanic families. The main families, in regard to the number of species, were: Malvaceae (fourteen), Asteraceae (eight), Poaceae (seven) and Fabaceae (five), which represent
68% of the species found (Table 4). Similar results were found in surveys conducted by Otsubo et al. (2002), Albuquerque et al. (2008) and Guglieri et al. (2009), which also emphasized the mentioned families as being those of utmost number of species of weeds in cassava fields.
The composition of the weed community was considered to be heterogeneous in comparison
to the survey conducted by Albuquerque at al. (2014), who by evaluating the occurrence of weed in cassava fields located in the cerrado (tropical savanna ecoregion of Brazil) of Roraima (Boa Vista, RR), reported a population of 27 weed species, distributed in 21 genera and 8 families. Conversely, in the phytosociological survey conducted by Huziwara et al. (2009), in the municipality of Campos de Goytacazes/RJ, it was identified 10 species that belong to 9 genera and 9 families of invasive plants of cassava fields.
Regarding the fresh mass produced by the infesting community, it was observed an increase of 47.6% in the treatments with fertilizers (12,390.51 gm-2) in comparison with the treatments without fertilizers (8,393.28 g m-2) (Table 4), indicating that the application of fertilizers to cassava crops favors the growth of weeds, which may lead to larger losses in the production of roots due to the competition. According to Cruz and Pelacani (1993), among the effects caused by the presence of weeds, the shade provided by species that grew more rapidly in the initial growth phase of the crop appears to be more relevant, because as the percentage of shading on cassava grows, the plant height increases, without increasing the accumulation of biomass in the stem and with reduction of the leaf area index. These authors conclude that, with less light exposure, the dry matter of the stem and leaves and the root yield are impaired. As a consequence, shading promotes delayed growth and decrease in the growth rate of the tuberous root.
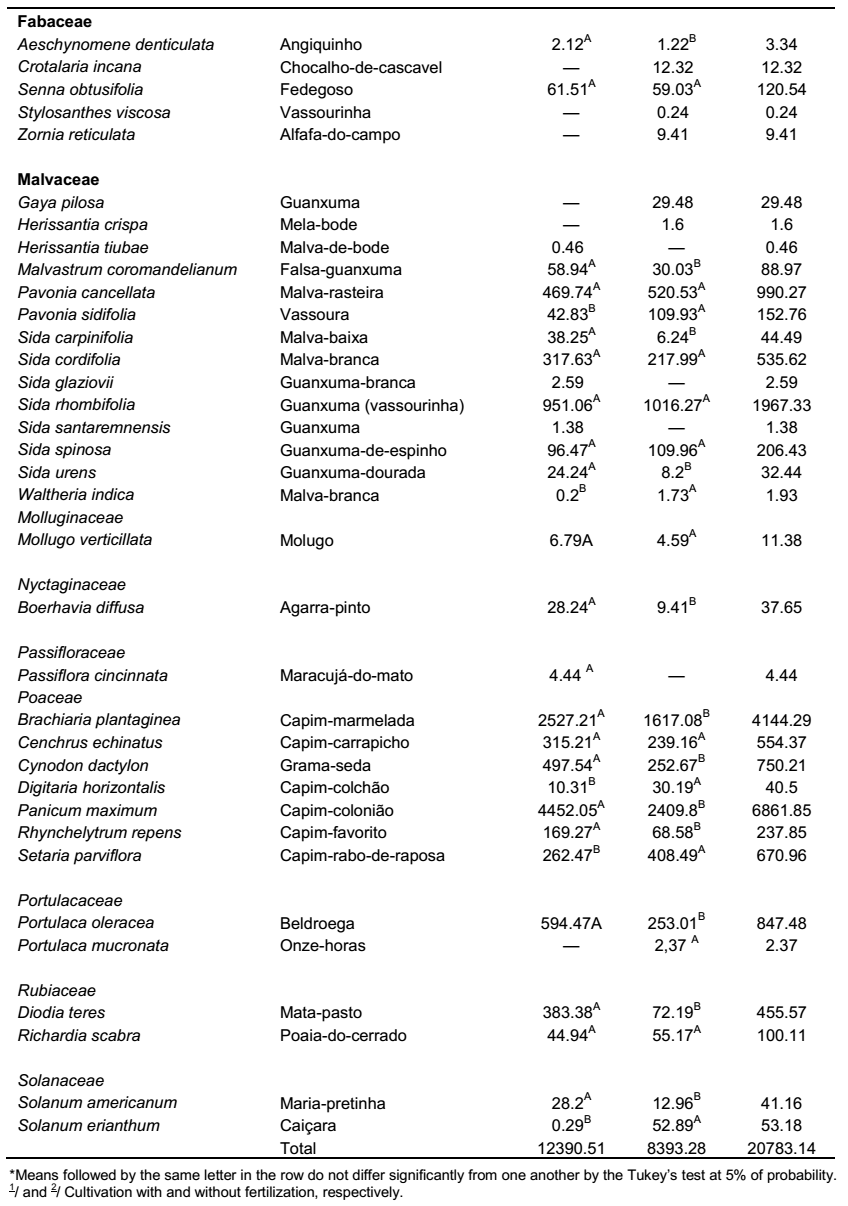
Despite being less expensive, weed control during second year can be difficult, because crop shoot has already been formed, which makes it difficult to enter into the field (Peressin and Carvalho, 2002). In this case, between the two cycles, the crop is in physiological rest. Falling leaves and plant reduced metabolic activity characterize this phase and its duration is related to environmental conditions especially. Therefore, it is during this period that a new infestation starts, which was also observed in this study, mainly in fertilized treatments from 350 DAP. It is therefore necessary to control these plants, to avoid possible losses and to facilitate crop harvesting (Silva et al., 2012).
The species with higher
production of fresh mass were:
Panicum maximum (33.02% in relation to the total mass),
Brachiaria plantaginea (19.94%),
Sida rhombifolia (9.47%),
Pavonia cancellata (4.76%),
Portulaca oleracea (4.08%),
Cynodon dactylon (3.61%) and
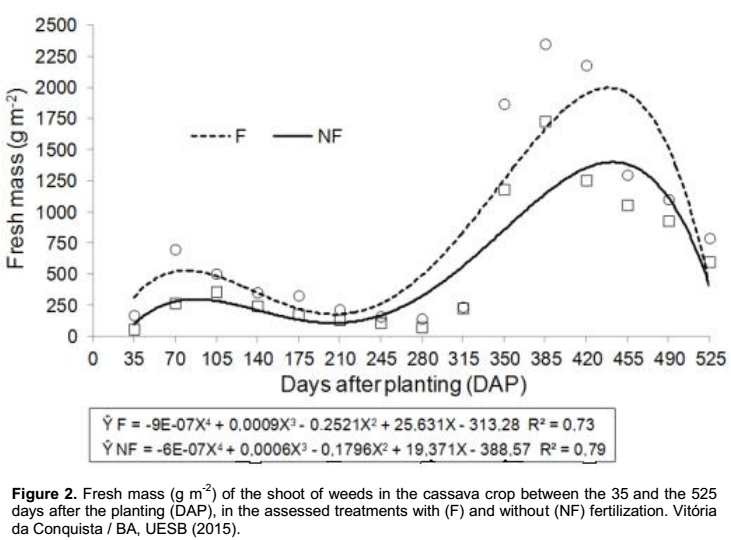
Setaria parviflora (3.23%) (Table 4). In regard to the percentage of total fresh mass produced in the treatments with and without fertilization, these species represented 78.72 and 77.17%, respectively (Figure 3). Concerning the responses of these species to fertilization,
P.
maximum,
B.
plantaginea and
C.
dactylon (Poaceae) had significant increases in fresh mass when fertilizer was applied, representing 84.74; 56.28 and 96.91%, respectively.
Contrarily, S. parviflora had higher production of fresh mass in the non-fertilized treatments, reducing this rate in 35.74% for fertilized ones. In the family of Malvaceae, S. rhombifolia and P. cancellata were unresponsive to fertilization, showing a sensitive reduction in the fresh mass in the fertilized cultivation (6.42 and 9.76%, respectively), without differing, however, from the unfertilized cultivation. As for P. oleracea (Portulacaceae) the fertilization provided an increase of 135% in yield of fresh mass in comparison with the unfertilized cultivation (Table 4). Such results demonstrate that weed responses to fertilizers are variable with regards to fresh mass production.
According to Brighenti and Oliveira (2011), some weed species have greater efficiency to use fertilizers to grow faster, increasing the competition against crop. Within the weeds of greater occurrence in the experimental area, the species P. maximum, B. plantaginea and S. rhombifolia stood out, of which exhibited higher percentage of fresh mass than the remaining, representing 64 and 60% of the total fresh mass measured in the treatments with and without fertilization, respectively (Figure 3).
The species P. maximum, commonly known as “capim-colonião”, exhibited high values of fresh mass in the shoot, being responsible for 35.93% of the fresh mass in the fertilized cultivation and 28.71% in the unfertilized one (Figure 3); suggesting high power of competitiveness due to its high biomass producing capacity in comparison to the other species. Its occurrence was recorded in some periods of the first cycle of crop, however, with reduced production of fresh mass. Nevertheless, from the 350 days after planting the cassava, phase characterized by the beginning of the local rainy season (Figure 1), it was observed a significant increase in the production of fresh mass in the plant shoot.
Among the probable factors that are responsible for the occurrence and elevated production of fresh mass in “capim-colonião”, the following stand out: the presence of propagules of the species in seedbanks in the area, defoliation of cassava plants, fertilizer supply and beginning of the rainy season; these were the conditions that, certainly, favor the establishment and the development of P. maximum in the area, since the species is very light-demanding, fertility and soil moisture.
B. plantaginea, commonly known as “capim-marmelada”, exhibited a good adaptation and aggressiveness in the cultivated area, being found in all the assessments of the infesting community; representing 20.39% of the total fresh mass obtained in the fertilized cultivation and 19.26% in the unfertilized cultivation (Figure 3). This Poaceae, originated from Africa, has the seed as the main way of propagation, which is characterized by exhibiting primary dormancy in the maturation process (Lorenzi, 2008); hence, the germination is distributed throughout the time, which impairs its control (Kissmann, 1997).
The high competing potential of the “capim-marmelada” was also verified in the study performed by Aspiazú et al. (2010), in which by evaluating the efficiency in watering cassava plants in coexistence with weed, it was verified that the B. plantaginea is very efficient at using water, principally for having a metabolism C4, and it remains competing with the cassava even under temporary conditions of shortage of water.
The Poaceae family is highlighted as one of the most important group in cassava fields. Pinotti et al. (2010) identified the species Brachiaria decumbens and Digitaria horizontalis are economically important in cassava fields in the municipality of Pompéia-SP. Whereas Albuquerque et al. (2014), in a phytosociological study on the cassava crop in the cerrado (tropical savanna ecoregion of Brazil) of Roraima, concluded that the species that exhibited the highest values of dry mass were the Poaceae Digitaria sanguinalis, Brachiaria brizantha, B. decumbens and Brachiaria humidicola. As reported by Maciel et al. (2010), many species of the Poaceae family are perennial and produce a large quantity of seeds, increasing their dissemination and colonization of different environments.
The species S. rhombifolia, commonly known as “guanxuma”, exhibited widespread occurrence in the experimental area, being recorded in every assessment of the infesting community; accounting for 7.67% of the total fresh mass that was produced by the fertilized cultivation and 12.1% in the unfertilized one (Figure 3). Such occurrence may be due to its high potential of infestation since this species exhibits high yield of seeds and easy dispersion.
The yield of fresh mass of “guanxuma” was larger in the second cycle of the crop, after the 350 days after planting of cassava, however, without considerable differences among the obtained values in the cultivation with and without fertilizers; demonstrating that the occurrence and production of fresh mass of this species are independent of the supply of fertilizer to the crop. Conversely, this performance can be explained by the higher efficiency of the Poaceae P. maximum and B. plantaginea in using the fertilizer supplied to the crop and the environment, and, consequently, in the production of fresh mass; which may have not favored the growth of the S. rhombifolia in the fertilized cultivation, in relation to the unfertilized one.
S. rhombifolia occurs in annual and perennial crops, being highly competitive due to its radicular system, which can reach 50 cm in depth (Kissmann and Groth, 2000; Lorenzi, 2008). Reports indicated that this plant can yield up to 28.2 thousand seeds per m-2 in only a cycle in the summer as a weed in soybean fields (Fleck et al., 2003). Within the species of “guanxuma” that occur in Brazil, it is considered the most disseminated species and its control is the most problematic, regardless the agricultural environment (Constantin et al., 2007). It was also reported as weed in cassava fields by Azevêdo et al. (2000) and Albuquerque et al. (2008), corn fields (Macedo et al., 2003), sugarcane fields (Oliveira and Freitas, 2008) and soybean field (Voll et al., 2005).
Generally, the application of fertilizers on cassava fields, performed as to support the crop that is suffering detrimental effects of weeds, also favored the production of fresh mass of weeds, chiefly, in the second year of the crop cycle; therefore, depending on the intensity of the competition in this phase, the crop might be negatively affected, because according to Procópio et al. (2005), depending on the management, the application of macronutrients may benefit more the species of weed than the crop.
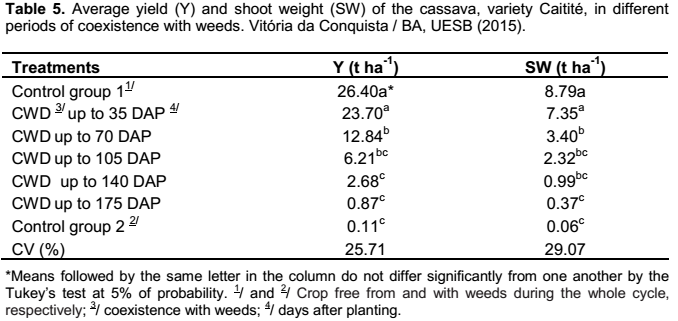
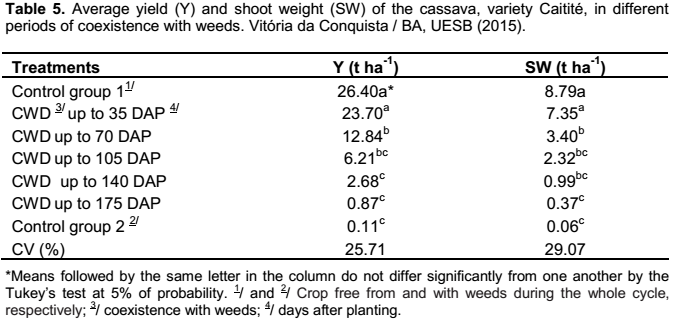
Regarding the first group of treatments (periods of coexistence of crop and weeds), the root yield and the shoot weight of the cassava plant were not influenced by the fertilizer provided to the crop, being only influenced by the different periods of coexistence with the weed community (Table 5). By evaluating the yield, it was observed the decline in root production when the periods of coexistence of weeds and crop were similar or superior to 70 days between the planting and the weeding. The yield loss was about 90% when the first weeding was executed after the 140 days of coexistence of crop and weeds (Table 5).
The highest productivities of roots were observed in the control group free from competition and in the treatment in which the coexistence of crop and the weeds occurred up to 35 days after the planting (Table 5). Similar results to the ones found in this study were also verified by Carvalho et al. (2004), as they found out that the cassava crop, in the municipality of Cruz das Almas, Bahia, can coexist with weeds for a period from 20 to 30 days after the sprouting, about 35 to 45 days after planting, without significant loss in the production of roots.
The weeding that was performed after 70 days of coexistence of crop and weeds resulted in losses of 51% in productivity of cassava, in relation to that obtained in the cultivation kept free from weeds during the whole cycle (Table 5). Alcântara et al. (1983) also reported the that weeding on the 60 days after the sprouting showed reduction in the root and shoot yield, which accorded with the data obtained by Carvalho et al. (1990).
According to Johanns and Contiero (2006), in a study carried out in Marechal Cândido Rondon-PR, it was found out that the competition of weeds and cassava, cultivar “Fécula Branca”, between the 60 and 90 days after the planting reduces significantly the yield of the crop, concluding that, in this period, there is more competition for resources. Albuquerque et al. (2008), in Viçosa-MG, reported that the cassava crop, cultivar “Cacauzinha”, is more sensitive to competition in periods of coexistence between the 25 and 75 days after the planting. Biffe et al. (2010), in Maringá-PR, found out that weeding performed up to 100 days after planting considerably increased the production of cassava roots, cultivar “Fécula Branca”.
The results verified in this study in which the lowest productivities were obtained in the treatments with weeding done after 140 days of coexistence and in the control group without weeding (Table 5), accord to some authors, which the cases where is reported the greater interference of weeds, the productivities are below 10% of the results obtained in the weeded control (Carvalho et al., 1993; Moura, 2000; Johanns and Contiero, 2006; Silva et al., 2012).
In shoot weight, it was noted that periods of coexistence with weeds from the 70 days after planting cassava considerably reduced the production of the plant shoot, and, consequently, the plant growth (Table 5). This small growth of the cassava shoot contributes to the decrease in photosynthetic tissue and, eventually a larger accumulation of carbohydrates in the roots, reducing the final yield of the crop (Viana et al., 2001).
In both cultivation, with and without fertilizers, the harvest index of the crop exhibited significant reduction from the 105 days of coexistence with weeds, with the less expressive results found in the treatments with CWD up to 175 DAP and in the control group with no weeding; however, despite the losses due to competition, superior indexes were observed in the unfertilized cultivation, more specifically in the treatments CWD up to 70 DAP, up to 105 DAP and in the control group kept in the among the weeds during the whole cassava cycle, in relation to the fertilized cultivation (Table 6).
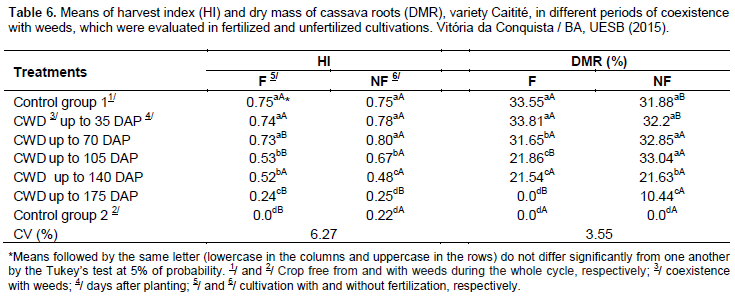
In relation to the percentage of dry mass of roots, in the fertilized cultivation, the losses were recorded starting from the 70 days of coexistence of crop and weeds; whereas in the unfertilized cultivation, these losses were retarded, being only verified in periods of coexistence starting from the 140 days after the planting cassava. In both cases, the least impressive results were verified in the treatments CWD up to 175 DAP and in the control group without weeding.
In the fertilized cultivation, it was recorded that the percentages of dry mass were higher than the ones found in the unfertilized cultivation, in the control group free from weeds and in the initial period of coexistence of crop and weeds (CWD up to 35 DAP). In the unfertilized cultivation, relating to the fertilization, higher percentages were obtained in longer periods of coexistence, namely the treatment CWD up to 105 DAP and up to 175 DAP (Table 6).
These results suggest that the fertilization with NPK alters the degree of competitiveness between the crop and the weed community, favoring the growth of species of weed after the 35 days after the planting of cassava, leading to the retardation of the beginning of the formation of roots and, consequently, the accumulation of dry mass. A similar behavior was observed by Pereira et al. (2012), in which was found out that while evaluating the growth of cassava and weeds in response to phosphorous fertilization the fertilization tends to contribute to a faster growing rate for weeds than for cassava. To Carvalho et al. (2007a), there is a greater development of the roots in the initial periods of the development cycle of weeds, resulting in an increase of nutrient uptake by radicular interception, leading to a quick accumulation of nutrients by the roots and, consequently, a lower availability for the crop.
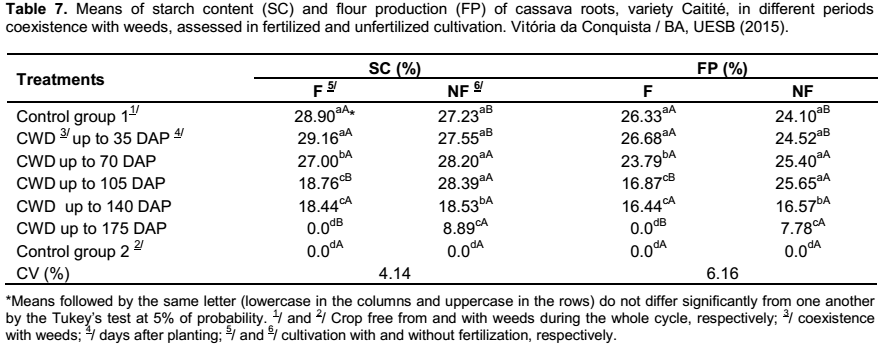
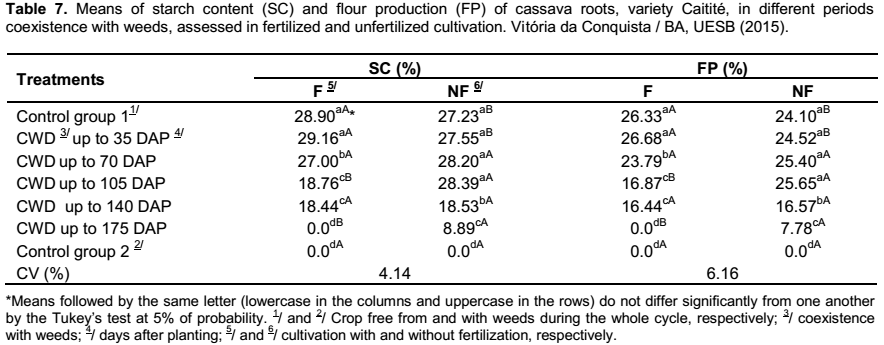
In respect to the starch content and the flour production, as similar as the results observed for the dry mass of roots, in the fertilized cultivation, it was verified a decrease of these values when the periods of coexistence of crop and weeds were the same or superior to 70 days between the planting and the weeding. In cultivation without fertilization , the losses were recorded only after the 140 days of coexistence of crop and weeds. In both cultivation, the lowest results of starch content and flour production were observed in the treatments CWD up to 175 DAP and in the control group without weeding (Table 7). The results are in accordance with Silva et al. (2012), which affirmed that the presence of weeds in competition with the cassava crop may reduce the percentages of dry mass and starch content in the roots.
The effect of fertilization in the control group free from weeds and in the treatment CWD up to 35 DAP were recorded higher percentages of starch content and flour production, than the unfertilized cultivation. Conversely, in the treatments of coexistence of crop and weeds (up to 105 DAP and up to 175 DAP), in the unfertilized cultivation, it was obtained results that were higher than those found in the fertilized cultivation (Table 7); indicating that the effects of competition are sharp when the cultivation of cassava is submitted to fertilization.
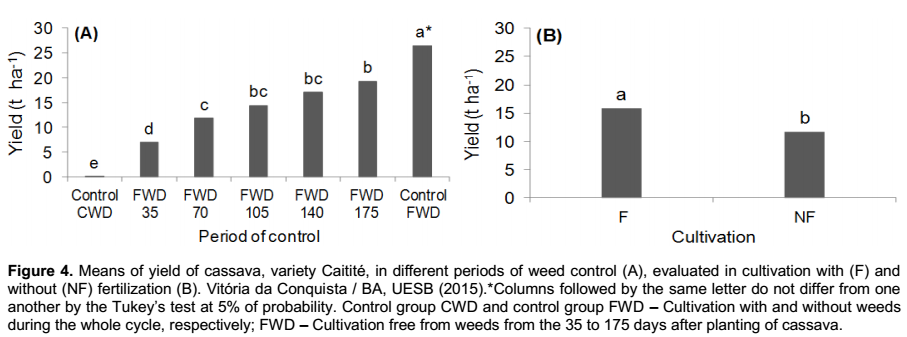
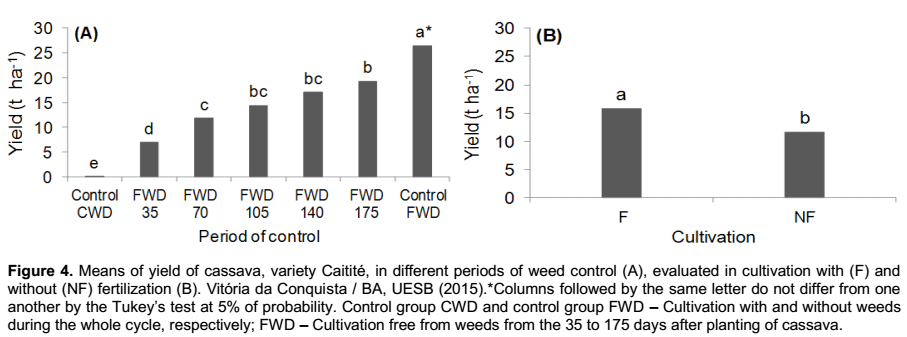
Regarding the root yield that was obtained in the second group of treatments (initial periods of weed control), it was verified that there was no significant interaction between the periods of weed control and the fertilization, occurring only isolated effects of these factors (Figure 4).
By evaluating the productivity of cassava in the different periods of weed control, it was verified that the cultivation free from weeds during the whole cycle has the greater results, with 26.4 t ha-1, 37% more than the productivity found in the treatment free from weeds up to the 175 days after the planting of cassava; showing, in this case, that the management of the crop in clean areas during the whole cycle leads to the maximum economical potential of yielding roots (Figure 4A). The differences in productivity found within the treatments are probably due to a recurring infestation of weeds in the second year of the crop cycle (350 DAP) as a result of the beginning of the local rainy season (Figure 1), resulting in restarting the growth of weeds, as observed in the increase in fresh mass in this period (Figure 2), and the resumption of the competition with the crop. Therefore, in cassava fields with cycles longer than a year, it must be planned strategies of weed control that also cover the second year of the crop cycle.
Different results were found by Carvalho et al. (2004), in which, by studying the cultivar “Cigana preta” (BMG 116), in the municipality of Cruz das Almas, Bahia, indicate the necessity of weed control up to 135 days after planting the cassava. However, the periods of weed control are not absolute and they vary a lot, as it depends on factors that affect the production system, such as the cultivation spacing, species and population density of weeds, used cultivar, farm management, and hydric regime, among others. Therefore, the periods of weed control must be analyzed considering the location, environment characteristics and the management of the cassava field (Silva et al., 2012).
The yield of the cultivation that was kept with weeds during the whole cycle was severely affected, with losses exceeding 95% in comparison with the one free from weeds (Figure 4A). These losses are in accordance with the ones reported by Peressin et al. (1998) and Mattos and Cardoso (2005), as they affirmed that the losses in root yield as a result of competition with weeds during the cultivation of cassava may reach more than 90%, depending on the length of the coexistence and population density of weeds.
The fertilization boosted root yield by 35% in comparison with the unfertilized cultivation (Figure 4B), indicating that the improvement in soil fertility directly affects the cassava productivity. A similar effect was observed by Cardoso et al. (2005), which by studying the effect of nitrogen fertilization (0, 50, 100, 200, 300 and 400 kg ha-1 of N) on the agronomic characteristics of cassava, variety “Sergipe”, in the municipality of Vitória da Conquista-BA, it was verified that the increase in root yield in relation to the amount of nitrogen applied, reaching up to 22.1 t ha-1 when it is applied 400 kg of N, which represents an increase of 20% in comparison with the unfertilized cultivation.
Likewise, Alves et al. (2012), by evaluating the effect of NPK (0, 200, 400 and 600 kg ha-1) on the root yield, in the municipality of Moju-PA, it was concluded that in sandy soils and of low fertility, the variety of cassava “Paulozinho” responded linearly to the growing amounts of mineral fertilizer NPK. Nonetheless, Fidalski (1999) verified that only the phosphate fertilization (0, 30, 60, 90 and 120 kg ha-1 of P2O5) contributed to the increase in root yield of cassava, cultivar “Fibra”, when it is submitted to NPK fertilization in sandy soils in the northwest of Paraná.
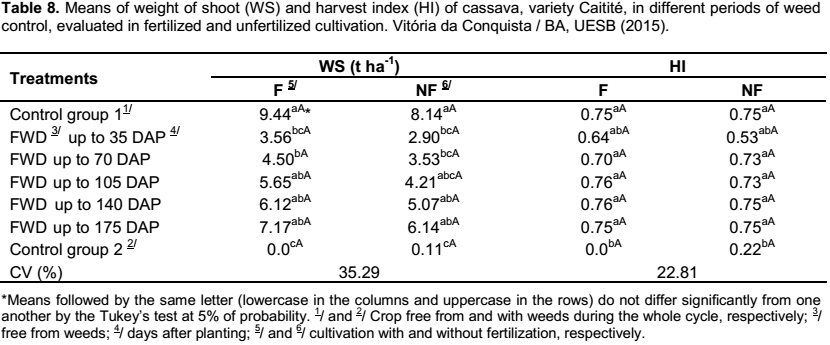
Relating to the weight of the shoot and the harvest index of the crop, it was observed that there were no differences between the fertilized and the unfertilized cultivation for the periods of weed control, demonstrating that the application of fertilizers to the crop has no influence on these variables (Table 8).
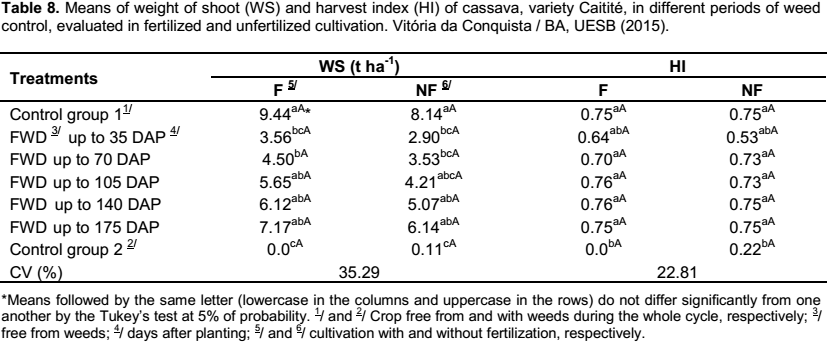
In both cultivations, with and without fertilization, it was verified that as the periods of weed control increased, there was an increase in weight of the shoot of cassava, with the higher result observed in the treatment that was kept free from weeds during the whole crop cycle, although not differing from the treatments FWD up to 105 DAP, up to 140 DAP and up to 175 DAP (Table 8). The data accord with the ones obtained by Albuquerque et al. (2008), in which the cultivation kept free from weeds exhibited the highest weight of the shoot, though not differing from the treatments free from weeds up to 75, 100 and 125 days after planting the cassava, cultivar “Cacauzinha”. According to Silva et al. (2012), the production of the plant shoot is of great importance for the cassava production, supplying material to the propagation and to the production of forage, besides of being the part of the plant responsible for absorbing light and providing photoassimilates to the roots.
As for the harvest index, in the fertilized and unfertilized cultivation, except for the treatment kept with the weeds during the whole crop cycle, the remaining treatments did not differ from one to another (Table 8). In accordance with Conceição (1986), a good harvest index should be at least of 60%; therefore, the treatments exhibited a good harvest index. The fact that the harvest index remained unchanged for the majority of the treatments shows that the reduction of the plant shoot is closely linked to the decrease in root yield, that is, the weeds influenced the general growth of the cassava crop.
According to Moreira et al. (2014), the harvest index, alone, does not provide the precise information about the performance of the cassava plant, as the high values of this index can either be obtained by the increase of the production of roots or by the decrease of production of the shoot.
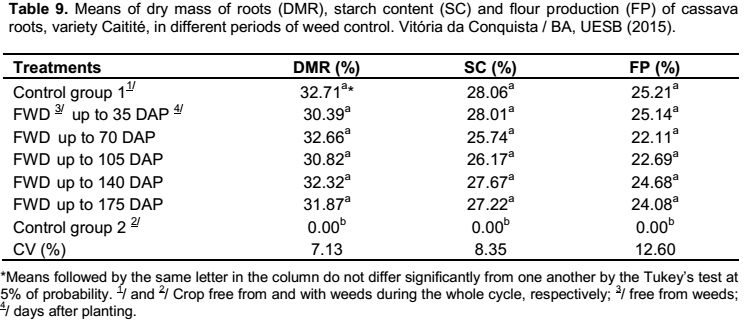
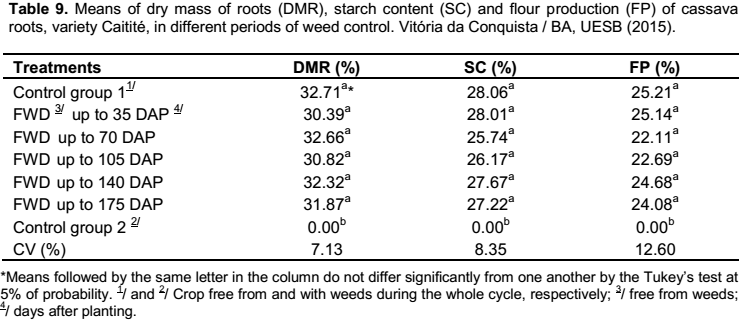
For the variables of dry mass of roots, starch content and flour production regarding the second group of treatments, there was no influence of fertilization, only of the different periods of weed control (Table 9). On the contrary, Cardoso et al. (2005) studied the effects of nitrogen fertilization on these characteristics, however, there were minor increases, which according to the authors, it is somewhat not economical advantageous.
When the crop was kept without weeding during the whole cycle, the production of roots remained below 3.0 kg ha
-1, making it inviable to determine the percentages of dry mass, starch content and flour production; while the remaining treatments did not differ from one to another (Table 9). These results
showed that these characteristics are not influenced by the presence of weeds up to the 175 days after planting cassava. Similar results were obtained by Albuquerque et al. (2008), which the periods of weed control (25, 50, 75, 100 and 125 DAP) did not differ from each other for these characteristics when cultivating the cassava cultivar “Cacauzinha”.
According to Correia et al. (2005), cassavas have about 30 to 40% of dry mass in tuberous roots however, the amount of this content relies on factors, such as: variety, age of plants, soil, planting conditions, climate and plant health. The content of dry mass is the characteristic that determines the higher or lower industrial yield of roots, since it is directly related to several products that are derived from cassava (Vidigal-Filho et al., 2000).
Generally, by analyzing the experiment as a whole, it was verified that although cassavas kept free from weeds during the whole cycle have exhibited the higher productivity, it is known that this is not a recommended practice, as maintaining the soil totally free from weeds, there is an increase in erosion and also a higher production cost. According Aguiar to et al. (2011), a significant parcel of the production cost can be attributed to the weed control, which can vary according to the species of weed and their population density. Therefore, this study, for the studied conditions, the first weeding should be performed near to 35 DAP and the last ones, near to 175 DAP.
The fertilization of cassava, when competing with weeds, provided an increase of productive factors (dry mass, starch content and flour production) when the period of coexistence was up to 35 DAP, though, in longer periods of coexistence after the planting, the losses by competitions were greater than the losses found in the unfertilized cultivation.
In conclusion it can be said that weed community found in cassava plantation was heterogeneous with 50
species distributed into 39 genera and 15 botanic families, and predominated by P. maximum, B. plantaginea, S. rhombifolia, P. cancellata, P. oleracea, C. dactylon, and S. parviflora.
The use of fertilizers in cassavas provided more production of fresh mass of weeds, mainly, in the second year of the crop that is, from the 350 DAP, which was found maximum in P. maximum and B. plantaginea.
The interference of weeds on the cassava yield was higher when the crop was subjected to fertilization with NPK, in periods of coexistence starting from the 35 DAP. Moreover, the fertilization promoted the increase of factors related to productivity like, dry mass, starch content and flour production either in absence or in coexistence with weeds up to 35 DAP. The competition with weeds resulted in high losses of root yield of cassava; thus, it is necessary to control these plants in a period situated somewhere between the 35 and 175 DAP.
The authors have not declared any conflict of interests.
REFERENCES
|
Aguiar EB, Bicudo SJ, Curcelli F, Figueiredo PG, Cruz SCS (2011). Épocas de poda e produtividade da mandioca. Pesqui. Agropec. Bras. 46(11):1463-1470.
Crossref
|
|
|
|
Albuquerque JAA, Evangelista MO, Mates APK, Alves JMA, Oliveira NT, Sediyama T, Silva AA (2014). Occurrence of weeds in cassava savanna plantations in Roraima. Planta Daninha 32(1):91-98.
Crossref
|
|
|
|
|
Albuquerque JAA, Sediyama T, Silva AA, Alves JMA, Finoto EL, Neto FA, Silva GR (2012). Desenvolvimento da cultura de mandioca sob interferência de plantas daninhas. Planta Daninha 30(1):37-45.
Crossref
|
|
|
|
|
Albuquerque JAA, Sediyama T, Silva AA, Carneiro JES, Cecon PR, Alves JMA (2008). Interferência de plantas daninhas sobre a produtividade da mandioca (Manihot esculenta). Planta Daninha 26(2):279-289.
Crossref
|
|
|
|
|
Alcântara EN, Carvalho JEB, LIMA PC (1983). Determinação do período crítico de competição das plantas daninhas com a cultura da mandioca (Manihot esculenta Crantz). Belo Horizonte: EPAMIG,
|
|
|
|
|
Alves RNB, Modesto Júnior MS, Ferreira ER (2012). Doses de NPK na adubação de mandioca (Manihot esculenta L.) variedade Paulozinho em Moju-Pará. Revista Raízes e Amidos Tropicais 8: 65-70.
|
|
|
|
|
Armstrong RD, Brown RF, Helyar KR (1993). The use of nitrogen, phosphorus and lime to limit the competitive ability of Aristida armata in the establishment phase. Aust. J. Agric. Res. 44(1):167-178.
Crossref
|
|
|
|
|
Aspiazú I, Sediyama T, Ribeiro Júnior JI, Silva AA, Concenco G, Ferreira EA, Galon L, Silva AF, Borges ET, Araújo WF (2010). Water use efficiency of cassava plants under competition conditions. Planta Daninha 28(4):699-703.
Crossref
|
|
|
|
|
Azevêdo CLL, Carvalho JEB, Lopes LC, Araújo AMA (2000). Levantamento de plantas daninhas na cultura da mandioca, em um ecossistema semi-árido do Estado da Bahia. Magistra 12(1):41-49.
|
|
|
|
|
Biffe DF, Constantin J, Oliveira Júnior RS, Franchini LHN, Rios FA, Blainski E, Arantes JGZ, Alonso DG, Cavalieri SD (2010). Período de interferência de plantas daninhas em mandioca (Manihot esculenta) no Noroeste do Paraná. Planta Daninha 28(3):471-478.
Crossref
|
|
|
|
|
Brighenti AM, Oliveira MF (2011). Biologia de plantas daninhas. In: Oliveira Júnior RS, Constantin J, Inoue MH (Eds.). Biologia e manejo de plantas daninhas. Curitiba: Omnipax, pp. 1-36.
|
|
|
|
|
Cardoso AD, Viana AES, Barbosa RP, Teixeira PRG, Cardoso JNS, Fogaça JJNL (2013). Levantamento fitossociológico de plantas daninhas na cultura da mandioca em Vitória da Conquista, Bahia. Biosci. J. 29(5):1130-1140.
|
|
|
|
|
Cardoso JNS, Viana AES, Matsumoto SN, Sediyama T, Carvalho FM (2005). Efeito do nitrogênio em características agronômicas da mandioca. Bragantia 64(4):651-659.
Crossref
|
|
|
|
|
Carvalho FM, Viana AES, Matsumoto SN, Rebouças TNH, Cardoso CEL, Gomes IR (2007b). Manejo do solo em cultivo com mandioca em treze municípios da região sudeste da Bahia. Ciência e Agrotecnologia 31(2):378-384.
Crossref
|
|
|
|
|
Carvalho JEB (2000). Plantas daninhas e seu controle. In: Mattos PLP, Gomes JC (Coords.). O cultivo da mandioca. Cruz das Almas: Embrapa Mandioca e Fruticultura, pp. 42-52.
|
|
|
|
|
Carvalho JEB, Araújo AMA, Azevedo CLL (2004). Período de controle de plantas infestantes na cultura da mandioca no Estado da Bahia. Cruz das Almas: Embrapa Mandioca e Fruticultura. (Comunicado Técnico, 109). P 7
|
|
|
|
|
Carvalho JEB, Caldas RC, Costa Neto AO, Cardoso SS, Mascarenhas L, Barbosa CV (1993). Período crítico de competição das plantas daninhas com a cultura da mandioca em um ecossistema do Nordeste brasileiro. Rev. Bras. de Mandioca 12(1):85-93.
|
|
|
|
|
Carvalho JEB, Lyra Filho HP, Caldas RC, Pereira RC, Queiroz GM, Alves AAC, Rezende GO (1990). Período crítico de competição das plantas daninhas com a cultura da mandioca em três ecossistemas do Nordeste brasileiro. Rev. Bras. de Mandioca 9(1):29-40.
|
|
|
|
|
Carvalho LB, Bianco S, Pitelli RA, Bianco MS (2007a) Estudo comparativo do acúmulo de massa seca e macronutrientes por plantas de milho var. BR-106 e Brachiaria plantaginea. Planta Daninha 25(2):293-301.
Crossref
|
|
|
|
|
Cock JH, Franklin D, Sandoval G, Juri P (1979). The ideal cassava plant for maximum yield. Crop Sci. 19:271-279.
Crossref
|
|
|
|
|
Conceição AJ (1986). A mandioca. São Paulo: Nobel, P 382.
|
|
|
|
|
Constantin J, Oliveira Júnior RS, Kajihara LH, Arantes JGZ, Cavalieri SD, Alonso DG (2007). Controle de diferentes espécies de guanxumas com aplicações sequenciais de flumiclorac-pentil. Acta Sci. Agron. 29(4):475-480.
Crossref
|
|
|
|
|
Correia AD, Farias ARN, Mattos, PLP (2005). Utilização da mandioca e de seus produtos na alimentação humana. In: Souza LS, Farias ARN, Mattos PLP, Fukuda WMG (Eds.). Processamento e utilização da mandioca. Cruz das Almas: Embrapa Mandioca e Fruticultura Tropical. Cap. 7:221-298.
|
|
|
|
|
Cruz JL, Pelacani R (1993). Fisiologia da mandioca. In: Curso Nacional de Mandioca. Cruz das Almas. Anais. Cruz das Almas: Embrapa/CNPMF, P 38.
|
|
|
|
|
EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA – EMBRAPA (2006). Sistema brasileiro de classificação de solos. Rio de Janeiro: Embrapa Solos P 412.
|
|
|
|
|
Fidalski J (1999). Respostas da mandioca à adubação NPK e calagem em solos arenosos do noroeste do Paraná. Pesqui. Agropec. Bras. 34(8):1353-1359.
Crossref
|
|
|
|
|
Fleck NG, Rizzardi MA, Agostinetto D, Vidal RA (2003). Produção de sementes por picão-preto e guanxuma em função de densidades das plantas daninhas e da época de semeadura da soja. Planta Daninha 21(2):191-202.
Crossref
|
|
|
|
|
Fukuda WMG, Caldas RC (1987). Relação entre os conteúdos de amido e farinha de mandioca. Rev. Bras. de Mandioca 6:57-63.
|
|
|
|
|
Grossmann J, Freitas AC (1950). Determinação do teor de matéria seca pelo peso específico em raízes de mandioca. Rev. Agron. 14(160/162):75-80.
|
|
|
|
|
Guglieri A, Caporal FJM, Vinci-Carlos HC, Pinto BEM (2009). Fitossociologia de plantas espontâneas em um mandiocal implantado em pastagem cultivada em Mato Grosso do Sul, Brasil. Rev. Ciênc. Agrárias 51:127-141.
|
|
|
|
|
Huziwara E, Ogliari J, Freitas SP, Paes HMF, Lemos GCS (2009). Levantamento fitossociológico de plantas daninhas na cultura da mandioca no município de Campos dos Goytacazes, RJ. Rev. Raízes Amidos Trop. 5(1):468-472.
|
|
|
|
|
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA - IBGE (2014). Disponível em:
View. Acesso em: 20 de dezembro de 2014.
|
|
|
|
|
Johanns O, Contiero R (2006). Efeitos de diferentes períodos de controle e convivência de plantas daninhas com a cultura da mandioca. Rev. Ciênc. Agron. 37(3):326-331.
|
|
|
|
|
Kissmann KG (1997). Plantas infestantes e nocivas. 2.ed. São Paulo: Basf Brasileira. Tomo I. pp. 415-420.
|
|
|
|
|
Kissmann KG, Groth D (2000). Plantas infestantes e nocivas. 2.ed. São Paulo: BASF, Tomo III, P 723.
|
|
|
|
|
Lorenzi H (2008). Plantas daninhas do Brasil: terrestres, aquáticas, parasitas e tóxicas. 4.ed. Nova Odessa: Plantarum, P 640.
|
|
|
|
|
Macedo JF, Brandão M, Lara JFR (2003). Plantas daninhas na pós-colheita de milho nas várzeas do Rio São Francisco, em Minas Gerais. Planta Daninha 21(2):239-248.
Crossref
|
|
|
|
|
Maciel CDC, Poletine JP, Oliveira NAM, Guerra N, Justiniano W (2010). Levantamento fitossociológico de plantas daninhas em calçadas do município de Paraguaçu Paulista-SP. Planta Daninha 28(1):53-60.
Crossref
|
|
|
|
|
Mattos PLP, Cardoso EMR (2005). Plantas daninhas. Disponível em:
|
|
|
|
|
Moreira GLP, Viana AES, Cardoso AD, Santos VS, Matsumoto SN, Andrade ACB (2014). Intervalos entre podas de duas variedades de mandioca. Biosci. J. 30(6):1757-1767.
|
|
|
|
|
Moura GM (2000). Interferência de plantas daninhas na cultura da mandioca (Manihot esculenta Crantz) no Estado do Acre. Planta Daninha 18(3):451-456.
Crossref
|
|
|
|
|
Nogueira FD, Gomes JC (1999). Mandioca. In Ribeiro AC, Guimarães PTG, Alvarez Viegas VH (Eds.). Recomendações para uso de corretivos e fertilizantes em Minas Gerais: 5ª aproximação. Viçosa, Comissão de Fertilidade do Solo do Estado de Minas Gerais, pp. 312-313.
|
|
|
|
|
Oliveira AR, Freitas SP (2008). Levantamento fitossociológico de plantas daninhas em áreas de produção de cana-de-açúcar. Planta Daninha 26(1):33-46.
Crossref
|
|
|
|
|
Otsubo AA, Mercante FM, Martins CS (2002). Aspectos do cultivo da mandioca em mato Grosso do Sul. Dourados: Embrapa Agropecuária Oeste; Campo grande: UNIDERP. P 219.
|
|
|
|
|
Pereira GAM, Lemos VT, Santos JB, Ferreira EA, Silva DV, Oliveira MC, Menezes CWG (2012). Crescimento de mandioca e plantas daninhas em resposta à adubação fosfatada. Rev. Ceres 59(5):716-722.
Crossref
|
|
|
|
|
Peressin VA, Carvalho JEB (2002). Manejo integrado de plantas daninhas em mandioca. In: Marney Pascoli Cereda. (Org.). Cultura de Tuberosas Amiláceas Latino Americanas. São Paulo: Fundação Cargill 2:302-349.
|
|
|
|
|
Peressin VA, Monteiro DA, Lorenzi JO, Durigan JC, Pitelli RA, Perecin D (1998). Acúmulo de matéria seca na presença e na ausência de plantas infestantes no cultivar de mandioca SRT 59 - Branca de Santa Catarina. Bragantia 57(1):135-148.
Crossref
|
|
|
|
|
Pinotti EB, Bicudo SJ, Curcelli F, Dourado WS (2010). Levantamento florístico de plantas daninhas na cultura da mandioca no município de Pompéia – SP. Revista Raízes e Amidos Tropicais 6:120-125.
|
|
|
|
|
Pitelli RA (1985). Interferência de plantas daninhas em cultivos agrícolas. Informe Agropecuário 11(1):16-26.
|
|
|
|
|
Procópio SO, Santos JB, Pires FR, Silva AA, Mendonça ES (2005). Absorção e utilização do fósforo pelas culturas da soja e do feijão e por plantas daninhas. Rev. Bras. Ciênc. Solo 29(6):911-921.
Crossref
|
|
|
|
|
Silva AA, Ferreira FA, Ferreira LR, Santos JB (2007). Biologia de plantas daninhas. In: Silva AA, Silva JF. Tópicos em manejo de plantas daninhas. Viçosa: UFV, pp. 17-62.
|
|
|
|
|
Silva DV, Santos JB, Ferreira EA, Silva AA, França AC, Sediyama T (2012). Manejo de plantas daninhas na cultura da mandioca. Planta Daninha 30(4):901-910.
Crossref
|
|
|
|
|
Silva DV, Silveira HM, Ferreira EA, Carvalho FP, Castro Neto MD, Silva AA, Sediyama T (2014). Aspectos fisiológicos da mandioca após a aplicação dos herbicidas fluazifop-p-butil e fomesafen. Rev. Ceres 61(2):178-183.
Crossref
|
|
|
|
|
Viana AES, Sediyama T, Lopes SC, Cecon R, Silva AA (2001). Efeito do comportamento e de incisões no córtex da mandioca sobre o cultivo da mandioca (Manihot esculenta Crantz). Acta Sci. 23(5):1263-1269.
|
|
|
|
|
Vidigal-Filho OS, Pequeno MG, Scapim CA, Vidigal MCG, Maia RR, Sagrilo E, Simon GA, Lima RS (2000). Avaliação de cultivares de mandioca na região noroeste do Paraná. Bragantia 59(1):69-75.
Crossref
|
|
|
|
|
Voll E, Gazziero DLP, Brighenti AM, Adegas FS, Gaudêncio CA, Voll CE (2005). A dinâmica das plantas daninhas e práticas de manejo. Londrina: Embrapa Soja. (Documentos, 260) P 85.
|
|