Full Length Research Paper

ABSTRACT
Experiments were conducted with arsenic-hyperaccumulator, Pteris vittata, and non-hyperaccumulator, Pteris ensiformis, subjected to various arsenic toxicity levels (0 to 1500 mg As kg-1) in peat-moss or soil for up to 14 weeks, to evaluate phenotypic responses and total lipid with fatty acid profiles, and P. vittata influence on rhizosphere enzyme activities. P. ensiformis exhibited significant phenotypic toxicity response compared to P. vittata. P. vittata sequestered significantly (p<0.5) higher arsenic (5,160.6 mg As kg-1) than P. ensiformis (313.4 mg As kg-1). Cellular damage and physiological death occurred in P. ensiformis with alteration in fatty acids and lipid compositions but no significant changes in P. vittata. Both showed no detectable C16:3 but a decrease in C18:3, which was more pronounced in P. ensiformis (51.2%); hence, are reported as C18:3 plants for the first time. Acid phosphatase, β-glucosidase, alrysulphatase, and N-acetyl-β-D-glucosaminidase activities were inhibited by As and significantly (p<0.5) negatively correlated to As concentration. Enzyme inhibition range from 6% in β-glucosidase to 67% in N-acetyl-β-D-glucosaminidase. These results revealed P. vittata phenotypic tolerance to As stress was mediated through metabolic-readjustment, especially its fatty acid and lipid compositions. Additionally, P. vittata rhizospheric events modulate soil enzyme activities in the As-contaminated soil.
Key words: Fern, heavy metal, phenotypic response, fatty acids, acid phosphatase, β-glucosidase, alrysulphatase, N-acetyl-β-D-glucosaminidase, rhizosphere.
Abbreviation: BF, Bioaccumulation factor; DBI, Double bond index; FAMEs, Fatty acids methyl esters; GLU, β-glucosidase; NAG, N-acetyl-β-D-glucosaminidase; PHOS6, Acid phosphatase; ROS, Reactive oxygen species; RZS, Rhizosphere soil; SUL, Arylsulfatase; TF, Translocation factor; THAM, Tris-hydroxymethyl aminomethane.INTRODUCTION
Heavy metals and metalloids contamination have received considerable attention in the past decades due to increasing environmental contamination from anthropogenic activities: industrial and municipal waste, as well as agrochemicals (Khan et al., 2017). Some heavy metals and metalloids (Cu, Fe, Zn, Se, Si, Ni, Mo, Co) are essential elements required by plants in traceamounts for cellular metabolic processes but are toxic at high concentrations. On the other hand, non-essential metals or metalloids (As, Al, Cr, Pb, Cd, Hg) are toxic for plants even in trace amounts. Under heavy metals and metalloids stress, plant growth and performance are impacted, triggering complex adaptive responses such as membrane lipid and fatty acid modifications, alteration of root architecture, and plant physiological processes at the root-soil interphase that can influence major enzymes that are involved in nutrient cycling and availability. Assessing these responses may provide better understanding of tolerance and nutrient management for plants in heavy metal-contaminated soil and provide insights into use of adapted plants for bioremediation.
Plants, as sessile organisms, are generally exposed to a plethora of abiotic (drought, salinity, nutrient deficiency, heavy metals and metalloids contamination, etc.) and biotic stresses in their environment, which influence their growth and development, survival, and productivity (Upchurch, 2008; Ramakrishna, 2014; Bhalerao et al., 2015; Goyal et al., 2020). Plants, thus, have evolved varied mechanisms of agronomic and biochemical adaptations under extreme ecological and stress conditions. Common phenotypic responses to heavy metals and metalloids toxicity in plants include growth retardation, leaf chlorosis and necrosis, impaired photosynthesis, respiration, mineral nutrition uptake, gene expression, initiation of senescence and program cell death (Caille et al., 2004; Shaibur et al., 2008; Steiner et al., 2012). Some of these symptoms are similar to those observed in nutrient deficient plants. For example, aluminum (Al) toxicity phenotype is similar to phosphorus deficiency phenotype, which is purple discoloration on stems and leaves, followed by leaf veins yellowing and dead leaf tips. Aluminum toxicity may mimic P deficiency phenotype and calcium deficiency mimicking drought or root damage stress (Bojórquez-Quintal et al., 2017).
At the cellular levels, abiotic stress such as heavy metals and metalloids exposure elicits metabolic disruption and cell membranes are generally the first targets of such perturbations. Therefore, responses mediated through the membrane structure usually involve induced alteration in lipids concentration and membrane fatty acids composition (Peixoto et al., 2001; Farid et al., 2013). This change in fatty acids composition may result in remodeling of the plasma membrane fluidity leading to increased membrane permeability (Cumming and Taylor, 1990; Kaba?a et al., 2008), especially to reactive oxygen species (ROS) molecules, and other molecules.
In heavy metal contaminated soil, through association with rhizospheric microbes and arbuscular mycorrhizal, plants can effect changes in metal speciation, toxicity, mobility, extracellular enzyme activities and nutrient mineralization, solubilization, and bioavailability (Gadd, 2010; Kim et al., 2010; Shahid et al., 2014). Furthermore, these microbes play a critical role in plant nutrients cycling, degradation of xenobiotics, plant pathogens control, maintenance of soil structure, and plant growth promotion (Hassan et al., 2019). In agricultural soil, the rhizosphere, a region of soil immediately surrounding plant roots and nutrient-rich, is the main site of these biochemical activities, nutrients mineralization, and the complex ecological interactions between plant, soil, and microorganisms. Extracellular enzymes produced and released here mediate reactions of biogeochemical importance especially nutrients release for plant growth. Some key enzymes of agronomic importance include arylsulfatase (SUL) that catalyzes the hydrolysis of organic sulfur with aromatic radical (Tabatabai and Bremner, 1970); β-glucosidase (GLU) involved in carbon cycling and energy release for microbial growth (Deng and Popova, 2011) acid phosphatase (PHOS6), predominant in acid soils, catalyzes the hydrolysis of phosphomonoesters to release inorganic phosphorus (Eivazi and Tabatabai, 1977); and N-acetyl-β-D-glucosaminidase (NAG) critical to N mineralization in soils (Ekenler and Tabatabai, 2002). Soil enzyme activities are measured as a good indicator of soil health because of their sensitivity to ecological disturbance which is easily detected in fast changes in the activities associated with them such as mineralization, soil physical properties, microbial growth, etc. (Alkorta et al., 2003); Dick and Kandeler, 2005). They can therefore serve as good predictors of effects of perturbations on soil quality and availability of mineralized nutrients and help in determining the state of health and performance of the plants, especially in contaminated environment. Studies have shown that these activities may be inhibited by environmental contaminants, especially heavy metals and metalloids such as arsenic (As); hence, affecting plants access to nutrients needed for growth and development. Plants in response may alter the soil chemistry through rhizosphere activities such as root exudations that form organic complexes with nutrients, chelate heavy metals and metalloids and may enhance nutrient access and uptake (Nguyen, 2003). Although there are several literature on the impact of heavy metals and metalloids on soil enzymes (Tabatabai, 1977; Karaca et al., 2010; Angelovi?ová et al., 2014), little is however known about extracellular enzyme activities in the root zone of hyperaccumulators in contaminated soil and the influence of the plant rhizodeposition. Plant produce higher amount of root exudates in response to stress such as heavy metals (Wenzel et al., 2003; Fu et al.,2017; Luo et al., 2017).Organic compounds contained in these exudates have been shown to cause changes in soil redox potential, reducing soil pH, metals mobilization by forming soluble chelate complexes with heavy metals (Montiel-Rozas et al., 2016) thus influencing both plant roots interact with different microorganisms and rhizosphere enzymes activity and metal uptake by the root. Heavy metals and metalloids are also known to depress soil enzyme activities (Wang et al., 2007). In a time-series experiment, Gao et al. (2010) reported an increase in inhibition of phosphatase, urease and dehydrogenase activity with increasing Cd, Cu, Zn, Pb and As concentrations over time.
Some plants are however, tolerant to heavy metal and metalloid stresses. These plants have evolved mechanisms to withstand heavy metals and metalloids toxicity, which mostly involve avoiding metal uptake through metal-cell wall binding, active metal ions efflux, metal immobilization through chelation by organic acids produced by the root, mycorrhization, etc. Once in the plant, however, heavy metals and metalloids toxicity is reduced by the production of antioxidant enzymes, production of ROS as signaling molecules, and metal chelating followed by sequestration of the metals in vacuoles. Under normal conditions, ROS production and destruction are controlled through cellular homeostasis (Raja et al., 2017) and involves signaling molecules derived from fatty acids (De Carvalho and Caramujo, 2018). However, during stress such as exposure to heavy metals and metalloids, oxidative burst may occur in which the production of ROS supersedes the scavenging ability of the cell. The ROS are cytotoxic, and their cellular effects include lipid peroxidation, proteins denaturation and nucleotide degradation (Raja et al., 2017) disrupting cellular functions of fatty acids, among others. Fatty acids are structural components of the cell membrane and precursors of signaling molecules (e.g. jasmonates) involved in response to ROS production. Linolenic acid (C18:3 ), for example, is involved in stress response and is released from the membrane lipids as precursor of jasmonates (JAs) and phyto-oxylipin biosynthesis in response to abiotic (e.g. freeze-thawing) or biotic (insects or pathogens) stresses (Blée, 2002; Upchurch, 2008); oleic acid (C18:1) is involved in pathogen defense, for instance in Arabidopsis sp. (Kachroo et al., 2001); oleic (C18:1) and linoleic (C18:2) acids modulate the growth, development, and mycotoxin production in Aspergillus spp. (Wilson et al., 2004). Investigations have shown compositional change in fatty acids of different plant species following heavy metal stress. In tomato plant, Verdoni et al. (2001) reported overall changes in leaf fatty acids composition with a decrease in the percentage of tri-unsaturated (mainly C18:3) fatty acids in the leaves and increased production of C18:2, C18:0 and C16:0) in plant exposed to cadmium stress. Comparing the cadmium (Cd) stress in Brassica napus (rapeseed), a non-hyperaccumulator and a well-known Cd-hyperaccumulator plant, Brassica juncea (Indian mustard), Nouairi et al. (2006) observed a 52% decrease in leaf total lipids in B. napus whereas B. juncea had a 19% compared to the control plant. Nouairi et al. (2006) also reported a decline in polyunsaturated fatty acids (mainly linolenic acid [C18:3] and hexadecatrienoic acid [C16:3]), and increase in the level of saturated fatty acids (C18:0 and C16:0) with B. juncea (the Cd-hyperaccumulator) showing more stability in the cellular membrane compared to B. napus (a non- hyperaccumulator of Cd).
Plants that can tolerate and hyperaccumulate heavy metals and metalloids have been suggested as good candidates for remediation of heavy metals and metalloids polluted soils. Such plants can prevent ROS damage and hyperaccumulate heavy metals and metalloids from the contaminated environment in their tissues. Many plant species, including the ferns (Pteris spp.), Brassica juncea, Salix spp., (willow), Populus sp. (poplar), etc. are examples of tolerant and hyperaccumulators of heavy metals and metalloids (Nouairi et al., 2006; Liu et al., 2018). The knowledge of the mechanisms that differentiate tolerant species from non-tolerant ones is still limited. Pteris vittata is the first identified hyperaccumulator of arsenic (As), a ubiquitous metalloid known to be carcinogenic (Ma et al., 2001). P. vittata can tolerate and sequester exceptionally high amounts of As in its aboveground tissue, making it a suitable candidate for remediation of As contaminated environment. Although the mechanism of As uptake as well as antioxidant activities have been described in P. vittata, changes in fatty acid profiles due to As exposure in soil remains unknown, and how this may differ in its close relative, non-arsenic tolerant, and non-hyperaccumulators like P. ensiformis (slender brake) is yet to be explored. Furthermore, information on the influence of As-challenged P. vittata on soil enzyme activity is limited. Knowledge of this is important for understanding of tolerance of plants to heavy metals and metalloid stress and may provide information developing strategies for enhancing efficiency of hyperaccumulator for more cost-effective remediation. The objectives of this study were to: i) screen phenotypic response of P. vittata and P. ensiformis to As toxicity, ii) evaluate the effects of As on the ferns total lipids and fatty acids composition, and iii) assess the influence of P. vittata on soil phosphatase, β-glucosidase, alrysulphatase, N-acetyl-β-D-glucosaminidase activities in As-contaminated soil.
MATERIALS AND METHODS
Plant and soil materials
Two fern species, the arsenic hyperaccumulator and tolerant P. vittata (Chinese brake fern) and the non-hyperaccumulator and non-tolerant Pteris ensiformis (slender brake fern) seedlings were obtained from ARC Ferns LLC (Apopka, FL) at five-leaf stage. The plants were transplanted into peat moss potting material for acclimatization for four weeks.
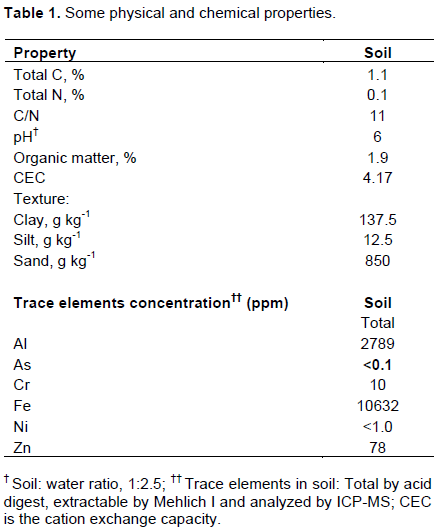
Soil was collected from Tuskegee University’s George Washington Carver Agricultural Experiment Station located in Tuskegee, Alabama (32° 26’ 15.59” N 85° 44’ 0.8.70’ W). The soil is a Marvyn soil with taxonomic classification fine-loamy, kaolinitic, thermic Typic Kanhapludults (sandy loam at 0-15 cm). The primary land use of the area is foraging and has not received any soil amendment in the 5 years prior to soil collection. The top (0-15 cm) soil was collected, sieved to pass through a 2 mm mesh sieve to obtain a homogenous soil sample then air-dried. The soil baseline physiochemical parameters (total C, total N, pH, organic matter, CEC, texture, and elemental concentrations) were determined at the Soil, Forage and Water Testing Lab at Auburn University, Auburn, AL (Table 1).
Experimental designs
Two subsequent experiments were set up in a complete randomized design with a factorial arrangement and each treatment replicated three times. The As treatment included control (no As) and increasing rate of As (As2O3, 99.99% trace metal basis; Sigma-Aldrich Chemical Company, St. Louis, MO, product number 202673) spiked in two planting medium types. The peat moss treatments (0, 50, 250, and 500 mg As kg-1) were done under laboratory conditions by placing the peat moss-potted plants directly in an appropriate volume of As containing Hoagland solution (25%) needed to attain 60% water holding capacity, such that the peat moss imbibed the solution completely. Also, As-spiked soil mixtures (0, 50, 100, 150, 200, 500, 1000, and 1500 mg As kg-1 soil) were cured first under fume hood for 12 weeks before ferns transplantation. Transplanted ferns were then transferred to the greenhouse and maintained under natural photoperiod, ambient temperature and humidity conditions.
Phenotypic screening (visual observation) in Arsenic contamination soil and peat moss
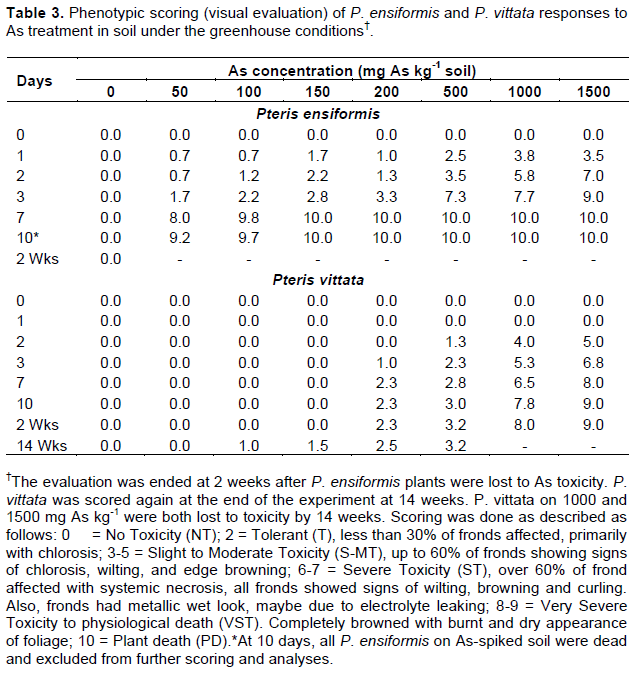
The four-weeks-old acclimatized ferns were subjected to treatments either. As-spiked peat moss or As-spiked soil for up to 10 days or 14 weeks, respectively. Phenotypic evaluations of plants under As toxicity were performed at days 0, 5, and 10 for plants in As-amended peat moss in a lab setting, and days 0, 1, 2, 3, 7, and 10, and week 14 in As-spiked soil in the greenhouse. The phenotypic responses were assessed by visual observation of the plants for signs of As stress such as chlorosis, necrosis, wilting (decrease water potential), etc. and scored using a rubric of 0-10 (where 0 = tolerant or no toxicity effect; 1-2 = tolerant/no toxicity (nt), less than 30% of fronds affected, primarily with chlorosis; 3-5 = slight to moderate toxicity (s-mt), up to 60% of fronds showing signs of chlorosis, wilting, and edge browning; 6-7 = severe toxicity (st), over 60% of frond affected with systemic necrosis, all fronds showed signs of wilting, browning and curling, fronds with a metallic wet look, 8-9 = very severe toxicity to physiological death (vst), completely browned with burnt and dry appearance of foliage; 10 = plant death).
Sample preparation for total elemental, lipid and fatty acid composition profiles
Each treated and control plant was carefully removed from the pot on day 10 (in peat moss) and week 14 (in soil), the roots (and rhizomes) were gently shaken to remove adhering peat moss or soil and rinsed thoroughly with deionized water. The plant was separated into aboveground (fronds/leaves) and belowground (roots and rhizomes) tissues then air-dried for 72 h followed by oven drying at 60°C for 48 h. The samples were ground to pass 0.5 mm mesh and store at 4°C until utilized for total elemental and fatty acids methyl esters (FAMEs) analyses. Total lipids extraction and FAMEs analyses, with C19:0 (Nu-Chek Prep Inc., Elysian, Minnesota) as internal standard, were performed according to Folch et al. (1957) and modified according to Abugri et al. (2013). FAMEs evaluation was performed using Gas Chromatography-Mass Spectrometry (GC-MS) using a 1 µl spitless injection on an Agilent 6890 GC with a 5975 MS equipped with a Restek Stabilwax-DA column 30 m × 0.25 mm i.d and 0.25 µm film thickness. A GLC 463 (Nu-Chek Prep Inc., Elysian, Minnesota) fatty acids standards mix was utilized as the methyl ester reference standards. Fatty acid desaturation was measures as double bond index (DBI), calculated as: DBI = {[C16:1] + [C18:1] + 2 × [C18:2] + 3 × [C18:3]} (Liu and Huang, 2004).
Total elemental analysis in plant material
The air-dried and ground plant tissues from above were microwave digested using HNO3 extraction techniques (USEPA method 3051a; USEPA, 2007). Total elemental concentrations of the filtered and diluted digests were analysed by inductively coupled plasma mass spectrometry (ICP-MS) at the Laboratory for Environmental Analysis at the Centre for Applied Isotope Studies at the University of Georgia, Athens, GA.
Soil enzyme analysis
Soil samples were collected from rhizosphere soil (RZS) of soil-potted P. vittata, 14 weeks post-treatment, and assayed for arylsulfatase (SUL), β-glucosidase (GLU), acid phosphatase (PHOS6), and N-acetyl-β-D-glucosaminidase (NAG) activities. The soil enzyme assays were performed using the microplate fluorometric method as described by Deng et al. (2011) with some modifications for high throughput and a simultaneous assay of multiple enzymes in multiple soil samples. The modification included: increase in the assay volume (2 ml 96-deep-well plate utilized instead of the 350 µml microplates) to improve sample representation and assay reproducibility (samples aliquots were later transferred into corresponding well in a black 350 µl microplate for reading in the fluorometer after termination of the reaction); a blender (Waring Commercial, Torrington, CT) was utilized to homogenize the sample at maximum speed for 1 min before transferring to a beaker for continuous stirring over a magnetic stirrer (Bell et al., 2013; German et al., 2011); use of a “standard buffer pH 6” all the enzymes calibration curve [test showed no significant difference among the buffer pHs when used for calibration curve (data not shown)]; THAM buffer (pH 12) was used for assay reactions instead of NaOH, and the plate layout was reformatted for a high throughput assay. Final enzyme activity was calculated and expressed in µmol h-1 g-1 soil.
Statistical analysis
The data presented are means of three replicates and two experiments. The peat moss experiment was set up as a complete randomized design (2×4×3) with factorial arrangement: 2 plants species × 4 As concentrations, and 3 replications. In the greenhouse study, the experimental design was set up as 2x8x3 completely randomized factorial design: 2 plants species x 8 As concentrations, and 3 replications. However, due to As toxicity to P. ensiformis, only P. vittata was maintained up to 14 weeks in the greenhouse experiment, therefore soil enzyme data are only presented for P. vittata. Fatty acid analysis was done on As treatments 0 to 150 mg As kg-1 peat moss for both P. ensiformis and P. vittata, and treatments 0 to 500 mg As kg-1 soil in greenhouse study only for P. vittata. Statistical analysis was performed using SAS, XLSTAT, or Analyze-It software. Analysis of variance according to the General Linear Model of SAS was utilized to test for treatments effects and means were separated by Tukey’s multiple comparison test using p<0.05 level of significance.
RESULTS AND DISCUSSION
This study concomitantly evaluated several characteristics (phenotype observations, lipid profiles, soil hydrolytic enzyme activities) in P. vittata and P. ensiformis during growth and development in As-spiked peat moss and soil at different concentrations. Both Pteris spp. responded differentially to As concentration as indicated by the phenotypic response and fatty acids changes, as well as inhibition of major soil enzymes described in this study.
Phenotypic response of P. ensiformis and P. vittata to Arsenic toxicity
Both Pteris species in As-spiked peat moss and the As-spiked soil showed phenotypical responses to As toxicity during the 14 week-experiment. The visual severity of toxicity effect increased with increase in As treatment and duration of treatment. Phenotypically, As toxicity response was more severe in P. ensiformis than P. vittata in both As-spiked peat moss (Table 2) and soil (Table 3). In the As-spiked peat moss, P. ensiformis showed severe phenotypic responses (wilting, lodging, and frond rolling) within 24 h of As treatments. This was followed by progressive wilting, with significant foliar chlorosis, general browning, necrotic lesions, as well as frond curling and lodging. At 50 mg As kg-1, moderate frond chlorosis, wilting, and frond edge browning was observed in up to 40% of the plant at 5-day post-treatment and this increased to 60% at 10 days post-treatment compared to 0% for P. vittata. Moreover, at 5-days post-treatment in 500 mg As kg-1 peat moss, over 70% of P. ensiformis fronds showed browning, wilting and curling, as well as frond edge necrosis. The fronds also exhibited a metallic wet look, suggesting electrolyte leaking, and there was physiological death at 10-days post-treatment (Figure 1). P. vittata, on the other hand, only showed signs of frond edge and tip necrosis on older fronds and slight chlorosis in less than 30% of its fronds 10-days post-treatment in 500 mg As kg-1 peat moss but no observable toxicity response in the 50 mg As treatment.


Similar severe phenotypic response to heavy metal toxicity has been reported in physic nut (Jatropha curcas L.) exposed to Al for 75 days and was described as having chlorotic leaves with small necrotic spots on the edge and also winding in the young leaves while the older leaves had marginal chlorosis that progressed to the center of the frond (Steiner et al., 2012). Leaf rolling and small necrotic spotting have been reported in other heavy metal toxicity, including nickel (Ni) (Rahman et al., 2005) and Zn (Sagardoy et al., 2009). Symptoms recorded in this study were similar to that reported for fenugreek (Trigonella foenum-graecum L. cv. Azad) exposed to As treatment (Talukdar, 2013) which included biomass reduction, (wilting), chlorosis, reduction of photosynthetic activity, lipid peroxidation, low yield, and accelerated ageing. This was also observed for beans (Phaseolus vulgaris L.) (Stoeva et al., 2005), tomato (Solanum lycopersicum) (Miteva, 2002), rice (Oryza sativa) (Rahman et al., 2008) and in sorghum (Sorghum bicolor) (Shaibur et al., 2008).
Greenhouse experiment for both P. vittata and P. ensiformis in As-spiked soil showed signs of chlorosis and browning homogeneously over the fronds, and frond necrosis at very high As concentrations (1,000 and 1,500 mg As kg-1) after 48 h of treatment. These effects progressed rapidly in P. ensiformis at all As treatment levels, resulting in physiological death at 10-days post-treatment. The symptoms were similar to those described above for As-spiked peat moss but more severe. In P. vittata, there was initial uniform chlorosis in old fronds, followed by frond edges and tips browning and necrosis at 500 mg As kg-1 and above. These symptoms stabilized at 10-days post-treatment in P. vittata exposed to 500 mg As kg-1 soil, but symptoms progression continued in the 1,000 and 1,500 mg As kg-1 soil. After 24 days, there was 100% leave browning with a metallic hue. New fronds emerged from the rhizome in these treatments (1,000 and 1,500 mg As kg-1 soil) after 48 h, but these fronds quickly developed chlorosis, browning, wilting, edge rolling and senescence. Singh et al., (2006) also reported chlorosis and necrosis in P. ensiformis after As treatment (in hydroponic system) compared to no effect in P. vittata at the same treatment levels (0, 133, 267 µM As, which are equivalent to 0, 10, and 20 mg As kg-1 in this study). The response in P. vittata at the very high As concentrations (1000 and 1500 mg kg-1 soil) observed in this study was equally reported by Caille et al. (2004) in P. vittata grown on soil with an average As concentration of 4550 mg As kg-1. They reported chlorosis and necrosis in the fronds of the fern after 198 days and were not able to collect any root sample for analysis. Similarly, fresh plant tissues harvested from very high As treatments in this study were insufficient for the necessary downstream analyses. Only 0-150 mg As treated P. vittata harvested at 14 weeks post treatment were therefore utilized for further analyses. Moreover, all As treated P. ensiformis died within 2 weeks of As treatment, therefore, excluded.
The phenotypic responses of both Pteris spp. to As toxicity reported in this study are similar to responses described in other plants under heavy metal toxicity stress (Rahman et al., 2005; Steiner et al., 2012; Talukdar, 2013). The phenotypic observations resemble symptoms of other abiotic stresses-nutrient deficiency. The similarity in symptoms suggests disruption in the cell membrane, primary site of nutrient transport. This disruption may deplete or remodel cell membrane lipid and fatty acid concentrations and composition resulting in the phenotypic changes observed (chlorosis, wilting, etc.).The chloroplasts are plants sites of light-harvesting for sugar production-photosynthesis. It is also an important site of ROS production (as is mitochondria and the peroxisomes). The fronds chlorosis may be due to inhibition of photosynthetic enzymes and chloroplast membrane lipid/fatty acids remodeling leading to membrane permeability to more radicals or more seriously, gene expression reprogramming. These alterations may induce a response from other thylakoid membrane proteins resulting in impaired/inhibition of photosynthetic pigments production (Kleine and Leister, 2016; Paunov et al., 2018).
A direct effect of As that may explain the observed necrosis and browning is the potential participation of As in biological reactions replacing P and imparting the cellular energy currency, ATP, producing ADP-AsV instead thus uncoupling oxidative phosphorylation. The toxicity at high As treatment (500 mg As kg-1 in peat moss and above 1000 mg As in soil) as observed in phenotype changes in P. vittata, a known As-tolerant and hyperaccumulator, suggest a possible As toxicity tolerance threshold for the fern. Ma et al. (2001) reported P. vittata tolerant to soil spiked with 1,500 ppm As and accumulating 15,861 ppm As in 2 weeks. In this study, while the P. vittata was able to tolerate the As treatment, plants in soil spiked with greater than 500 mg As showed toxicity visual symptoms which resulted in the loss of biomass and growth retardation. The difference in threshold may be due to the fern ecotypes, P. vittata obtained from around a contaminated site (up to 1,603 mg kg-1 As) were used in Ma et al. (2001) study, while in this study plants were obtained from a nursery. As-spiked peat moss material was more potent than the As-spiked soil; due to higher bioavailability of As in the peat moss compared to the soil. However, P. vittata phytoextraction and sequestration of As was at a magnitude many orders higher than in P. ensiformis which died after few days due to high As toxicity.
Arsenic and total elemental concentrations of P. ensiformis and P. vittata
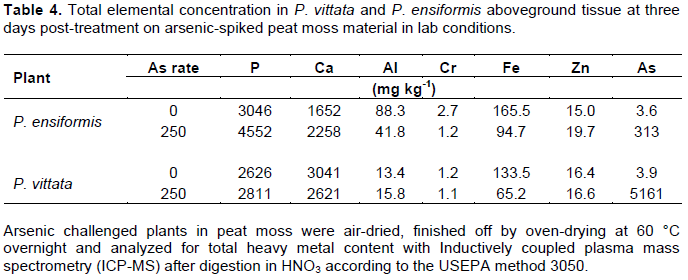
Arsenic accumulation was several order of magnitudes higher in P. vittata fronds compared to P. ensiformis in similar As-spiked peat moss at 3-days post-treatment (Table 4). However, P. ensiformis had higher P concentration compared to P. vittata. In both ferns, As treatment enhanced P uptake with a 49.4% increase in P. ensifomis and only 7.0% in P. vittata compared to their respective controls. These observations are contrary to those reported by Huang et al. (2006) in P. vittata grown on acid-washed quartz sand and supplemented with half-strength Hoagland nutrient. However, in other studies (Liao et al., 2004; Lessl et al., 2015) in which P. vittata was grown on As-contaminated soil, results obtained in present study are consistent with their observations. The results showed positive correlations between plant tissue As and P concentrations, as well as between soil As treatments and plant tissue As concentrations.
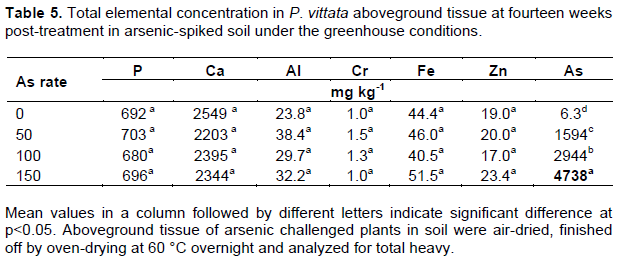
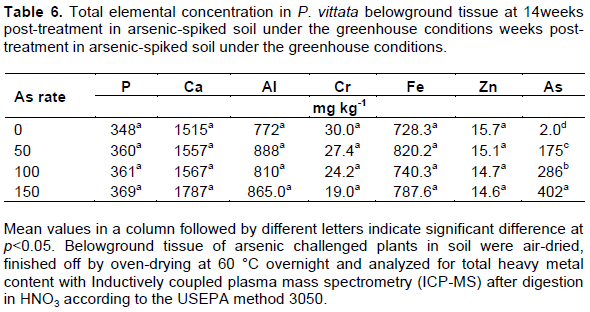
Analysis of root and fronds elemental concentrations in P. vittata in the greenhouse study revealed a similar trend observed in the As-spiked peat moss study. The frond and root As concentrations increased with increasing soil As concentration, with a 10-fold higher As in the fronds compared to the roots (Table 5). The bioaccumulation factor (BF), a ratio of plant tissue As to soil As averaged 30.9 in the fronds but just 3.0 in the root. The translocation factor (TF), ratio of frond As to root As average was 10.4 for P. vittata grown on As-spiked soil. These values are similar to those reported by Lessl et al. (2015) who observed a BF of 26 for frond and 3.5 for root, and TF of 7.8 for P. vittata cultivated on As-contaminated soil (0.83 to 96.7 mg As kg-1 soil). However, they reported a much higher BF and TF for naturally growing P. vittata in the same soil (BF: of 25 for root, 232 for fronds, and TF: of 17). Phosphorus, an analog of As, showed only slightly increase at 50 mg As kg-1 soil (1.6%) compared to the control, however there was no significant difference among treatments (p<0.5). The P content was otherwise stable (692.8±9.7 mg As kg-1 in the fronds; 359.5±8.9 mg As kg-1 in the roots) compared to As (Tables 4 to 6). There was a strong correlation between the soil As and plant tissue As in both root and frond (r2 = 0.97 and 0.99 respectively).
Effect of As stress on lipid and fatty acids concentrations
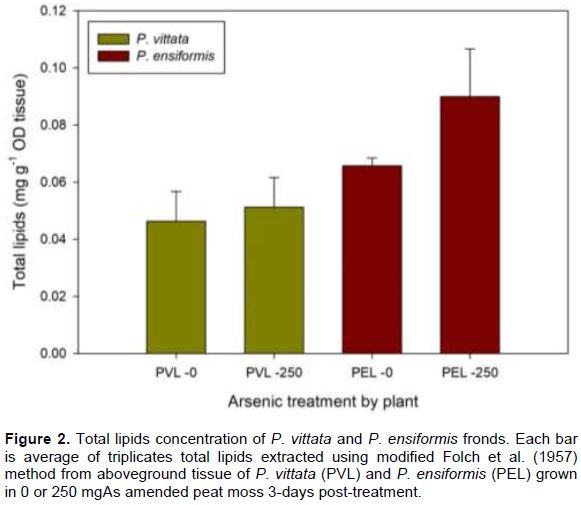
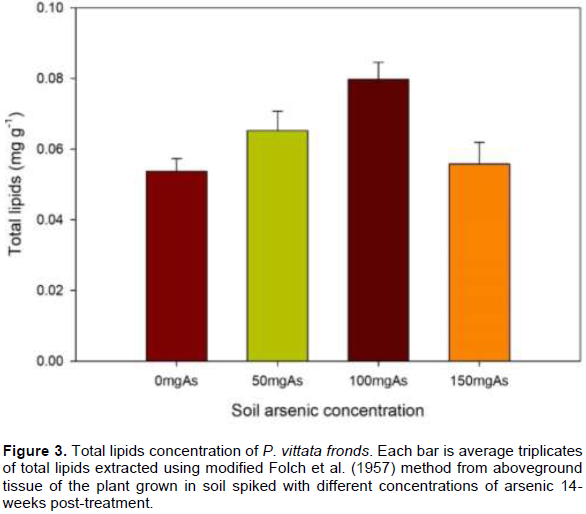
The frond is the primary site of As sequestration in these ferns, the effect of As on lipid and fatty acid compositions in the frond was therefore evaluated in this study. The total lipid composition of P. vittata and P. ensiformis challenged on A-spiked peat moss is shown in Figure 2. There was an overall increase in total lipids concentration at higher As concentration in both fern species, on the average, 36.8% more in P. ensiformis and 10.4% in P. vittata compared to their respective controls. P. ensiformis generally has more total lipids concentration than P. vittata both in control and As-spiked peat moss (Figure 2). P. vittata in As-spiked soil also showed higher total lipids concentration compared to the control, 21.4, 48.4, and 3.9% in 50 mg As, 100 mg As and 150 mg As Respectively for 14 weeks, showed a decrease in total frond lipid concentration (Figure 3). This result is contrary to data reported by Nouairi et al. (2006) in comparing a Cd hyperaccumulating Brassica juncea and non-hyperaccumulating Brassica napus. They reported a decrease in total lipid concentrations in the non-hyperaccumulator but increase in the lipid concentration of the hyperaccumulator with increase in Cd treatment similar to data reported in this study.
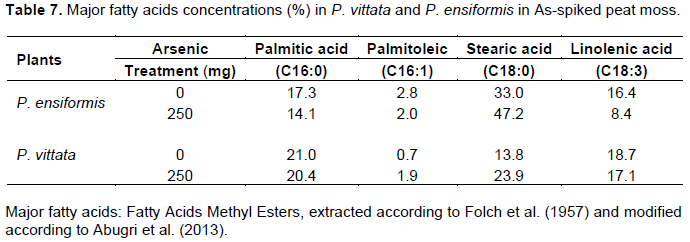
Tables 7 and 8 show the changes in major fatty acids in the and greenhouse experiment respectively. There was no detectable cis-7,10,13-hexadecatrienoic acid (C16;3) in both fern species in peat moss and in soil treated with or without As. The polyunsaturated fatty acid, α-linolenic acid (C18:3) decrease with increase in As treatment in P. ensiformis and P. vittata in As-spiked peat moss compared to the control (Table 7). Similarly, C18:3 decreased with increase in As treatment in As-spiked soil (Table 8). The saturated fatty acid, stearic acid (C18:0) on the other hand increase with increase in As treatment. Overall, fatty acid unsaturation expressed as double bond index (DBI) a decline with increase in As treatment. There is, therefore, a high level of protection against lipid peroxidation in P. vittata compared to P. ensiformis. Data showed prevalence of C18:3 and the absence of detectable C16:3 in both P. vittata and P. ensiformis fronds demonstrating for the first time that they are both C18:3 plants (Browse and Somerville, 1991; Mongrand et al., 1998). Linolenic acid (C18:3) is a precursor for the biosynthesis of jasmonic acid, a defensive mechanism activator in plant (Weber, 2002) in response to stress. In P. ensiformis, there was over 49% reduction in C18:3 fatty acid in favor of stearic acid (C18:0), as a result of lipid peroxidation, decreasing the polyunsaturated fatty acid content of the fronds. The consequence of this is membrane damage and loss of integrity, which was evident by the phenotypic observation (wet look due to possible cellular leakage). P. vittata, on the other hand, showed a marginal change in the C18:3 fatty acid in response to As toxicity ensuring membrane integrity and tolerance to As toxicity.

Although there was no known literature on the fatty acid composition of P. vittata to compare this study’s fatty acids data to, lipid peroxidation, which results in fatty acid compositional changes have been reported in plants under stress (Srivastava et al., 2005; Watanabe et al., 2014; Casado et al., 2015). The reported increase in lipid peroxidation with the increase in As stress is consistent with the increase in overall fatty acid saturation (with increase in As concentration) that is reported here. The differences observed in the level of severity of the As treatments between P. vittata and P. ensiformis confirmed that the degree of As toxicity is dependent on the plant species, exposure time, source of contamination, media, bioavailable As, etc. as suggested by Pourrut et al. (2011).
Soil enzyme activity
Arsenic treatments significantly (p<0.0001) affected all the rhizosphere enzyme (arylsulfatase, β-glucosidase, acid phosphatase, and N-acetyl-β-D-glucosaminidase) activities studied. Enzyme activities ranged from 6.2% in GLU in 50 mg As treatment to 69.8% in NAG at 500 mg As treatment. In general, enzyme activities significantly decreased in soil treated with higher than 50 mg As and there was no significant difference in the inhibition within each enzyme in 100 mg As to 500 mg As soil. However there was general trend of decrease in all enzyme activities with increase in As concentration except in acid phosphatase (Figure 4). The highest average activities were recorded for acid phosphatase (up to 1.42 µmol g soil-1 h-1 in 500 mg As) and the lowest in SUL (0.12 µmol g soil-1 h-1 in 150 mg As). Juma and Tabatabai (1977) identified As among the most effective inhibitors of phosphatase with > 50% average inhibition in three soils treated with 25 µM trace element g-1 soil, and Karaca et al. (2002) reported inhibition of GLU activity by Cd in soil amended with Cd and sewage. Das et al. (2017) similarly reported a 2.5% decrease in GLU activity in P. vittata rhizosphere spiked with 100 mg As compared to control In a time-series experiment, Pan and Yu (2011) reported inhibition of 21.3 to 30.0% in acid phosphatase in soil treated with a combination of Cd and Pb after two weeks. Acid phosphatase activity inhibition by other heavy metals has been reported in other studies such as by antimony (An and Kim, 2009), and Cd (Karaca et al., 2002). In soil, As toxicity is known to inhibit soil enzymes and disrupt plant growth-promoting bacteria especially those involved in nutrient cycling (e.g. nitrogen fixation), and nutrient supply to plant (Porter and Sheridan, 1981).
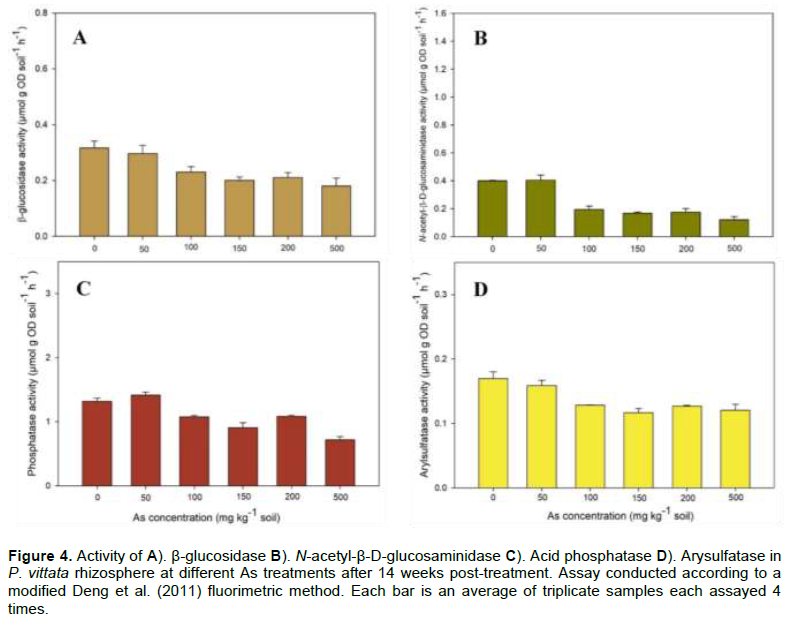
In this study, all the P. ensiformis grown on the As-spiked soil were all lost after 10 days, due to a combination of aboveground ROS induce events and belowground root damage. In a study by Pajuelo et al. (2008), alfalfa grown on As-contaminated soil were lost as a result of root necrosis, root hair damage, and infection susceptibility. They postulated that the loss of the root prevented the plant access to nutrient needed for cellular activity and maintenance. In this study, there was evidence of root necrosis in P. ensiformis (data not shown) with no sign of new root growth upon examination of the dead plants. The fronds also had a metallic wet look, as a result of leakage of cellular contents due to possible membrane disruption and damage.
Enzyme activity results from this study is consistent with observations from other studies such as Lorenz et al. (2006) who reported inhibition of alkaline phosphatase, arylsulfatase, protease, and invertase in soil contaminated with As and Cd. Investigating the effect of As fractions on β-glucosidase, urease, acid/alkaline phosphatase, and arylsulfatase activities, Bhttacharryya et al. (2008) reported significant differences in the enzyme activities in the studied soil, but, all the enzyme activities were inhibited compared to in the control soils. They demonstrated a strong negative correlation between the enzyme activities and As concentration, similar to the observed results in this study.
Among different soil enzymes, β-glucosidase, N-acetyl-β-D-glucosaminidase, acid phosphatase, and arylsulfatase are critically important in mineralization of key plant nutrients (C, N, P, and S respectively). Heavy metals and metalloids inhibit these soil enzyme activities directly by binding enzyme active site or complexing the substrate (Juma and Tabatabai, 1977), and indirectly by influencing microbial community growth, structure, and enzyme production (Lorenz et al., 2006; Gao et al., 2010) The soil surrounding plant roots presents a different interface for complex interactions between the plant, microbes, and soil with opportunity for influence by the plant on events in the rhizosphere. P. vittata is known to produce exudates high in dissolved organic carbon and phytate, especially in contaminated soil (Tu et al., 2003).
The production of a higher amount of exudate under As stress suggest changes in physiological activities in the above-ground tissue in response to the soil As concentration. The exudates provide nutrients for promotion of soil microbial growth, thus enhance extracellular enzyme production, hence higher enzyme activities in the rhizosphere and rhizoplane (Smalla et al., 2001).
Activities of GLU, NAG, SUL, and PHO6 were inhibited with increase in As concentration, but there was no significant difference in inhibition in As concentration 100 mg As and higher. This suggests influence from other factors such as the plants’ aboveground events influencing exudates production and modulation of rhizospheric events. Increase inhibition of enzyme activities with the increase in As concentration is due to enhanced bioavailability of As caused by desorption of As from As-spiked soil by phytic acid and oxalic acid found in the root exudate of P. vittata (Tu et al., 2003; Bhattacharyya et al., 2008; Fu et al., 2017). The desorbed As is therefore available to form complexes with enzymes (or their substrates) and inhibit their activities. The inhibition of enzyme activity did not seem to affect the uptake of some nutrients such a P, which was higher in plants grown in As-spiked soil compared to the control. This suggests P. vittata is capable of adequate nutrients uptake and utilization in the As contaminated soil. It has been shown that the bioavailability rather than total heavy metal or metalloid concentrations in the media determines toxicity on biological systems (Olaniran et al., 2013). Moreover, heavy metals and metalloids show different reactions and mobility between the bulk and rhizosphere soils (and rhizoplane soil) and along concentration gradients in the soil. This concentration gradients are influenced by root exudates, the composition of which depends on the plant’s biology (species, nutrient status, growth stage, etc.) and environmental factors (including soil physiochemical properties) (Gregory and Atwell, 1991). Exudates production and their effect on heavy metals and metalloids are, however, dependent on the roles of heavy metals and metalloids in the plant metabolism. For example, heavy metals such as Zn and Cu are essential for plant growth (photosynthesis and protein synthesis), while non-essential elements like Cd and As have toxic effects on plant growth (membrane damage and ROS stress) (Raja et al., 2017). Plants, therefore, may develop strategies to prevent or modulate bioavailability and uptake of these toxic non-essential heavy metals or metalloids by producing exudates which form complexes and regulate heavy metals solubility, hence, the bioavailability of the metal ions (Wenzel et al., 2003; Guo et al., 2006; Luo et al., 2017). Soil enzymes, the primary mediator of soil biochemical processes is also inhibited by heavy metal and metalloid too and influenced by rhizospheric events modulating heavy metal and metalloid bioavailability (Karaca et al., 2010).
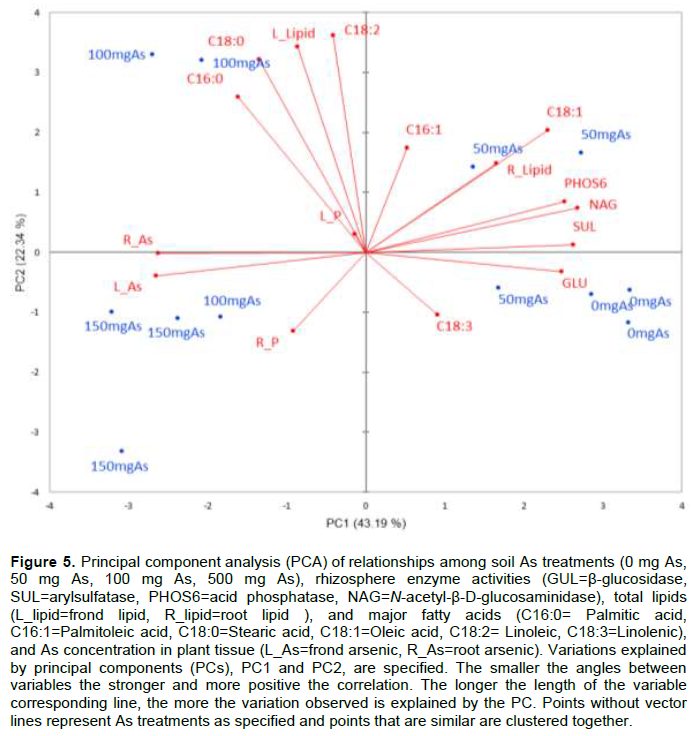
The combination of detailed physiological and biochemical results in this study with P. vittata under As stress has added to our understanding of the interrelatedness of plant response to heavy metals and metalloids. Using a principal component analysis (PCA) biplot to reduce the dimensionality of the data while retaining the variability in the data set, on Figure 5 shows the correlation between the belowground (enzyme activities, root P and As concentrations) and aboveground parameters (total lipids, fatty acids, P, and As). Total of 65.5% of the variations are explained by PC1 and PC2 (or belowground and aboveground events) ;43% of the total variations is explained by PC1 and 22% by PC2. The multivariate analysis showed that P. vittata response to As toxicity are mediated by both below- and above-ground events and are influenced by the concentration of As in the medium. The higher As treatments promoted fatty acids desaturation and higher lipid production, and positively correlated with As sequestration by P. vittata. All enzyme activities and polyunsaturated fatty acids clustered with lower As treatment while the saturated fatty acids and As are associated with higher As concentration. The strong negative correlation between total As (and P) accumulated by P. vittata and soil enzymes activity indicate an efficient nutrient uptake and utilization mechanism that may confer tolerance and ability to thrive in As-contaminated soil. The remaining, approximately 35%, variations are explained by residual PCs, suggesting more confounding factors contributing to response of P. vittata to As contamination.
CONCLUSION
This study evaluated comparatively phenotypic and biochemical responses of As non-hyperaccumulator, P. ensiformis and the As hyperaccumulator, P. vittata in As-spiked peat moss and soil. Additionally, the research focused on providing an integrative understanding of the relationship between Pteris spp. responses and its complex interactions with, and influence on rhizosphere associated hydrolytic enzyme activities in As-spiked soil. Our data showed that P. vittata critical threshold for arsenic tolerance in As-spiked soil under greenhouse conditions was 500 mg As kg-1 soil while P. ensiformis was not tolerance to all concentrations tested (50 mgAs to 1500 mg As kg-1 soil) in both peat moss and soil. Overall, fatty acid unsaturation declined with increase in As concentration. The decline was more dramatic in P. ensiformis compared to P. vittata. This difference could be related to the difference in As tolerance between the ferns and also explain the severe phenotypic responses displayed by P. ensiformis compared to P. vittata. This study demonstrated that P. vittata has minimal morphological change compared to P. ensiformis in response to As treatment. Furthermore, P. vittata, in As-spiked soil, influenced soil enzyme activities, possibly through its association with soil microorganisms, for release and uptake of needed nutrients in the As-spiked soil. The results from this study added important dimensions to our knowledge of P. vittata response to As stress and emphasizes the need for exploring integrated system approach in the study to provide better understanding of mechanism of heavy metals/metalloids tolerance and enhancing phytoremediation efficiency as well as consideration of native non-hyperaccumulator engineering for phytoremediation.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
This study was supported by funds from the USDA USDA-NIFA 1890 Grant#2014-38821-22448-iBREED to Tuskegee University Plant Biotech and Genomic Research Laboratory. The USDA McIntire-Stennis-project to Tuskegee University Soil, Environmental Laboratory and USDA-NIFA EVANS-ALLEN programs. The authors thank Dr. M. Allam Baheth of Baaheth Research and Development Laboratory, Mobile AL for his technical advice and mentorship to the first author.
REFERENCES
|
Abugri DA, Tiimob BJ, Apalangya VA, Pritchett G, McElhenney WH (2013). Bioactive and nutritive compounds in Sorghum bicolor (Guinea corn) red leaves and their health implication. Food Chemistry 138(1):718-23. |
|
|
Alkorta I, Aizpurua A, Riga P, Albizu I, Amézaga I, Garbisu C (2003). Soil enzyme activities as biological indicators of soil health. Reviews on environmental health 18(1):65-73. |
|
|
An YJ, Kim M (2009). Effect of antimony on the microbial growth and the activities of soil enzymes. Chemosphere 74(5):654-9. |
|
|
Angelovi?ová L, Lodenius M, Tulisalo E, Fazekašová D (2014). Effect of heavy metals on soil enzyme activity at different field conditions in Middle Spis mining area (Slovakia). Bulletin of Environmental Contamination and Toxicology 93(6):670-5. |
|
|
Bell CW, Fricks BE, Rocca JD, Steinweg JM, McMahon SK, Wallenstein MD (2013). High-throughput fluorometric measurement of potential soil extracellular enzyme activities. JoVE (Journal of Visualized Experiments) 81:e50961. |
|
|
Bhalerao SA, Sharma AS, Poojari AC (2015). Toxicity of nickel in plants. International Journal of Pure and Applied Bioscience 3(2):345-55. |
|
|
Bhattacharyya P, Tripathy S, Kim K, Kim SH (2008). Arsenic fractions and enzyme activities in arsenic-contaminated soils by groundwater irrigation in West Bengal. Ecotoxicology and Environmental Safety 71(1):149-56. |
|
|
Blée E (2002). Impact of phyto-oxylipins in plant defense. Trends in plant science 7(7):315-22. |
|
|
Bojórquez-Quintal E, Escalante-Magaña C, Echevarría-Machado I, Martínez-Estévez M (2017). Aluminum, a friend or foe of higher plants in acid soils. Frontiers in Plant Science 8:1767. |
|
|
Browse J, Somerville C (1991). Glycerolipid synthesis: biochemistry and regulation. Annual Review of Plant Physiology and Plant Molecular Biology 42(1):467-506. |
|
|
Caille N, Swanwick S, Zhao FJ, McGrath SP (2004). Arsenic hyperaccumulation by Pteris vittata from arsenic contaminated soils and the effect of liming and phosphate fertilization. Environmental Pollution 132(1):113-20. |
|
|
Casado NAR, Horcasitas MDCM, Vázquez RR, García FJE, Vargas JP, Castolo AA, Ferrera-Cerrato R, Guzmán OG, Calva GC (2015). The Fatty Acid Profile Analysis of Cyperus laxus Used for Phytoremediation of Soils from Aged Oil Spill-Impacted Sites Revealed That This is a C18:3 Plant Species. Plos One 10:e0140103. |
|
|
Cumming JR, Taylor GJ (1990). Mechanism of metal tolerance in plants: physiological adaptations for exclusion of metal ions from the cytoplasm. In. Alscher RG, Cumming JR (Eds.), Plant Biology, Wiley-Liss, New York 12:329-356. |
|
|
Das S, Chou ML, Jean JS, Yang HJ, Kim PJ (2017). Arsenic-enrichment enhanced root exudates and altered rhizosphere microbial communities and activities in hyperaccumulator Pteris vittata. Journal of hazardous materials 325:279-287. |
|
|
De Carvalho CC, Caramujo MJ (2018). The various roles of fatty acids. Molecules 23(10):2583. |
|
|
Deng S, Kang H, Freeman C (2011). Microplate fluorimetric assay of soil enzymes. In Dick RP (Ed.), Methods of Soil Enzymology, Madison, WI. SSSA, pp. 311-318. |
|
|
Deng S and Popova I (2011). Carbohydrate Hydrolases. In: Dick RP (ed) Methods of soil enzymology. Soil Science Society of America, Madison pp. 185-210. |
|
|
Dick RP, Kandeler E (2005). Enzymes in soils. In D. Hillel (Ed.), Encyclopedia of Soils in the Environment pp. 448-456. |
|
|
Eivazi F, Tabatabai MA (1977). Phosphatases in soils. Soil Biology and Biochemistry 9(3):167-172. |
|
|
Ekenler M, Tabatabai MA (2002). Effects of trace elements on β-glucosaminidase activity in soils. Soil Biology and Biochemistry 34(11):1829-1832. |
|
|
Farid M, Shakoor MB, Ehsan S, Ali S, Zubair M, Hanif MS (2013). Morphological, physiological and biochemical responses of different plant species to Cd stress. International Journal of Chemical and Biochemical Sciences 3(2013):53-60. |
|
|
Folch J, Lees M, Sloane-Stanley GH (1957) A simple method for the isolation and purification of total lipids from animal tissues. Journal of Biological Chemistry 226(1):497-509. |
|
|
Fu JW, Liu X, Han YH, Mei H, Cao Y, de Oliveira LM, Liu Y, Rathinasabapathi B, Chen Y, Ma LQ (2017). Arsenic-hyperaccumulator Pteris vittata efficiently solubilized phosphate rock to sustain plant growth and as uptake. J. Hazard Mater 33:68-75. |
|
|
Gadd GM (2010). Metals, minerals and microbes: geomicrobiology and bioremediation. Microbiology 156(3):609-643. |
|
|
Gao Y, Zhou P, Mao L, Zhi YE, Shi WJ (2010). Assessment of effects of heavy metals combined pollution on soil enzyme activities and microbial community structure: modified ecological dose-response model and PCR-RAPD. Environmental Earth Sciences 60(3):603-612. |
|
|
German DP, Weintraub MN, Grandy AS, Lauber CL, Rinkes ZL, Allison SD (2011). Optimization of hydrolytic and oxidative enzyme methods for ecosystem studies. Soil Biology and Biochemistry 43(7):1387-1397. |
|
|
Goyal D, Yadav A, Prasad M, Singh TB, Shrivastav P, Ali A, Dantu PK, Mishra S (2020). Effect of Heavy Metals on Plant Growth: An Overview. In: Naeem M, Ali A, Sarvajeet A, Gill S (Eds.), Contaminants in Agriculture, Springer, Cham pp. 79-101. |
|
|
Gregory PJ, Atwell BJ (1991). The fate of carbon in pulse-labelled crops of barley and wheat. Plant and Soil 136(2):205-213. |
|
|
Guo G, Zhou Q, Ma LQ (2006). Availability and assessment of fixing additives for the in situ remediation of heavy metal contaminated soils: a review. Environmental Monitoring Assessment 116:513-28. |
|
|
Hassan MK, McInroy JA, Kloepper JW (2019). The interactions of rhizodeposits with plant growth-promoting rhizobacteria in the rhizosphere: a review. Agriculture 9(7):142. |
|
|
Huang RQ, Gao SF, Wang WL, Staunton S, Wang G (2006). Soil arsenic availability and the transfer of soil arsenic to crops in suburban areas in Fujian Province, southeast China. Science of the total environment, 368(2):531-541. |
|
|
Juma NG, Tabatabai MA (1977). Effects of trace elements on phosphatase activity in soils. SSSAJ. 41(2):343-346. |
|
|
Kaba?a K, Janicka-Russak M, Burzy?ski M, K?obus G (2008). Comparison of heavy metal effect on the proton pumps of plasma membrane and tonoplast in cucumber root cells. Journal of Plant Physiology 165(3):278-288. |
|
|
Kachroo P, Shanklin J, Shah J, Whittle EJ, Klessig DF (2001). A fatty acid desaturase modulates the activation of defense signaling pathways in plants. Proceedings of the National Academy of Sciences 98(16):9448-9453. |
|
|
Karaca A, Cetin SC, Turgay OC, Kizilkaya R (2010). Effects of heavy metals on soil enzyme activities. In Sherameti, Irena, Varma, Ajit, Soil heavy metals,. Springer, Berlin, Heidelberg 19:237-262. |
|
|
Karaca A, Naseby DC, Lynch JM, (2002). Effect of cadmium contamination with sewage sludge and phosphate fertilizer amendments on soil enzyme activities, microbial structure and available cadmium. Biology and Fertility of Soils 35:428-434. |
|
|
Khan MA, Khan S, Khan A, Alam M (2017). Soil contamination with cadmium, consequences and remediation using organic amendments. Science of the Total Environment 601:1591-1605. |
|
|
Kim KR, Owens G, Naidu R, Kwon SL (2010). Influence of plant roots on rhizosphere soil solution composition of long-term contaminated soils. Geoderma, 155:86-92. |
|
|
Kleine T, Leister D (2016). Retrograde signaling: organelles go networking. Biochimica et Biophysica Acta 1857(8):1313-1325. |
|
|
Lessl JT, Guan DX, Sessa E, Rathinasabapathi B, Ma LQ (2015). Transfer of arsenic and phosphorus from soils to the fronds and spores of arsenic hyperaccumulator Pteris vittata and three non-hyperaccumulators. Plant and Soil 390:49-60. |
|
|
Liao XY, Chen TB, Lei M, Huang ZC, Xiao XY, An ZZ (2004). Root distributions and elemental accumulations of Chinese brake (Pteris vittata L.) from As-contaminated soils. Plant and Soil 261:109-116. |
|
|
Liu L, Li W, Song W, Guo M (2018). Remediation techniques for heavy metal-contaminated soils: Principles and applicability. Science of The Total Environment 633:206-219. |
|
|
Liu, X and Huang, B (2004). Changes in fatty acid composition and saturation in leaves and roots of creeping bentgrass exposed to high soil temperature. Journal of the American Society for Horticultural Science, 129(6):795-801. |
|
|
Lorenz N, Hintemann, T, Kramarewa T, Katayama A, Yasuta T, Marschner P, Kandeler E (2006). Response of microbial activity and microbial community composition in soils to long-term arsenic and cadmium exposure. Soil biology and biochemistry 38(6):1430-1437. |
|
|
Luo Q, Wang S, Sun LN, Wang H (2017). Metabolic profiling of root exudates from two ecotypes of Sedum alfredii treated with Pb based on GC-MS. Scientific reports 7(1):1-9. |
|
|
Ma LQ, Komar KM, Tu C, Zhang W, Cai Y, Kennelley ED (2001). A fern that hyperaccumulates arsenic. Nature 411:438-438. |
|
|
Miteva E (2002). Accumulation and effect of arsenic on tomatoes. Communications in Soil Science and Plant Analysis 33(11):1917-1926. |
|
|
Mongrand S, Bessoule JJ, Cabantous F, Cassagne C (1998) The C16:3/C18:3 fatty acid balance in photosynthetic tissues from 468 plant species. Phytochemistry 49:1049-1064. |
|
|
Montiel-Rozas MM, Madejón E, Madejón P (2016). Effect of heavy metals and organic matter on root exudates (low molecular weight organic acids) of herbaceous species: An assessment in sand and soil conditions under different levels of contamination. Environmental Pollution 216:273-281. |
|
|
Nguyen C (2003). Rhizodeposition of organic C by plants: mechanisms and controls. Agronomie 23:375-396. |
|
|
Nouairi I, Ammar W, Youssef N, Miled D, Habib GM, Zarrouk M (2006). Comparative study of cadmium effects on membrane lipid composition of Brassica juncea and Brassica napus leaves. Plant Science 170(3):511-519. |
|
|
Olaniran AO, Balgobind A, Pillay B (2013). Bioavailability of heavy metals in soil: impact on microbial biodegradation of organic compounds and possible improvement strategies. International Journal of Molecular Science 14:10197-10228. |
|
|
Pajuelo E, Rodríguez-Llorente ID, Dary M, Palomares AJ (2008). Toxic effects of arsenic on Sinorhizobium-Medicago sativa symbiotic interaction. Environmental Pollution 154(2):203-211. |
|
|
Paunov M, Koleva L, Vassilev A, Vangronsveld J, Goltsev V (2018). Effects of different metals on photosynthesis: cadmium and zinc affect chlorophyll fluorescence in Durum Wheat. International Journal of Molecular Science 19(3):787. |
|
|
Peixoto HP, Cambraia J, Santanna R, Mosquim PR, Moreira MA (2001). Aluminum effects on fatty acid composition and lipid peroxidation of a purified plasma membrane fraction of root apices of two sorghum cultivars. Journal of Plant Nutrition 24:1061-1070. |
|
|
Porter JR, Sheridan RP (1981). Inhibition of nitrogen fixation in alfalfa by arsenate, heavy metals, fluoride, and simulated acid rain. Plant Physiology 68(1):143-148. |
|
|
Pourrut B, Shahid M, Dumat C, Winterton P, Pinelli E (2011). Lead uptake, toxicity, and detoxification in plants. Review of Environmental Contamination and Toxicology 213:113-136. |
|
|
Rahman H, Sabreen S, Alam S, Kawai S (2005). Effects of nickel on growth and composition of metal micronutrients in barley plants grown in nutrient solution. Journal of Plant Nutrition 28(3):393-404. |
|
|
Rahman MA, Hasegawa H, Rahman M, Miah M, Tasmin A (2008). Straighthead disease of rice (Oryza sativa L.) induced by arsenic toxicity. Environmental and Experimental Botany 62(1):54-59. |
|
|
Raja V, Majeed U, Kang H, Andrabi KI, John R (2017). Abiotic stress: Interplay between ROS, hormones and MAPKs. Environmental and Experimental Botany 137:142-157. |
|
|
Sagardoy R, Morales F, López-Millán AF, Abadía A, Abadía J. (2009). Effects of zinc toxicity on sugar beet (Beta vulgaris L.) plants grown in hydroponics. Plant Biology 11:339-350. |
|
|
Shahid M, Xiong T, Masood N, Leveque T, Quenea K, Austruy A, Foucault Y, Dumat C (2014). Influence of plant species and phosphorus amendments on metal speciation and bioavailability in a smelter impacted soil: a case study of food-chain contamination. Journal of soils and sediments 14(4):655-665. |
|
|
Shaibur MR, Kitajima N, Sugawara R, Kondo T, Alam S, Huq SMI, Kawai S (2008). Critical toxicity level of arsenic and elemental composition of arsenic-induced chlorosis in hydroponic sorghum. Water, Air, and Soil Pollution 191(1):279-292. |
|
|
Singh N, Ma LQ, Srivastava M, Rathinasabapathi B (2006). Metabolic adaptations to arsenic-induced oxidative stress in Pteris vittata L and Pteris ensiformis L. Plant Science 170(2):274-282. |
|
|
Smalla K, Wieland G, Buchner A, Zock A, Parzy J, Kaiser S, Roskot N, Heuer H, Berg G (2001). Bulk and rhizosphere soil bacterial communities studied by denaturing gradient gel electrophoresis: plant-dependent enrichment and seasonal shifts revealed. Applied and Environmental Microbiology 67(10):4742-4751. |
|
|
Srivastava M, Ma LQ, Singh N, Singh S (2005). Antioxidant responses in hyperaccumulator and sensitive fern species to arsenic. Journal of Experimental Botany 56(415):1335-1342. |
|
|
Steiner F, Zoz T, Junior ASP, Castagnara DD, Dranski JAL (2012). Effects of aluminum on plant growth and nutrient uptake in young physic nut plants. Semina: Ciências Agrárias 33(5):1779-1788. |
|
|
Stoeva N, Berova M, Zlatev Z (2005). Effect of arsenic on some physiological parameters in bean plants. Biologia Plantarum 49(2):293-296. |
|
|
Tabatabai M (1977). Effects of trace elements on urease activity in soils. Soil Biology and Biochemistry 9(1):9-13. |
|
|
Tabatabai MA, Bremner JM (1970). Arylsulfatase activity of soils. Soil Science Society of America Journal 34(2):225-229. |
|
|
Talukdar D (2013). Arsenic-induced oxidative stress in the common bean legume, Phaseolus vulgaris L. seedlings and its amelioration by exogenous nitric oxide. Physiology and Molecular Biology of Plants 19(1):69-79. |
|
|
Tu C, Ma LQ, Zhang W, Cai Y, Harris WG (2003). Arsenic species and leachability in the fronds of the hyperaccumulator Chinese brake (Pteris vittata L.). Environmental Pollution 124(2):223-230. |
|
|
Upchurch RG (2008). Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotechnology. Letters 30(6):967-977. |
|
|
USEPA method 3051A (2007). US Government Printing Office, Washington, USA, |
|
|
Verdoni N, Mench M, Cassagne C, Bessoule JJ (2001). Fatty acid composition of tomato leaves as biomarkers of metal?contaminated soils. Environmental Toxicology and Chemistry 20(2):382-388. |
|
|
Wang Y, Shi J, Wang H, Lin Q, Chen X, Chen Y (2007). The influence of soil heavy metals pollution on soil microbial biomass, enzyme activity, and community composition near a copper smelter. Ecotoxicology and Environmental Safety 67(1):75-81. |
|
|
Watanabe T, Kouho R, Katayose T, Kitajima N, Sakamoto N, Yamaguchi N, Osaki M (2014). Arsenic alters uptake and distribution of sulphur in Pteris vittata. Plant, Cell and Environment 37(1):45-53. |
|
|
Wenzel WW, Unterbrunner R, Sommer P, and Sacco, P (2003). Chelate-assisted phytoextraction using canola (Brassica napus L.) in outdoors pot and lysimeter experiments. Plant and soil. 249(1):83-96. |
|
|
Wilson RA, Calvo AM, Chang P-K, Keller NP (2004). Characterization of the Aspergillus parasiticus D12-desaturase gene: a role for lipid metabolism in the Aspergillus-seed interaction. Microbiology 150:2881-2888. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


