ABSTRACT
Knowledge of the dispersion of adults and nymphs of Edessa meditabunda in soybean model is needed to determine the control actions and thus enable proper use of strategies to avoid losses in production. This research aimed to conduct probabilistic analysis of the spatial distribution patterns of adults and nymphs of E. meditabunda in two regions in soybean Bt and non-Bt. For the evaluations, the method of cloth-to- beat was used, where one sample per plot was collected randomly from each experimental area. Evaluations were performed at 7, 14, 21, 28, 35, 42, 49, 56, 63, 70, 77, 84, 91 and 98 days after emergence. For data analysis, the rates of dispersion (variance/average Morisita index and exponent k Negative Binomial Distribution) and theoretical frequency distributions (Poisson, Binomial and Negative Binomial Positive) at 5% probability were calculated. The peak population of adults and nymphs of E. meditabunda was observed in the reproductive stage of soybean Bt and non-Bt in Dourados and Douradina. From the results obtained, it can be concluded that the Bt technology had the spatial arrangement of adults and nymphs of E. meditabunda, and is set in probabilistic arrangements with negative binomial distribution (aggregate) in the two regions surveyed. In non-Bt soybeans for adults and nymphs of E. meditabunda, spatial arrangements had set in probabilistic arrangements of positive and negative binomial distribution (uniform), as the days after emergence, for the region of Dourados and Douradina.
Key words: Glycine max L., damage, horizontal dispersion, spatial arrangement.
Soybean [Glycine max (L.) Merrill] worldwide, plays the primary role of oilseed production and for consumption (Leal-Costa et al., 2008; Silva et al., 2012; Vianna et al., 2013). This fact is explained by importance of both products for animal consumption through soybean meal, and for human consumption by the oil (Silva et al., 2010). Brazil has great relevance for agribusiness, verified by the increase of cultivated areas, mainly by increasing productivity through the use of new technologies (Zadinello et al., 2012; Fonseca et al., 2013a).
For maximum potential of soybean production, it is necessary to have the best conditions for integrated pest management. Bedbugs, considered the most important pest of soybean by feeding directly from the seed, is responsible for damages that reflect the reduced production, seed quality and transmission of diseases (Belorte et al., 2003; Marsaro Junior et al., 2010; Souza et al., 2013).
The stink bug, Edessa meditabunda (Fabricius, 1974) (Hemiptera: Pentatomidae) is a major pest in many regions of Brazil, due to its occurrence from the vegetative stage to grain maturity (Corrêa-Ferreira and Panizzi, 1999; Golin et al., 2011). Adults of E. meditabunda use their mouthparts to feed on stems of soybean plants and drill the pods, seeds, soy, making them shrunken and wrinkled, affecting consequently the yield and quality of grain (Panizzi and Parra, 2009). The Edessa is the largest genus of the family Pentatomidae, and is neotropical in occurrence (Silva et al., 2006).
The damage caused by E. meditabunda when uncontrolled can cause leaf retention, reduced seed size, reduced oil content and germination and reduction in soybean yield (Lourenção et al., 1999; Gonçalves et al., 2008).
To reduce losses, control of this insect is accomplished by chemical insecticide applications from the vegetative phase of the plant and are not always efficient and selective to natural enemies (Sosa-Gomez and Silva, 2010; Fiorin et al., 2011).
To reduce the use of insecticides, there was a long search for specific technologies with low persistence in the environment, including integrated pest management, based on plant resistance to insects. With the advent of plant breeding, the genes of bacteria such as Bacillus thuringiensis (Bt) and Bacillus sphaericus, the main bodies were used to confer resistance of plants to insects on a commercial scale (Sharma et al., 2000; Theoduloz et al., 2003).
Recent advances in agricultural biotechnology have resulted in transgenic plants that are efficient alternatives and have less environmental impact for lepidopteran control in areas where they are considered pests (Williams et al., 1998; Lolas and Meza-Basso, 2006).
Studies of population dynamics provides useful information on the development models involving pest management (Gilbert et al., 1976), considering the possibility of obtaining data on the distribution of the population over a certain period of time (Odum, 1988). Therefore, such studies can be successfully employed in IPM programs (Neto et al., 1976).
The management of E. meditabunda in soybean should include the adoption of control measures based on population levels of species monitored by periodic sampling. The first step in designing a sampling plan is the knowledge of the spatial distribution of the species of interest, to establish appropriate criteria on population (Barbosa, 2003). Thus, knowledge of a fast and efficient way of sampling the pest, especially on large areas of cultivation, is essential and the MIP is satisfactorily applied (Fernandes et al., 2003).
The spatial distributions of pests in crops are aggregated, uniform and random. Such distributions are called negative binomial, Poisson and binomial positive, respectively (Barbosa and Perecin, 1982). This classification is based on the relationship between the variance and mean of the data (Elliott, 1979). However, despite the benefits obtained from the Bt cultivar, exactly how transgenic plants affect the populations of organisms in an agroecosystem is not well known (Rodrigues et al., 2010).
Knowledge of the spatial distribution of soybean stink bugs can be an important parameter to be considered to support the decision making of the place and time most suitable to increase the efficiency of their control in integrated pest management and reduce control costs and damage environmental (Kuss-Roggia, 2009).
In this context, this study aimed to evaluate the spatial distribution of adults and nymphs of E. meditabunda in soybean Bt and non-Bt in two regions under field conditions.
The experiment was conducted under field conditions in two experimental areas located at Farm Rincão Porã, the geographic coordinates are 22°14'25'' S, 54°42'60.7'' W and altitude of 403 m in the town of Dourados and Farm Boa Sorte in the geographical coordinates 22°01'07'' S, 54°32'15'' W and altitude of 310 m in the municipality of Douradina during harvest 2011/2012. The soil of the area is classified as Typic Distroferric (RH) of loamy soil. The climate, according to Köppen's humid mesothermal, is Cwa type, with annual average temperatures and precipitation ranging from 20 to 24°C and 1.250 to 1.500 mm (Fietz and Fisch, 2006). For management of two experimental areas with soybean, the no-tillage system was used, and corn (Zea mays L) was used as preceding crop. The area of this experiment was dried with glyphosate combined with mineral oil (2.0 l.ha-1 + 0.5 l.ha-1), respectively.
The seeds used were Bt soybean AL 6910 Intact RR2 PRO™ and non-Bt RR BMX Potencia®, constituting the 2 treatments in the two different locations. Sowing was performed in the experimental areas, the first area was Farm Rincão Porã, sowing was done on 22/10/2011; in Farm Boa Sorte, second area, sowing was done on 29/10/2011, with a density of 15 seeds per meter, adopting a population of approximately 300.000 plants ha-1. The row spacing was 0.50 m. For fertilizer application in both cultivar, 300 kg ha-1 of NPK formulation (2:18:18) was used. The weed and disease control was carried out with application of herbicides, fungicides, and the application of insecticides was not carried out in the areas studied.
The spatial distribution of adults and nymphs of the stink E. meditabunda was evaluated in two regions (Dourados and Douradina) with two fields each containing 100 plots in each area, each plot consisted 11 rows with 5 m long, totaling 27.5 m2 with soybean Bt and non Bt, constituting the two treatments. The sampling methodology used was the method of cloth-to-beat, consisting of two sticks of wood connected by a white cloth, with a length of 1 m and width of 1.4 m. For the samples, one end of cloth was placed between the rows of soybeans, adjusted to the base of a row of plants and other plants extended over the adjacent row. The plants of a row (0.50 m2) were shaken vigorously in order to bring down the pest insects on the cloth (Sturmer et al., 2012). Samples were taken at 7, 14, 21, 28, 35, 42, 49, 56, 63, 70, 77, 84, 91 and98 days after emergence (DAE) in soybean Bt and non-Bt.
For data analysis, the mean and variance of the number of adults and nymphs of E. meditabunda per plot were obtained at each sampling date, using the relationship between these values as an indicator of the spatial distribution (Elliott, 1979). The dispersion indexes, as described below, were calculated using the Excel® program.
Variance/mean ratio: ratio between the variance and mean (I = s2/m), used to measure the deviation of a random arrangement of conditions, where values equal to unity indicates random spatial distribution, values smaller than unity are uniform distribution and values greater than the aggregate are distribution unit (Rabinovich, 1980). The departure from randomness can be tested using the Chi-square test with n-1 degrees of freedom, χ2 = (n-1) s2/m (Elliott, 1979). Morisita index: the index of Morisita (Iδ) is relatively independent on the medium and the number of samples. So when Iδ = 1, the distribution is random, when Iδ>1, the distribution is contagious type of Iδ and when <1, it indicates a regular distribution (Morisita, 1962).
Exponent k of the negative binomial distribution: the exponent k is a suitable dispersion index when the size and numbers of sample units are the same in each sample, as often this is influenced by the size of the sampling units. This parameter is an inverse measure of the degree of aggregation, in this case, negative values indicate a regular or uniform distribution, positive, values close to zero indicate aggregate provision and higher values indicate willingness for random eight (Pielou, 1977; Southwood, 1978; Elliot, 1979). On this point, Poole (1974) used another interpretation for it when 0<k<8, the index indicates a clustered distribution, and when 0>k>8, it points to random distribution.
Theoretical frequency distribution: Theoretical frequency distributions used to evaluate the spatial distribution of the observed species are presented according to Young and Young (1998). Poisson distribution: also known as random distribution, is characterized by having variance equal to the mean (m = s2). Positive binomial distribution: This describes the uniform distribution and has less average (s2<m) variance.
Negative binomial distribution: This has higher average variance, thus indicating clumped distribution, and has two parameters: the mean (m) and the parameter k (k>0). Chi-square test of grip: To check the fit of the data collected in the field to the theoretical frequency distribution test, the Chi-square test was used to compare the total grip of the observed frequencies in the sampled area, with frequencies expected according to Young and Young (1998); these frequencies are defined by the product of the probabilities of each class and the total number of sampling units used. To conduct these tests, to fix a minimum expected frequency equal to unity was chosen. Statistical analysis was performed using Chi-square test at 1 and 5% probability test.
The presence of adult E. meditabunda on soybean plants in Dourados was detected at 35 days after emergence (DAE) in soybean cultivars with Bt and non-Bt in Dourados and Douradina (Figure 1A and 1B). In 14 samples, a total of 470 insects were found, which is 52.76% for the Bt cultivar and 47.23% in non-Bt in Dourados and a total of 415 insects in Douradina, which is 53.73% for the Bt cultivar and 46.26% for non-Bt (Figure 1A and 1B). The peak population of adults was observed at 77 DAE (month of January) for cultivars Bt and non-Bt, respectively, in Dourados and Douradina (Figure 1A and B).
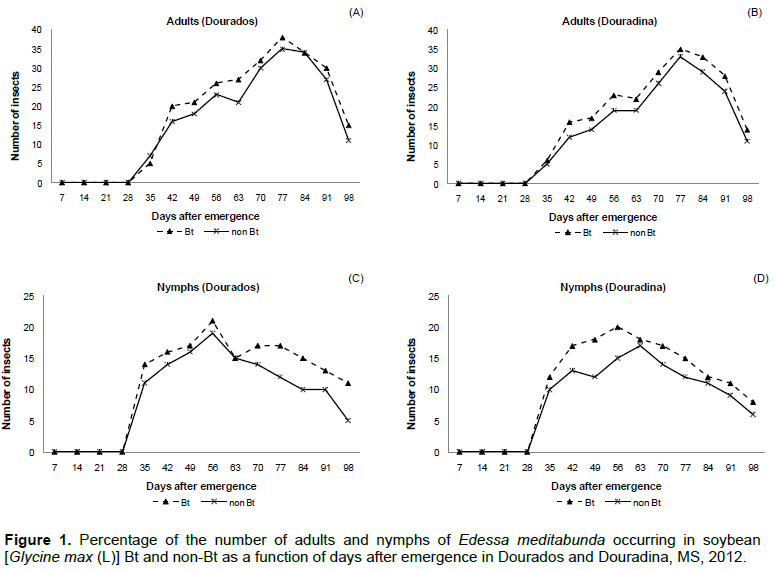
Typically, colonization of soybean fields by bedbugs starts at the end of the vegetative stage (Panizzi and Vivan, 1997), a fact that occurred in this study, which had infestations in the vegetative phase and peak population in the reproductive phase. This may be explained by the appearance of the pods; soybean becomes more nutritionally adequate for the development of bedbugs, which results in the increase of the population. The end of the development of pods and early grain filling period was considered critical (Corrêa-Ferreira and Panizzi, 1999; Husch et al., 2012).
Depending on the search for more suitable host plants for feeding, oviposition and development of their offspring, adult bed bugs migrate from one region to another, which results in different behavior and infestations, adjusting to the environment (Panizzi, 1991).
The results of the nymphs of E. meditabunda in Bt and non-Bt cultivars arose from the fifth sample, 35 DAE in Dourados and Douradina (Figure 1C and D). The largest numbers of nymphs of E. meditabunda were found in Bt cultivar, which represented 52.32% of total plots sampled in Dourados and Douradina, with 55.43% of sampled plots (Figure 1C and D). Thomazoni et al. (2010) and Barros and Degrande (2012) reported that the bedbug infestation in Bt cultivar was more intense as compared to non-Bt, requiring more sprays.
The highest population density of adults and nymphs of E. meditabunda found in Bt cultivar was due probably to the reduction of inter-specific competition between the various populations that made up the community of arthropods culture, it was caused probably by reduced caterpillar population as the major defoliating caterpillars culture as soybean caterpillar Anticarsia gemmatalis (Hübner, 1818), false-medideira caterpillar Chrysodeixis includens (Walker, 1857) and naked Rachiplusia (Guenée, 1852) and drill underarm Crocidosema aporema (Walsingham, 1914 ), those species that are regarded toxin Cry1Ac and therefore, were not found in this cultivar due to the resistance offered by these transgenic lepidopteran (Bernardi et al., 2012).
No preference of adults and nymphs of E. meditabunda were observed in relation to cultivars versus regions for soybean Bt and non-Bt. Fonseca et al. (2013) reported no difference in bedbug infestation between Bt and non-Bt varieties. Likewise, the number of attacked structures incurs no significant differences between plants Bt and non-Bt (Tomquelski, 2009; Kodama and Degrande, 2012). Cultivated areas throughout the year provide ideal conditions for the survival of polyphagous insects such as stink, whose population may increase to cause significant damage in several cultures (Corrêa-Ferreira and Panizzi, 1999; Chocorosqui, 2001).
Aggregation indices for adults of E. meditabunda
The variance/mean ratio (I) calculated for adults of E. meditabunda in cultivating Bt showed six samples in Dourados, seven in Douradina, with statistically higher values indicating the unit aggregation, and scored in four samples and three Dourados and Douradina in cultivating Bt, with values equal to unity indicating uniformity (Table 1).
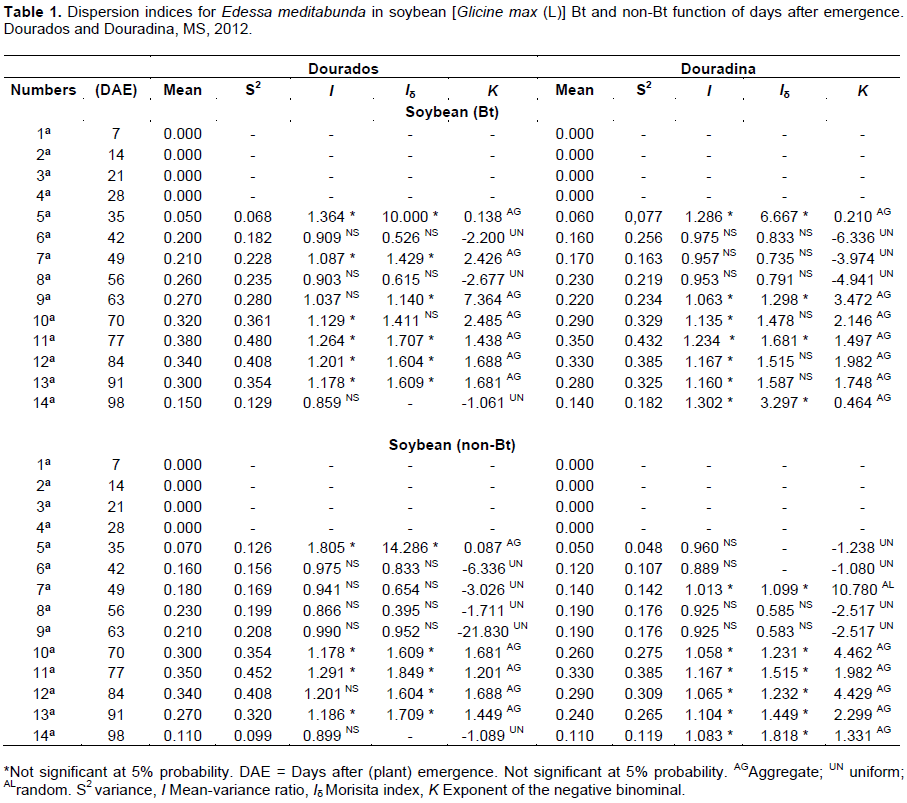
In non-Bt soybeans in Dourados, index I had five samples with statistically higher values than unity (aggregate) and six values equal to unity and theoretical distribution is uniform. In Douradina index I had six samples with statistically higher values than unity (aggregate) and five values equal to unity and theoretical distribution is uniform (Table 1).
In summary, the results of Morisita index (Iδ) for the treatment Bt, E. meditabunda adults, showed six samples in Dourados and five in Douradina, a total of ten larger than the unit (aggregate score) values. Analyzing the test Morista (Iδ) index for the non-Bt culture, Dourados was observed in ten samples, six values were greater than unity (aggregate score) and the unit in four equal values indicated uniformity; in no soy Bt in Douradina, ten samples were observed, six values were greater than unity (aggregate arrangement) and two equal to unity, tha t is, indicated uniformity (Table 1).
Analyzing the exponent K for Bt soybeans in Dourados, ten samples were found, seven indicated aggregate provision for adults of E. meditabunda, because the values were variable from 1.438 to 7.364, and three samples showed uniform with respective values -1.061, -2.200 and -2.677, while in Douradina, for this index and this cultivar, eight samples indicated aggregate provision and three uniform (Table 1).
Ten samples of soybean non-Bt were found in the field in Dourados, six of which showed aggregate provision for adults, and two showed uniform distribution. Five samples were observed in Douradina showing aggregated arrangement, uniform and five (49 DAE) had random arrangement dispersion index K (Table 1).
Aggregation indices for nymphs of E. meditabunda
The variance/mean (I) calculated for nymphs of E. meditabunda in cultivating Bt presented seven samplings in Dourados and four in Douradina, with statistically higher than the unity values, indicating aggregation, and had three samplings in Dourados, six in Douradina, in cultivating Bt, and values equal to unity indicate uniformity (Table 2). In soy non-Bt in Dourados and Douradina, the index I had five samples with statistically higher values ​​than the unity (aggregate ) and five values ​​equal to unity with theoretical distribution are uniform (Table 2).
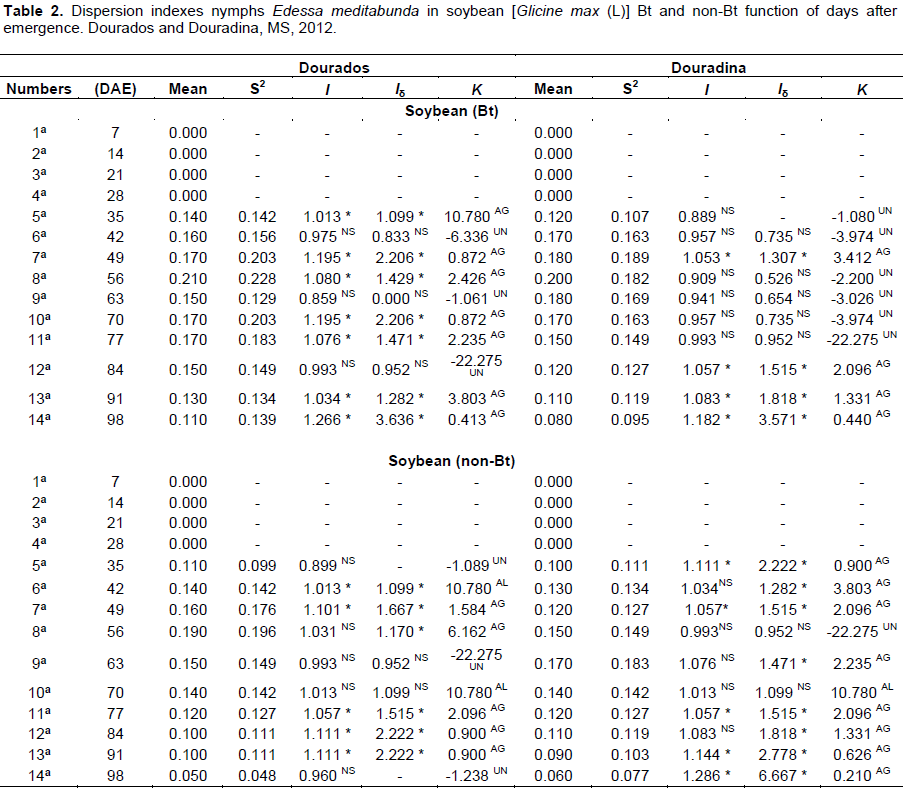

In summary, the results of Morisita index (Iδ) for the treatment Bt nymphs E. meditabunda, showed seven samples in Dourados and four in Douradina, a total of ten larger than the unity (aggregate score) values. Analyzing the test Morista (Iδ) index for the non-Bt culture, Dourados was observed to have ten samples, six values were greater than unity (aggregate arrangement) and two equal to unit with values indicating uniformity; in no soy Bt, Douradina was observed to have ten samples, eight had greater than unity (aggregate arrangement) and two identical with the drive statistical values equal to unity, is indicated uniformity values (Table 2).
Analyzing the exponent K for nymphs on biotech soybean in Dourados, showed the ten samples, seven indicated willingness to aggregate the nymphs of E. meditabunda, because the values were variable from 0.413 to 10.780, and three samples showed uniformity with respective values -1.061, -6.336 and -22.275; already in Douradina, there was found, the index and cultivar for four samples indicating aggregate willingness-six uniform arrangement (Table 2).
It was found in Dourados field that soy with no Bt, had ten samplings, indicating that five of them had nymphs aggregate provision, and three showed uniform distribution and (42 and 70 DAE) random arrangement by dispersion index K. In Douradina, eight samples were observed, indicating aggregate arrangement, uniform (56 DAE) and random (49 DAE) for the dispersion index K (Table 2).
Theoretical frequency distributions for adults of E. meditabunda
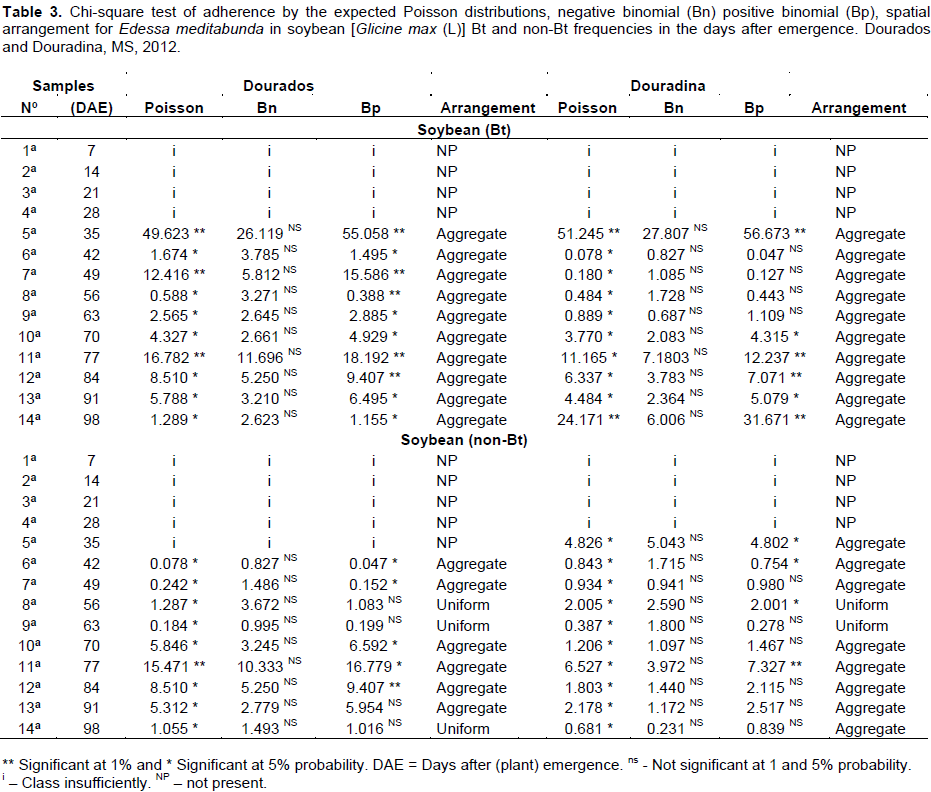
The adjustment tests of the frequencies of numerical classes for adult E. meditabunda observed in 10 samples had sufficient numbers of classes to carry out the adjustment test for Bt soybean and Dourados, Douradina (Table 3).
The values of adult E. meditabunda for cultivating Bt in Dourados and Douradina indicate that the data did not fit the theoretical models of negative binomial distribution (aggregate score) (Table 3). In the context of ecological statistics, the best fit is represented by the frequency distribution that has the lowest value of the value of Chi-square (c2) calculated (Melo et al., 2006). Like most adult samples, the value of c2 not mean of the negative binomial distribution method, consequently showed a better fit to this type of dispersion.
For non-cultivation of Bt Dourados, 10 samplings showed sufficient numbers of classes to carry out the adjustment test for the adults of E. meditabunda. The values of c2 calculated for adults indicated that the count data obtained from the field does not conform to theoretical models of Poisson distribution, and set negative binomial for six and three samples with binomial positive (Table 3).
10 samples in the region of Douradina have been found to have sufficient numbers of presented classes to carry out the adjustment test for adults E. meditabunda. The values of c2 calculated for adults E. meditabunda indicated that the count data obtained from the field does not conform to theoretical models of Poisson distribution, and adjusted for eight samples and two negative binomial positive samples (Table 3).
Theoretical frequency distributions for nymphs of E. meditabunda
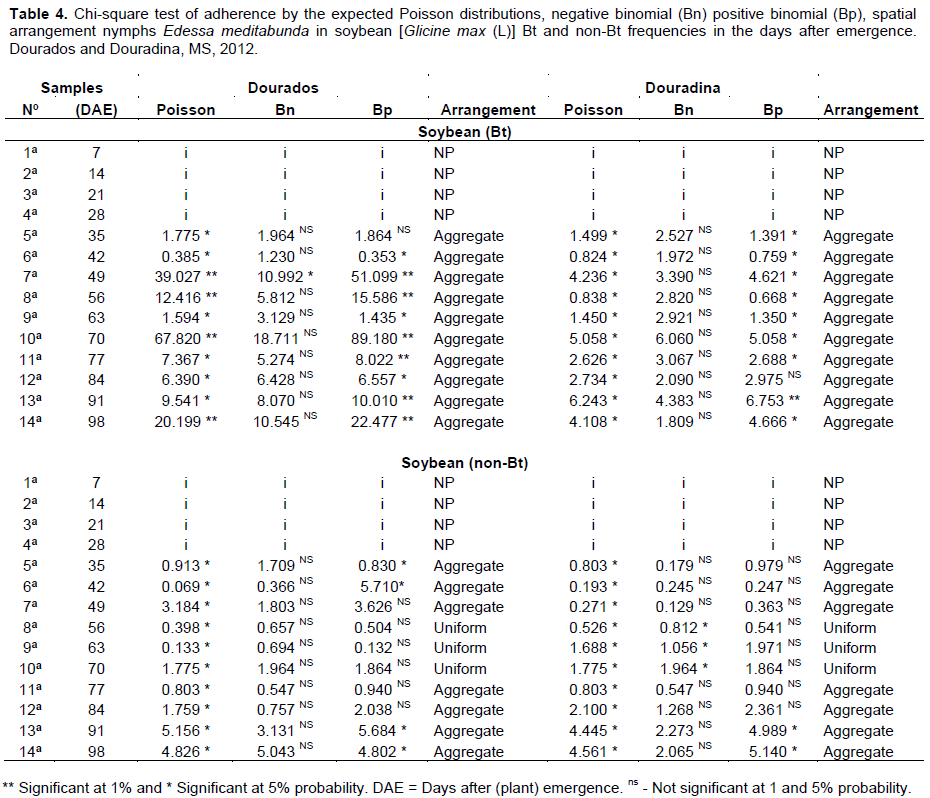
The adjustment tests of the frequencies of numerical classes for the nymphs of E. meditabunda observed in 10 samples had sufficient numbers of classes to carry out the adjustment test for Bt soybean in Dourados and Douradina (Table 4).
The values of the nymphs of E. meditabunda for Bt cultivar in Dourados and Douradina and indicate that the data fitted to theoretical models of the negative binomial distribution (aggregate score) in the samples analyzed (Table 4). In the context of ecological statistics, the best fit is represented by the frequency distribution that has the lowest value of the value of Chi-square (c2) calculated (Melo et al., 2006). Most samples of nymphs showed the value of c2 with no mean by the negative binomial distribution method, and consequently showed a better fit to this type of dispersion.
For not cultivating Bt, Dourados and Douradina 10 samples had sufficient numbers of classes to carry out the adjustment test for the nymphs of E. meditabunda. The values of c2 calculated for the nymphs indicated that the count data obtained in the field do not fit the theoretical models of the Poisson distribution, and adjusted for sampling 35, 42, 49, 77, 84, 91 and 98 DAE, in addition to the arrangement (negative binomial) and for samples of 56, 63 and 70 DAE, the uniform arrangement (positive binomial) (Table 4).
The colonization of soybean fields, by bedbugs often begins at the edges due to their migration from one area to another in soybean under different stages of development and areas with alternative hosts (Panizzi et al., 1980; Panizzi, 1991). This dynamic shift has been studied (Kuss-Roggia, 2009; Guedes et al., 2012).According to Panizzi and Vivan (1997), in soybean crops, leave eating bed bugs infestations occur in the vegetative phase, which agrees with the data of the present work, in which at 28 DAE there was infestations of adults and nymphs of E. meditabunda.
With the emergence of legumes, soy becomes more nutritionally adequate for the development of bedbugs, which results in the increase of the population, especially nymphs (Panizzi and Vivan, 1997). Thus there is increase in the population of adults and nymphs of E. meditabunda with the development of vegetables, having their peak population during grain filling. These data demonstrated the importance of management of bedbugs in agricultural areas, contributing to the realization of chemical or biological control by the time it reaches the economic injury level, and seeking to reduce the population peaks.
Likewise, it is very important to understand the dispersion and spatial distribution of soybean stink bugs, the study of these various indices of dispersion is used to calculate the spatial arrangement of a pest (Barbosa, 2003), and as a single index, it does not provide all considered ideal attributes of a statistical standpoint; the use of more than one index can provide greater certainty to conclusions (Martins et al., 2012).
The spatial arrangement of adults and nymphs of E. meditabunda in soybean Bt, when there is sufficient number of tests of fit, the theoretical frequency distribution classes proved to be aggregated in the two study areas. According to Rodrigues et al. (2010) and Fonseca et al. (2013b) report, aggregate behavior of adult insects on cultivation genetically, is modified. However, with respect to non-Bt soybean, adults and nymphs of E. meditabunda set the distribution of aggregate type and even the theoretical frequency distribution. These results are in agreement with Souza et al. (2013) that relate the spatial arrangement of the stink bug adults and nymphs in soybean fields.
It is possible to conclude that the spatial arrangement was influenced by cultivar genetically modified, versus days after emergence, since, in both cultivars, the individuals in the populations were assessed based on differences presented in the form of aggregated and uniform spatial distribution.
The aggregate spatial distribution model for the soybean crop requires a larger number of sample units than any other type of spatial distribution when performing a sampling process. The precise number of sample units for the sampling process of these insects is extremely important since these species are causing significant direct damage to the soybean crop.
So as to control these insect pests, it is possible to recommend the application of selective insecticides when necessary in localized areas of culture, where groups of these individuals are found. With this point, the number of sample units is dependent on the required degree of accuracy, which varies according to the purpose of research: population dynamics, crop damage, levels of economic development, losses and pest control (Silva and Costa, 1998; Guedes et al., 2012). Furthermore, such information should be considered in developing new sampling plans for pest to reduce the sampling time and increase the reliability of the results (Grigolli et al., 2012).
The peak population of adults and nymphs of E. meditabunda was observed in the reproductive stage of soybean Bt and non-Bt in Dourados and Douradina. From the results obtained, it can be concluded that the Bt technology had the spatial arrangement of adults and nymphs of E. meditabunda, and is set in probabilistic arrangements as negative binomial distribution (aggregate) in the two regions surveyed. In non-Bt soybeans for adults and nymphs of E. meditabunda, spatial arrangements had set in probabilistic arrangements for negative (aggregate) and positive binomial distribution (uniform), as the days after emergence, for the regions of Dourados and Douradina.
The authors have not declared any conflict of interest.
REFERENCES
|
Barbosa JC, Perecin D (1982). Modelos probabilísticos para distribuições de lagartas de Spodoptera frugiperda (J. E. Smith, 1797), na cultura do milho. Científica 10:181-191.
|
|
|
|
Barbosa JC (2003). Métodos estatísticos aplicados à entomologia. 1 edn, 254 p. Jaboticabal, SP: FUNEP.
|
|
|
|
|
Barros R, Degrande PE (2012). Desempenho do algodão-Bt como tática de controle de pragas em condições de campo. Científica 40:117-137.
|
|
|
|
|
Belorte LCC, Ramiro ZA, Faria AM (2003). Levantamento de percevejos pentatomídeos em cinco cultivares de soja [Glycine max (L.) Merrill, 1917] no município de Araçatuba, SP. Arq. Inst. Biol. 70:447-451.
|
|
|
|
|
Bernardi O, Malvestiti G, Dourado PM, Oliveira WS, Martinelli S, Berger GU, Head GP, Omoto C (2012). Assessment of the high-dose concept and level of control provided by MON 87701 x MON 89788 soybean against Anticarsia gemmatalis and Pseudoplusia includens (Lepidoptera: Noctuidae) in Brazil. Pest Manag. Sci. 68:1083-1091.
Crossref
|
|
|
|
|
Chocorosqui VR (2001). Bioecologia de espécies de Dichelops (Diceraeus) (Heteroptera: Pentatomidae) e danos em soja, milho e trigo no Norte do Paraná. Tese (Doutorado em Entomologia) - Universidade Federal do Paraná. 158 p.
|
|
|
|
|
Corrêa-Ferreira BS, Panizzi AR (1999). Percevejos da soja e seu manejo. Londrina, Embrapa-CNPSo, 45 p. (Circular técnica, 24).
|
|
|
|
|
Elliott JM (1979). Some methods for the statistical analysis of sample benthic invertebrates. 1 edn, 157 p. Ambleside, Freshwater Biological Association.
|
|
|
|
|
Fernandes MG, Busoli AC, Barbosa JC (2003). Distribuição espacial de Alabama argillacea (Hübner) (Lepidoptera: Noctuidae) em algodoeiro. Neotrop. Entomol. 32:107-115.
Crossref
|
|
|
|
|
Fiorin RA, Stürmer GR, Guedes JVC, Costa IFD, Perini CR (2011). Métodos de aplicação e inseticidas no controle de percevejos na cultura da soja. Semina. Ciências Agrárias 32:139-146.
Crossref
|
|
|
|
|
Fietz CR, Fisch GF (2006). O clima da região de Dourados, MS. Dourados, MS, Brasil: Embrapa Agropecuária Oeste, 1 edn. 32 p. Documentos, 85.
|
|
|
|
|
Fonseca PRB, Fernandes MG, Degrande PE, Mota TA, Kassab SO (2013a). Spatial distribution of adult Anthonomus grandis Boheman (Coleoptera: Curculionidae) and buds with feeding punctures on conventional and Bt cotton. Semina. Ciênc. Agrárias 34:1129-1138.
|
|
|
|
|
Fonseca PRB, Fernandes MG, Justiniano W, Cavada LH, Silva JAN (2013b). Leaf Chlorophyll Content and Agronomic Performance of Bt and Non-Bt Soybean. J. Agric. Sci. 5:117-125.
Crossref
|
|
|
|
|
Gilbert N, Gutierrez AP, Frazer BD, Jones RE (1976). Ecological relationships. San Francisco, W.H. Freeman. 256 p.
|
|
|
|
|
Golin V, Loiacono MS, Margaria CB, Aquino DA (2011). Natural incidence of egg parasitoids of Edessa meditabunda (F.) (Hemiptera: Pentatomidae) on Crotalaria spectabilis in Campo Novo do Parecis, MT, Brazil. Neotrop. Entomol. 40:617-618.
|
|
|
|
|
Gonçalves L, Almeida FS, Mota FM (2008). Efeitos da temperatura no controle, desenvolvimento e reprodução de Edessa meditabunda (Fabricius, 1764) (Hemiptera: Pentatomidae). Acta Biol. Par. 37:111-121.
|
|
|
|
|
Grigolli JFJ, Souza LA, Fraga DF, Busoli AC (2012). Boll weevil feeding preference on squares at different ages and square shedding time of cotton cultivars. Afr. J. Agric. Res. 7:4317-4323.
|
|
|
|
|
Guedes JVC, Arnemann JA, Stürmer GR, Melo AA, Bigolin M, Perini CR, Sari BG (2012). Percevejos da soja: novos cenários, novo manejo. Revista Plantio Direto 1:28-34.
|
|
|
|
|
Husch PH, Oliveira MCN, Sosa-Gómez DR (2012). Danos ocasionados por Edessa meditabunda (F.) e Piezodorus guildinii (West.) (Hemiptera: Pentatomidae) em sementes de soja. In: CONGRESSO BRASILEIRO DE SOJA, 6., 2012, Cuiabá. Anais. Londrina: Embrapa Soja.
|
|
|
|
|
Kodama E, Degrande PE (2012). Não-preferência para oviposição e viabilidade de ninfas de Bemisia tabaci Biótipo B (Hemiptera: Aleyrodidae) em algodão-Bt e em sua isolinha não-transgênica. Interciencia (Caracas) 37:377-380.
|
|
|
|
|
Kuss-Roggia RCR (2009). Distribuição espacial e temporal de percevejos da soja e comportamento de Piezodorus guildinii (Westwood, 1837) (Hemiptera: Pentatomidae) na soja (Glycine max (L.) Merrill) ao longo do dia. Tese (Doutorado em Agronomia) – Universidade Federal de Santa Maria.
|
|
|
|
|
Leal-Costa MV, Aragão FJL, Reinert F, Tavares ES (2008). Anatomia foliar de plantas transgênicas e não transgênicas de Glycine max (L.) Merrill (Fabaceae). Revista Biociências 14:23-31.
|
|
|
|
|
Lourenção AL, Pereira JCNA, Miranda MAC, Ambrosiano GMB (1999). Danos de percevejos e de lagartas em cultivares e linhagens de soja de ciclos médio e semi-tardio. Anais Soc. Entomol. Bras. 28:157-167.
Crossref
|
|
|
|
|
Lolas LN, Meza-Basso L (2006). Evaluation of native strains of Bacillus thuringiensis as an alternative of integrated management of the tomato leaf miner Tuta absoluta Meyrick; Lepidoptera: Gelechiidae in Chile. Agric. Téchnol. 66:235-246.
|
|
|
|
|
Marsaro Júnior AL, Silva Pereira PRV, Silva WR, Griffel SCP (2010). Flutuação populacional de insetos-praga na cultura da soja no estado de Roraima. Rev. Acad. Ciênc. Agrár. Ambient. 8:71-76.
|
|
|
|
|
Martins GLM, Vieira MR, Barbosa JC, Dini TA, Manzano AM, Alves BMS, Silva RM (2012). Distribuição espacial de Calacarus Heveae feres na cultura da seringueira em Marinópolis - São Paulo. Rev. Árvore 36:211-217.
Crossref
|
|
|
|
|
Melo EP, Fernandes MG, Degrande PE, Cessa RMA, Salomão JL, Nogueira RF (2006). Distribuição espacial de plantas infestadas por Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) na cultura do milho. Neotrop. Entomol. 35:689-697.
Crossref
|
|
|
|
|
Morisita M (1962). Id-index, a measure of dispersion of individuals. Res. Popul. Ecol. 4:1-7.
Crossref
|
|
|
|
|
Odum EP (1988). Ecologia 1 ed., 434 p. Rio de Janeiro: Guanabara Koogan.
|
|
|
|
|
Panizzi AR, Vivan LM (1997). Seasonal abundance of the neotropical brown stink bug, Euschistus heros in overwintering sites and the breaking of dormancy. Entomol. Exp. Appl. 82:213-217.
Crossref
|
|
|
|
|
Panizzi AR (1991). Ecologia nutricional de insetos sugadores de sementes. In: Panizzi, A.R., and J.R.P. Parra. 1991. Ecologia nutricional de insetos e suas implicações no manejo de pragas. São Paulo: Manole Cap. 7:253-287.
|
|
|
|
|
Panizzi AR, Galileo MHM, Gastal HAO, Toledo JFF, Wild CH (1980). Dispersal of Nezara viridula and Piezodorus guildinii nymphs in soybeans. Environ. Entomol. 9:293-297.
Crossref
|
|
|
|
|
Panizzi AR, Parra JRP (2009). A bioecologia e nutrição de insetos como base para o manejo integrado de pragas p.1107-1140. In Panizzi, A.R., e J.R.P. Parra. (eds) Bioecologia e nutrição de insetos: base para o manejo integrado de pragas. Brasília, Embrapa/CNPq, 1164 p.
|
|
|
|
|
Pielou EC (1977). Mathematical ecology. 2edn, New York: John Wiley & Sons. 385 p
|
|
|
|
|
Poole RW (1974). An introduction to quantitative ecology. 1edn, New York: McGraw Hill. 525p.
|
|
|
|
|
Rabinovich JE (1980). Introducion a la ecologia de problaciones animales. 1edn, 313 p. México, CECSA.
|
|
|
|
|
Rodrigues TR, Fernandes MG, Santos HR (2010). Distribuição espacial de Aphis gossypii (Glover) (Hemiptera, Aphididae) e Bemisia tabaci (Gennadius) biótipo B (Hemiptera, Aleyrodidae) em algodoeiro Bt e não-Bt. Rev. Bras Entomol. 54:136-143.
Crossref
|
|
|
|
|
Sharma HC, Sharma KK, Seetharama N, Ortiz R (2000). Prospects for using transgenic resistance to insects in crop improvement. Electronic J. Biotechnol. 3:1-26.
Crossref
|
|
|
|
|
Silva JPGF, Baldin ELL, Souza ES, Lourenção AL (2012). Assessing Bemisia tabaci (Genn.) biotype B resistance in soybean genotypes: Antixenosis and antibiosis. Chilean J. Agric. Res. 72:516-522.
Crossref
|
|
|
|
|
Silva CO, Andrade GF, Dantas MIS, Costa NMB, Peluzio MCG, Martino HSD (2010). Influência do Processamento na Qualidade Proteica de Novos Cultivares de Soja destinados à Alimentação Humana. Revista de Nutrição 23:239-397.
Crossref
|
|
|
|
|
Neto SS, Nakano O, Barbin D, Villa Nova eNA (1976). Manual de Ecologia dos Insetos 1 ed., 419 p. São Paulo: Agronômica Ceres.
|
|
|
|
|
Silva EJE, Fernandes JAM, Grazia J (2006). Caracterização do grupo Edessa rufomarginata e descrição de sete novas espécies (Heteroptera, Pentatomidae, Edessinae). Iheringia Série Zoologia 96:345-362.
Crossref
|
|
|
|
|
Silva MTB, Costa EC (1998). Tamanho e número de unidades de amostra de solo para amostragem de larvas de Diloboderus abderus (Sturm) (Coleoptera: Melolonthidae) em plantio direto. Anais da Sociedade Entomológica do Brasil 27:193-197.
Crossref
|
|
|
|
|
Sosa-Gómez DR, Silva JJ (2010). Neotropical brown stink bug (Euschistus heros) resistance to methamidophos in Paraná, Brazil. Pesq. Agropecu. Bras. 45:767-769.
Crossref
|
|
|
|
|
Southwood TRE (1978). Ecological methods. 2 edn, 525 p., New York: John Wiley & Sons.
|
|
|
|
|
Souza ES, Baldin ELL, Silva JPGF, Lourenção AL (2013). Feeding preference of Nezara viridula (Hemiptera: Pentatomidae) and attractiveness of soybean genotypes. Chilean J. Agric. Res. 73:351-357.
Crossref
|
|
|
|
|
Souza LA, Barbosa JC, Grigolli JFJ, Fraga DF, Maldonado W, Busoli AC (2013). Spatial Distribution of Euschistus heros (F.) (Hemiptera: Pentatomidae) in Soybean. Neotrop. Entomol. 42:412-418.
Crossref
|
|
|
|
|
Sturmer GR, Cargnelutti A, Filho L, Stefanelo S, Guedes JVC (2012). Eficiência de métodos de amostragem de lagartas e de percevejos na cultura de soja. Cienc. Rural 42:2105-2111.
Crossref
|
|
|
|
|
Theoduloz, C, Vega A, Salazar M, González E, Meza-Basso L (2003). Expression of a Bacillus thuringiensis δ-endotoxin cry1Ab gene in Bacillus subtilis and Bacillus licheniformis strains that naturally colonize the phylloplane of tomato plants (Lycopersicon esculentum Mill.). J. Appl. Microbiol. 94:1-7.
Crossref
|
|
|
|
|
Thomazoni D, Degrande PE, Silvie P, Faccenda O (2010). Impact of Bollgard® genetically modified cotton on the biodiversity of arthropods under practical field conditions in Brazil. Afr. J. Biotechnol. 9:6167-6176.
|
|
|
|
|
Tomquelski GV (2009). Ocorrência de pragas e custo de produção em algodoeiro geneticamente modificado (Bt) e convencional. Tese (Doutorado em Agronomia) – Universidade Estadual Paulista Júlio de Mesquita Filho, Ilha Solteira. 98p.
|
|
|
|
|
Vianna VF, Trevisoli SHU, Desiderio JA, Santiago S, Charnai K, Ferreira Junior JAF, Ferraudo AS, Mauro AO (2013). The multivariate approach and influence of characters in selecting superior soybean genotypes. Afr. J. Agric. Res. 8:4162-4169.
|
|
|
|
|
Williams JW, Summers RL, Webb T (1998). Applying plant functional types to construct biome maps from eastern North American pollen data: comparisons with model results. Quat. Sci. Rev. 17:607-628.
Crossref
|
|
|
|
|
Young LJ, Young JH (1998). Statistical ecology: a population perspective. 1 edn, Boston: Kluwer Academic Publishers. 565p.
Crossref
|
|
|
|
|
Zadinello R, Chaves MM, Santos RF, Bassegio D, Werncke eI (2012). Influência da aplicação de Glifosato na produtividade da soja. Acta Iguazu 1:1-8.
|
|