Full Length Research Paper

ABSTRACT
Cowpea production is common among smallholder subsistence farmers of Western Kenya due to its wide ecological adaptation. However, this crop is affected by cowpea rust Uromyces phaseoli var vignae (Baarel) Arth disease, causing economic yield losses with limited control measures. This study therefore, evaluated cowpea rust disease incidence (DI) and severity (DS) on growth of selected improved cowpea genotypes. A Randomized Complete Block Design (RCBD) in split plot arrangement experiment was conducted in Busia and Kakamega counties, Kenya during short rains of 2018 and long rains of 2019. Cowpea genotype: K80, KVU 27-1, Tumaini, Dakawa and local variety was the main factor and cropping system: pure stand cowpea and cowpea intercropped with maize was the sub plot-factor. Data were subjected to Mixed model ANOVA using SAS. The DI and DS were 41 and 39% respectively less in Dakawa and Tumaini with more cowpea leaf and grain yields compared to other genotypes. On the other hand, DI and DS were 35 and 56% respectively less in pure stand cowpea compared to intercrop cowpea. Pure stand cowpea also had more leaf and grain yields. The results indicate that Dakawa and Tumaini cowpea genotypes have potential resistance to cowpea rust and the conditions could be improved by planting cowpea in pure stand.
Key words: Cowpea genotype, cropping system, cowpea rust incidence, severity.
INTRODUCTION
The production of cowpea is more common among the subsistence smallholder farmers because of its wide ecological adaptation and tolerance to several biotic and abiotic stresses that include pests, diseases and drought. It is an important food source and is estimated to be the major protein source for more than 200 million people in sub-Saharan Africa. It is of major importance to the livelihoods of millions of relatively poor people in less developed countries of the tropics (FAO, 2013). Additionally, cowpea has many ecological benefits such as nitrogen fixation, heat and drought-tolerant crop, cover crop (Saidi et al., 2010). In addition, some cowpea varieties cause suicidal germination of the seed of Striga hermonthica, a parasitic plant that usually infests cereals with devastating effects in sub-Saharan Africa (Quin, 1997).
About a decade ago, world cowpea production was 5.72 million tons of which Africa produced 5.42 million tons; East Africa, 0.52 million tons and Kenya produced about 122,682 tons (FAO, 2013). In Kenya, cowpea grain production is estimated to be 0.53 tons/ha against the estimated production potential of 1.6 ton/ ha. The situation is worse for Western Kenya where yields are much lower than the average 0.53 tons/ ha. This scenario indicates that there is a huge yield gap in cowpea grain production in Kenya. There are a number of constraints that hinder sustainable cowpea production contributing to the production levels that are below production potential. Singh et al. (1997) reported that diseases, insect pests and parasitic weeds are the most important factors responsible for low cowpea yield in Nigeria and the same could explain the low cowpea yields in Kenya. One major disease that has been identified as a big threat to Cowpea production in East Africa particularly in Kenya and needs major attention is cowpea rust (Allen et al., 1998).
Cowpea rust caused by a fungus, Uromyces phaseoli var vignae (Baarel) Arth appeared in the late 1990s and occurs widely in Kenya. The disease interferes with normal root development and uptake of nutrients by plant roots resulting in reduced seed size and considerable yield loss. Many control measures that have been used against the disease have yielded limited impact. The most preferred control measure by many farmers is the chemical control using fungicides. The application of fungicides several times within a growing season by farmers to obtain a clean crop has been proofed not to be economical due to small land size in addition to negative environmental impacts as well as safety concerns (Mensah et al., 2018). Breeding and release of resistant cowpea cultivars has been found to be one of the cheap and most effective methods to control cowpea rust. Due to variations in genetic composition, significant differences have been found among cowpea genotypes on response to cowpea rust disease infestation (Mensah et al., 2018). Furthermore, new rust resistance cowpea genotypes released usually lose their resistance within a short period of time (Mensah et al., 2018). This means that new genotypes should be screened for their variability in response to rust infestation and those with high levels of resistance have to be bred constantly to replace the ones that are becoming or already susceptible.
Use of appropriate cropping system has been proven to provide numerous environmental benefits as well as contribute to pest and disease management (Ding et al., 2015). Cowpea is extensively grown in the low lands and mid altitude regions of Africa as either pure stand crop or are intercropped with cereals like sorghum, millet or maize. Research indicates that planting cowpea in pure stand yields more than intercrop (Francis, 1986). Despite the higher yields from pure stand cropping system, land is limited in western Kenya and this compels farmers to adopt alternative cropping systems like intercropping system.
Intercropping that entails growing two or more crops together on the same land during the same season is a traditional cropping system that is widespread among subsistence farming communities. Although many crops are intercropped, legume intercropping is common because legumes have the potential of biological nitrogen fixation (Vanlauwe et al., 2016). Other benefits of intercropping include: increase in yield per unit area and increase in economical returns as compared to pure stand crops. In addition, it is generally believed that one component of an intercropping system may act as a barrier or buffer against the spread of pests and disease causing pathogens within the intercropping system (Henrik and Peter, 1997). For instance intercropping maize-cowpea has been found to reduce the stem borer attack on maize (Henrik and Peter, 1997). In other instances, the canopy in the intercropping arrangement may create a favourable microclimate for the proliferation, infection and spread of disease causing pathogens. Thus, the effects of intercropping on the degree of infestation and spread of diseases have yielded varied results (Boudreau, 2013).
Sustainable cropping system is achieved when the cropping system design is based on the most important crops for particular region. Since maize is an importance staple food crop in many parts of Kenya especially western Kenya, development of sustainable cropping system for management of cowpea rust disease built around maize production is necessary. It is worth noting that at present, there is no single-cost efficient control measure to prevent rust infection particularly in different agro-ecological areas (Mensah et al., 2018). However, there is a need to bridge cowpea yield gap in order to improve households’ food and nutrition, income as well as protecting the environment. In this regard, the study seeks to evaluate cowpea rust disease incidence and severity on growth of selected improved cowpea genotypes under two cropping systems in Western Kenya.
MATERIALS AND METHODS
Experimental sites
The experiment was conducted in Kakamega and Busia Counties of western Kenya. In Kakamega County, the experiment was conducted at Kenya Agricultural, Livestock Research Organisation (KALRO) Kakamega station (00°16.9' N, 034° 46.07'E). In Busia County, the experiment was conducted at KALRO Alupe station (00° 28.0'N, 34° 07.00'E). The soil in Kakamega is classified as Ferralic-orthic acrisol (Jaetzold et al., 2007), deep, well drained highly weathered soil with inherently moderate fertility whereas the soil in Busia is well drained, very deep, dark red Orthic ferralisols (Jaetzold et al., 2007). Both soils are poor in nutrients, thus require regular fertilization. Kakamega and Busia sites represent the Upper Midlands 2 (UM 2) and Low Midlands 2 (LM 2) respectively with an altitude of approximately 1430 m and 1170 m a.s.l respectively (Jaetzold et al., 2007). The two counties have a generally cool wet climate receiving bimodal annual rainfall ranging between 1250-2000 mm in Kakamega and 760 -1750 mm in Busia and temperature range of between 14-27°C in Kakamega and 19-31°C in Busia (Jaetzold et al., 2007).
Experimental design
The experimental design was Randomized Complete Block Design (RCBD) in split plot arrangement. The main plot treatment was five cowpea genotypes namely K80, KVU27-1, Dakawa, Tumaini and local variety while sub-plot treatment was two cropping system namely pure stand and cowpea-maize intercrop resulting in a total of ten treatment combinations with three replications. The sub-plots measured 3 m wide and 5 m long and 0.5 m in between the subplots while the main plots were 1m apart. Two cowpea seeds were planted manually at recommended standard spacing of 0.6 m × 0.1 m in pure stand and 0.75 m x 0.3 m in intercrop respectively (KARI, 2000), resulting in 6 rows of cowpea plants in pure stand and 4 rows in intercrop. The plants were thinned to one plant per hole after emergence.
Parameters measured
All measurements were taken within the net area comprising two middle rows. Disease incidence was assessed using a quadrat measuring 1m by 1m which was thrown randomly within the sub-plot and the area under the quadrat analyzed. The incidence was described as the proportion of rust infected plants to the total number of plants in the quadrate and was scored using a scale of 0-9 (Mayee and Datar, 1986).
Disease severity was rated as a percentage of leaf area affected in the lower, mid and upper canopy of each of the plants under quadrat using a 0-8 visual scale score method in which a rating of 0 = no disease, 1 = disease severity up to 2.5%, 2 = disease severity 2.5-5%), 3 = disease severity 5-10%, 4 = disease severity 10-15%, 5 = disease severity 15-25%, 6 = disease severity 25-35%, 7= disease severity 35-67.5% and 8= disease severity 67.5-100%. Diseases incidences and severity were scored at an interval of three weeks starting from the first week after planting to the 15th week after planting marking the physiological maturity.
Numbers of leaves were determined by visual counting of all fully opened leaves on each of ten (10) randomly selected and tagged plants at an interval of three weeks starting from the first week after plant emergence up to the 15th week at physiological maturity (Agbogidi and Ofuoku, 2005). Leaf area measurements were taken at the same time with leaf count on the same plants. Leaf area (LA) was calculated as the product of the length and breadth at the broadest point of the longest leaf on the plant multiplied by 0.75, according to Abukutsa (2007). Leaf Area Index (LAI) was then calculated by dividing the LA by spacing.
Number of pods per plant was assessed in the field by visual counting of all pods on the ten (10) selected and tagged plants at physiological maturity. The numbers of pods were then divided by the number of cowpea plants to get number of pods per plant. Green leaf biomass was determined at physiological maturity. Leaf biomass samples were taken by plucking all fully opened mature leaves from each plant in the net area. Fresh weight was taken and recorded using an electronic balance (Woomer et al., 2011). Pods were harvested within the net area at physiological maturity. Pods were harvested with hand and fresh weights of pods recorded. The pods were air dried to a constant weight and then shelled and the weight of grains recorded. Grain yield in kg ha-1 was standardized to 13 % storage moisture content.
Data analysis
Data collected were subjected to analysis of variance to determine the effect of cowpea genotype, cropping system and their interaction on plant height, number of leaves, Leaf area index, disease incidences and severity, number of pods per plant, leaf and grain weight using mixed model (Mixed procedure SAS Institute 2012). The means were compared using Least Significance Difference (LSD).
RESULTS
Number of leaves, leaf area index, and disease incidence and severity
The two experimental sites of Busia and Kakamega which represent different agroecosystems in western Kenya showed a wide variability on weather conditions and therefore data were analyzed separately for each site. There was no seasonal effect on variables measured and therefore data were average across the seasons for each site. The mixed model ANOVA showed that both the main effects of genotype and that of cropping system on the number of leaves, disease incidence (DI), disease severity (DS) and leaf area index (LAI) were significant (P≤ 0.05) at both sites. Generally DI, DS and LAI were higher in Kakamega than Busia for all the genotypes under study. In Busia, Dakawa cowpea genotype had significantly 8% more leaves than K80, KVU 27-1, Tumaini and local variety genotypes (Table 1). Though Dakawa had higher number of leaves in Kakamega, the difference was not significant (Table 2).
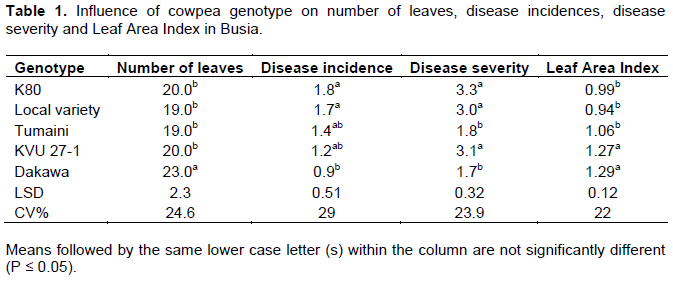
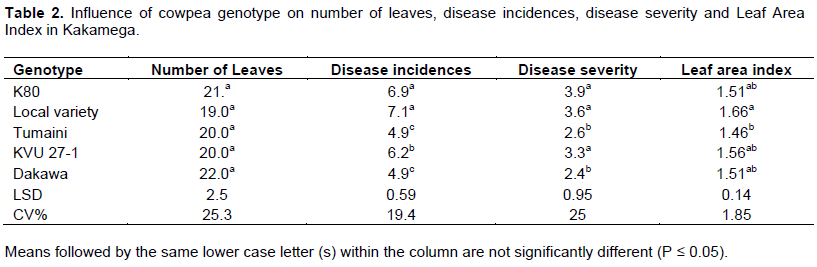
In assessing cowpea rust incidence, rust severity and leaf area index in Busia, Dakawa and Tumaini had comparable DI and DS which were 50 and 39% respectively low compared to K80, Local variety and KVU27-1 (Table 1). Similar trend was observed in Kakamega (Table 2).
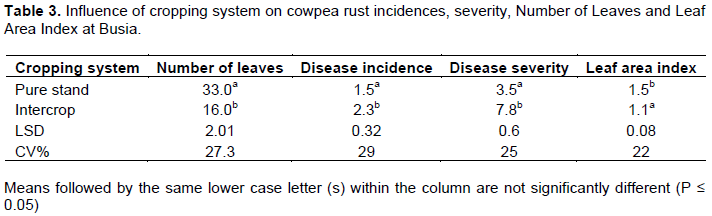
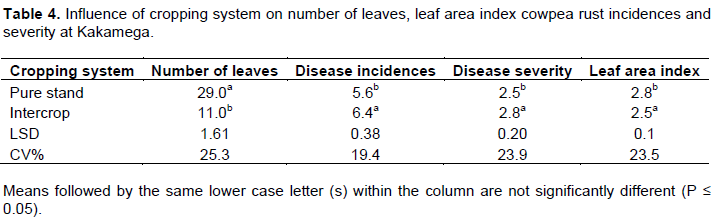
In Busia, cowpea plants had 2 times more leaves in pure stand than intercrop cowpea (Table 3). Disease incidence and disease severity were 35 and 56% respectively less in pure stand cowpea compared to intercrop cowpea. While LAI was 36% significantly higher in pure stand cowpea compared to intercrop cowpea. A similar trend was observed in Kakamega where the number of leaves was 3 times more in pure stand compared to intercrop cowpea (Table 4). Disease incidence and severity were 12 and 10% respectively less in pure stand compared to intercrop cowpea and leaf area index was 13% more in pure stand cowpea.
Interaction effects of cowpea genotype and cropping system on disease incidence and severity
The mixed model ANOVA showed that the interaction effect of cowpea genotype and cropping system on disease incidence (DI) and severity (DS) was significant (P≤ 0.05). In Busia, both DI and DS were significantly lowest in Dakawa cowpea genotype planted as pure stand compared to other treatment combinations (Figure 1). Similar trend was observed in Kakamega (Figure 2).
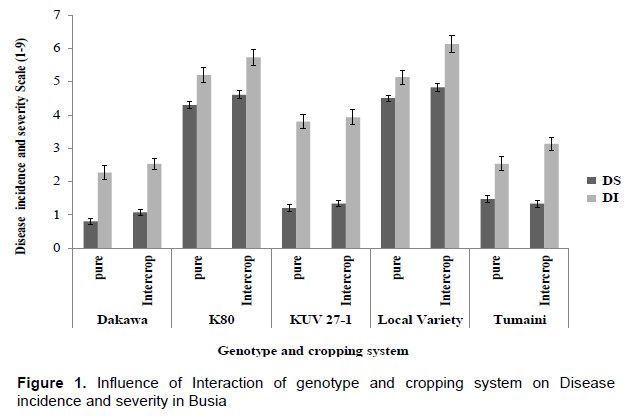
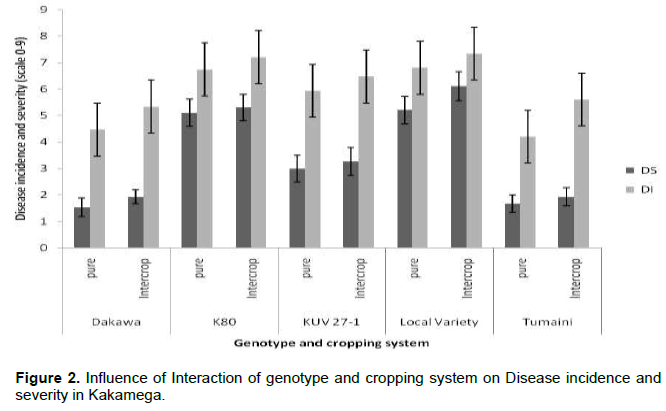
Influence of cowpea genotype on yield and yield components
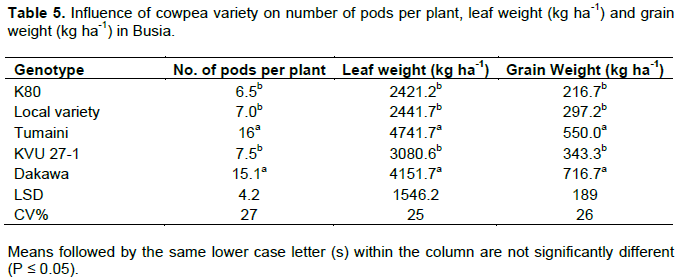
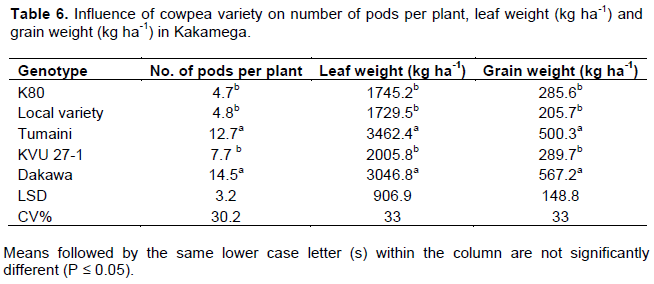
Main effect of genotype on number of pods per plant, leaf and grain yields was significant at both Busia and Kakamega (P ≤ 0.05). In Busia, Dakawa and Tumaini were statistically similar and had 69, 49 and 70% more pods per plant, leaf yield and grain yield respectively compared to statistically similar K80, local variety and KVU 27-1 (Table 5). In Kakamega similar trends were observed in which Dakawa and Tumaini had 68, 43 and 50% more pods per plant, leaf yield and grain yield respectively compared to other genotypes (Table 6).
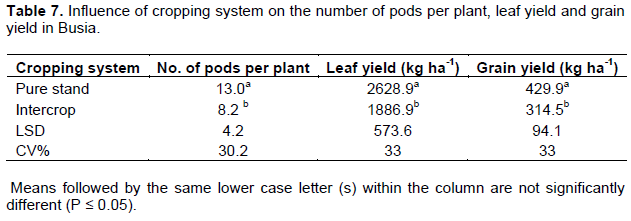
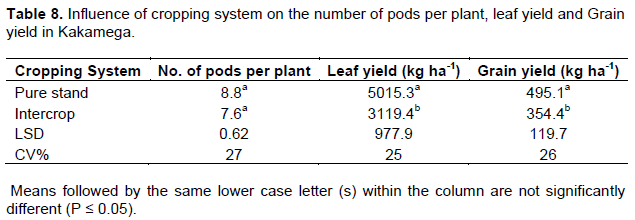
Influence of cropping system on number of pods per plant, leaf and grain yields
The main effect of cropping system on number of pods per plant, leaf and grain yields was significant in both Busia and Kakamega (P ≤ 0.05). In Busia, pure stand cowpea had 58, 39 and 38% more pods per plant, leaf yield and grain yield respectively compared to intercrop cowpea (Table 7). A higher but insignificant number of pods per plant were observed in pure stand cowpea in Kakamega while leaf and grain yields were 60 and 40% respectively more in pure stand cowpea compared to intercrop cowpea (Table 8).
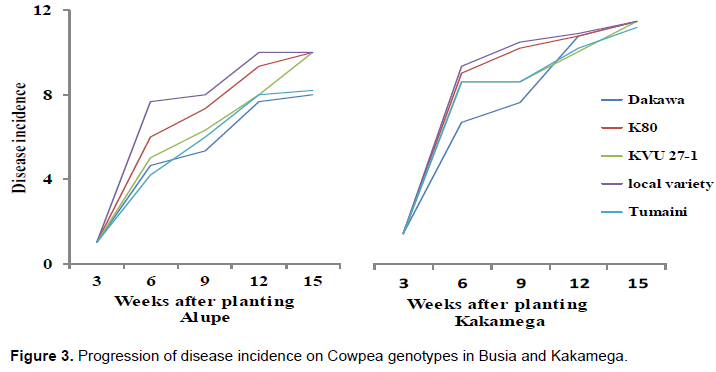
Cowpea rust disease incidence and severity progression over time
Disease incidence (DI)
There was a progressive increase in the intensity of DI with advancement in plant age both in Busia and Kakamega with Dakawa and Tumaini recording lower DI at all growth stages (Figure 3). The intensity of DI tends to peak at week 9 (flowering stage) and levels off for the period of 12-15 weeks (podding and physiological maturity stages).
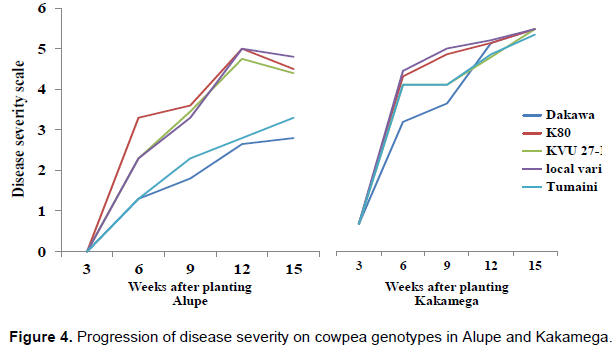
Disease severity (DS)
Disease severity increased with increase in plant age in all the cowpea genotypes in both Busia and Kakamega. Dakawa and Tumaini cowpea genotypes had lower DS at all growth stages compared with the local variety, K80, and KVU 27-1 genotypes (Figure 4). Disease severity rose steadily from vegetative (week 3 and 6), flowering (week 9) to podding (week 12) but slightly dropped at physiological maturity (week 15) in all the genotypes.
DISCUSSION
Cowpea rust was prevalent in both the two varied agroecosystem study locations of Busia and Kakamega. However, the disease was less prevalent in Busia, with low disease incidence and severity than in Kakamega. Though the data are not represented, rainfall was higher and temperature lower in Kakamega than in Busia. The higher rainfall and lower temperatures in Kakamega might have created higher relative humidity (Harrison et al., 1997; Lawrence, 2005) with low light intensities. This resulted in greater accumulation of cowpea rust fungi spores with high intensity of infestation and spread in Kakamega. High relative humidity implied long periods of leaf surface wetness which has been reported to favour the development and sporulation of fungal diseases (Dorrance et al., 2003; Gautam et al., 2013). The results are in line with the findings that environments in humid agro-ecological regions are more conducive for the growth and development of fungal disease-causing agents (Adegbite and Amusa, 2008). According to research done by Adandonon et al. (2003), disease incidence of cowpea stem rot was higher in the south and central zones of Benin Republic than its Northern zone as a result of different amount of rainfall received by the two zones. These results are in agreement with those of Mwang’ombe et al. (2007) who reported low prevalence of Angular Leaf Spot in beans at low altitude regions compared to those planted at high altitude regions.
The disease incidence and severity were lower in Dakawa and Tumaini genotypes and highest in local variety in both Busia and Kakamega. Cowpea genotypes that recorded low disease incidence rates also recorded low disease severity rates. Previous studies have also found a positive correlation between disease incidence and disease severity (Lawrence, 2005). The condition was more pronounced in the cropping system where pure stand cowpea exhibited significantly lower disease incidence rates and subsequently low disease severity scores compared to intercrop cowpea. Although this result contradicts the earlier findings that intercropping combinations result in decreased disease incidence and severity (Ihejirika, 2007; Boudreau, 2013), the intensified high relative humidity due to large plant canopy under intercropping environment has been found to increase fungal spore growth and development (Harrison et al., 1997). This could have resulted in high DI and DS in cowpea intercropped with maize in our study. Additionally, the microclimate created under cowpea-maize intercrop canopy could have reduced wind velocity resulting in decreased air circulation hence prolonged aerial fungal spore accumulation and cowpea leaf wetness favouring cowpea disease incidence and severity. The results are in agreement with the findings of Rothrock et al. (1985) who found that the incidence and severity of Southern stem canker of soybean was more in double cropped soybean/wheat compared to soybean monoculture.
In terms of growth and yields, Dakawa and Tumaini cowpea genotypes had generally higher leaf area index, leaf and grain yields compared to K80, local variety and KVU 27-1. Larger leaves with high LAI are known to increase surface area for photosynthesis and amount of biomass a plant produces (Balemi, 2009). The same trend was reflected in the cropping system where cowpea under pure stand had more number of leaves, more pods per plant and hence higher leaf and grain yield than in the intercrop stand. Intercropping legume with cereal has been found to create micro-climate which favoured disease manifestation and development with concomitant yield reductions in the affected crops (Simbine, 2013). Furthermore, other factors that prevail under intercropping such as minimum exposure to light that is essential for photosynthesis hence low dry matter accumulation, logging on the ground due etiolation and weak stems (Yang et al., 2014) could also limit plant development and productivity. This finding is in agreement with previous research that found out that abiotic and biotic stresses can reduce yield of crops. For example moisture stress has been documented to reduce the yield benefit from narrow row spacing in Kansas by more than 20% (Heitholt et al., 2005). The low disease incidence and severity in pure stand cowpea could have prompted leaf growth and development as a result of proper cell division and elongation in the meristematic tissues hence greater yields.
An exponential progression of disease was observed upto week 9 after planting after which the disease incidence and severity tend to flatten in both Busia and Kakamega experimental sites. The intensity of progression was however low in Dakawa and Tumaini compared with local variety, K80 and KVU 27-1. The low disease incidence at the initial vegetative stage (week 3 and 6) could be attributed to low fungal spores count during disease infection phase and also due to the vigorous growth and high immunity of the cowpea plants fighting the fungus. However, as weeks progress the cowpea rust fungus multiplies faster due to prevailing canopy created microclimate. This finding is in agreement with research conducted by Kone et al. (2017), who found out that as disease progresses it suppresses plasticity and recovery rate of cowpea genotypes. McCain and Hennen (1984) also found that younger leaves are immune to infection due to absence of well-developed stomata. On such leaves, the fungus fails to recognize stomata for infection to take place (Coutinho et al., 1994). Besides, the hydrophobic nature of young leaf surface may hinder fungal spore germination and penetration processes. The significant differences of dsease incidences and severities in cowpea genotypes and growth indicated the existence of varying tolerance and susceptibility among cowpea genotypes with growth period. The finding is in agreement with Schneider et al. (1975), who showed that cowpea genotypes have varying tolarence and susceptibility levels as time progresses upon cowpea rust infestation. This study clearly illustrates that there is variability in cowpea rust disease incidence and severity among the recommended improved cowpea genotypes in western Kenya. The study further provides information on cowpea rust disease prevelence under major cropping systems in two contrasting agroecosystems in western kenya.
CONCLUSIONS
From the current study it is evident that there is significant effect of cowpea genotype and cropping system on disease incidence, severity, and yields. Disease intensity and severity affects growth and yields of the affected crop. The results indicated that Dakawa and Tumaini cowpea genotypes have potential resistance to cowpea rust disease resulting in greater yields. The resistance and performance of these genotypes could be improved by planting them in pure stand. The authors recommend comprehensive investigations to determine the effects of different cowpea-maize combinations patterns on the occurrence and intensity of cowpea rust disease among the promising genotypes.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abukutsa MOA (2007). The Diversity of cultivated African leafy vegetables in three Communities in Western Kenya. African Journal of Food, Agriculture, Nutrition and Development 7(3):1-15. |
|
|
Adandonon A, Aveling TAS, Labuschagne N, Ahohuendo BC (2003). Epidemiology and Biological control of the causal agent of damping-off and stem rot of cowpea in the Oueme valley, Benin. Agronomical Science 6:1-2. |
|
|
Adegbite AA, Amusa AN (2008). The major economic field diseases of cowpea in the humid agro-ecologies of South-western Nigeria. African Journal of Biotechnology 7(25):4705-4712. |
|
|
Agbogidi OM, Ofuoku AU (2005). Response of Sour Soup (Annona muricata L.) to crude oil level, Journal of Sustainable Tropical Agriculture Research 16:98-102. |
|
|
Allen DJ, Buruchara RA, Smithson JB (1998). Diseases of common bean. In Allen, DJ, Lenn JM (Eds.). The Pathology of Food and Pasture Legumes.UK: International Wallingford CAB. pp. 179-265. |
|
|
Balemi T (2009). Effect of phosphorus nutrition on growth of potato genotypes with contrasting Phosphorus efficiency. African Crop Science Journal 17:199-212. |
|
|
Boudreau MA (2013). Diseases in Intercropping Systems. Annual Review of Phytopathology 51:499-519. |
|
|
Coutinho TA, Rijkenberg FHJ, Van Asch MAJ (1994). The effect of leaf age on infection of Coffee genotypes by Hemileia vastatrix. Plant Pathology 43:97-103. |
|
|
Ding XP, Yang M, Huang H, Chuan YR, He XH, Li CY, Zhu YY, Zhu SS (2015). Priming maize resistance by its neighbors: Activating 1,4-benzoxazine-3-ones synthesis and defense gene expression to alleviate leaf disease. Frontiers in Plant Science 6:1-12. |
|
|
Dorrance AE, Kleinhenz MD, Mc Clure SA, Tuttle NT (2003). Temperature, moisture and seed treatment effects on Rhizoctonia solani root rot of soybean. Plant Disease 87:533-538. |
|
|
FAO (2013). The State of Food Security in the World: The multiple dimension of food security. |
|
|
Francis CA (1986). Future perspectives of multiple cropping. In Francis CA (Eds). Multiple Cropping Systems. Response to Organic and Inorganic Nitrogen in Coastal Kenya. Macmillan, New York, pp. 351-370. |
|
|
Gautam HR, Bhardwaj ML, Kumar R (2013). Climate change and its impact on plant diseases. Current Science Journal 12(105):1685-1691. |
|
|
Harrison JG, Searle RJ, Williams NA (1997). Powdery scab disease of potato: Review. Plant Pathology 46:1-25. |
|
|
Henrik S, Peter P (1997). Reduction of stem borer damage by intercropping maize with Cowpea. Agriculture Ecosystem and Environment 62:13-19. |
|
|
Heitholt JJ, Farr JB, Eason R (2005). Planting configuration cultivar effects on Soybean production in low environments. Crop Science 45:1800-1808. |
|
|
Ihejirika GO (2007). Effects of maize intercroppng on Incidence and Serverity of Leafspot Disease (Cercospora arachidicola Hori) and Nodulation of Groundnuts. International Journal of Agricultural Research 2(5)504-507. |
|
|
Jaetzold R, Schmidt H, Horntz B, Shisanya C (2007). Farm management handbook of Kenya Vol II, Natural Conditions and Farm Management Information, Part A Western Kenya, Subpart AI Western Province. 2nd Edition, Ministry of Agriculture, Livestock and Marketing Kenya, in Cooperation with the German Agency for Technical Cooperation (GTZ). |
|
|
KARI (Kenya Agricultural Research Institute) (2000). Soil Fertility Management Handbook for Extension Staff and Farmers in Kenya. KARI Technical Note Series, No. 6:45. |
|
|
Kone N, Asare-Bediako E, Silue S, Kone D, Koita O, Menzel W, Winter S (2017). Influence of planting date on incidence and severity of viral disease on Cucurbits Under field condition. Annals of Agricultural Sciences 62(1):99-104 |
|
|
Lawrence MG (2005). The Relationship between Relative Humidity and the Dew point Temperature in Moist Air: A Simple Conversion and Applications. Bulletin of America Meteorology Society 86:225-233. |
|
|
Mayee CD, Datar VV (1986). Phytopathometry Technical Bulletin-1 (Special bulletin-3), Marathwada Agriculture University, Parbhani, Maharashtra, India, P. 29. |
|
|
McCain JW, Hennen F (1984). Development of uredinial thallus and sorus in the orange Coffee rust fungus Hemileia vastatrix. Phytopathology 74:714-721. |
|
|
Mwang'ombe AN, Magara IN, Kimendu JW and Buruchara RA (2007). Occurrence and Severity of Angular Leaf Spot of Common Beans in Kenya as Influenced by Geographical Location, altitude and Agroecological Zones. Plant Pathology Journal 6(3):235-241. |
|
|
Quin FM (1997). Introduction. In Singh, BB, Mohan Raj DR, Dashiell, IE, Jackai, LEN (Eds.), Advances in cowpea research. Co-publication of International Institute of Tropical Agriculture (IITA) and Japan International Research Center for Agricultural Sciences (JIRCAS), IITA, Ibadan, Nigeria. |
|
|
Rothrock CS, Hobbs TW, Philips DV (1985). Effects of tillage and cropping system on incidence and severity of Southern stem canker of soybean. Phytopathology 75:1156-1159. |
|
|
Saidi M, Itulia FM, Aguyo N, Mushenga PM (2010). Yields and profitability of dual-purposeSole Cowpea and cowpea-maize intercrop as influenced by leaf harvesting. Journal of Agriculture and Biology Science 5(5):65-71. |
|
|
Schneider KA, Brothers ME, Kelly JD (1975). Marker-assisted selection to improve drought Tolerance in common bean. Crop Science 37:51-60. |
|
|
Simbine M (2013). Effect of soybean (Glycine max) planting density on maize (Zea mays) yield in Mumias and Butere Western Kenya. Msc Thesis. |
|
|
Singh BB, Mohan Raj DR, Dashieli KK, Jackai L (1997). Advances in cowpea Research. Co-publication of International Institute of Tropical Agriculture (IITA) and Japan International Research Center for Agricultural Sciences, IITA Ibadan, Nigeria. |
|
|
Mensah TA, Tagoe SMA, Asare ATA, Agyirifo DS (2018). Screening of Cowpea (Vigna unguiculata L. Walp) Genotypes for Rust (Uromyces phaseoli var.vignae) Resistance in Ghana. Plant 6(4):67-74. |
|
|
Vanlauwe B, Coe R, Giller KE (2016). Beyond averages: new approaches to understand heterogeneity and risk of technology success or failure in smallholder farming. Experimental Agriculture 1(1):1-23. |
|
|
Woomer PL, Karanja, N, Kasamuli, SM, Bala A (2011). A revised manual for Rhizobium Methods and standard protocols. |
|
|
Yang F, Huang S, Gao R, Liu W, Yong T, Wang X, Wu X, Yang W (2014). Growth of Soybean seedlings in relay strip intercropping systems in relation to light quantity and Red: far red ratio. Field Crops Research 155:245-253. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


