Full Length Research Paper

ABSTRACT
Plant cocktails used as cover crop present a significant multifunctional resource compared to monoculture, which may increase functional diversity within crop rotation and is particularly effective for designing mixtures with characteristics that increase the multifunctionality and sustainability of the agroecosystem. The objective was to evaluate sustainable agroecosystems models that improve the efficiency of nutrient cycling for crops. The treatments were arranged in four blocks in a split-plot design with two soil management (tillage and no-till) systems and three cropping systems (2 plant cocktails [PCs] and 1 natural vegetation [NV]). C/N ratio, lignin content, dry biomass (DB) production and decomposition, nutrients accumulation and mineralization by PCs and NV were evaluated. Decomposition and release of nutrients were monitored by the litterbag-method. PCs had a DB production twice higher than NV, essential for the adoption of no-tillage systems. The order of nutrient release was K> N> Ca> P> Mg. PCs as cover crops and green manure, with or without predominance of legumes and use of a no-tillage system, could be a technological strategy in agroecosystems for nutrients cycling in semi-arid regions.
Keywords: plant mixture, soil management, mineralization rate, macronutrient, sustainable agriculture.
INTRODUCTION
Nowadays, land use systems that are mostly based along the lines of sustainable development has been soughted, such as adoption of no-tillage system. By means of a global meta-analysis, Pittelkow et al. (2015) observed that the use of a no-till system, with use of cover crop, promotes increased agricultural production compared to conventional systems in arid regions, particularly where water is limiting for crops growth. However, management recommendation for the use of cover crops in semi-arid region is currently limited by the lack of information (Liebig et al., 2015).
The direct influence on improving soil conditions by cover crops is the reduction of nutrient losses by leaching and erosion, maintaining of soil moisture, increasing of soil water infiltration, control of natural vegetation, supply of available nutrients and increasing biological diversity, especially when using leguminous species and improving soil structure when using grasses (Almeida et al., 2016).
Many studies have shown the benefits of the single cultivation of cover crops, but the value of varied cover crop plant cocktails (PC) has received less attention (Finney and Kaye, 2017). Recent recommendations support the use of PC instead of a single species in semi-arid environments, such as the use of leguminous and non-leguminous species, like grasses and oilseeds, to promote mulch stability in no-tillage system, but it still requires more detailed scientific studies (Wutke et al., 2014). The simultaneous cultivation of leguminous and non-leguminous species in the same area can be an effective tool to merge the advantages of individual species (Liu et al., 2017).
However, understanding the residue decomposition dynamic and nutrients immobilization/mineralization in the soil is necessary to identify the most adapted species combination (Xavier et al., 2017). Those species must have characteristics related to climate adaptation and tolerance to drought, biomass production and soil cover efficiency, biological fixation of nitrogen (BFN), decomposition rate and cycling and accumulation of nutrients (Roberts and Mattoo, 2018).
The knowledge of DB production and dynamic of nutrients release from cover crops residues is essential to promote their maximum persistence on the soil surface, which contributes to the maintenance of moisture, soil protection and the success of sustainable production system in a semiarid environment (Giongo et al., 2011). Cover crops with high biomass production and nutrient recycling are essential to maximize the productivity of successional crops in a no-till system (Neto et al., 2011). In this context, Alvarenga et al. (2001)recommended 6.0 Mg ha-1 of dry biomass for obtaining a good soil cover in no-till system in Cerrado (Brazilian Savanna), but the amount of dry biomass can vary according to the type of plant and the edaphoclimatic conditions of the region, which affect the biomass production and the decomposition process.
The evaluation of plant residues decomposition added to the soil by cover crops provides a better understanding of nutrient supply for important commercial crops (Xavier et al., 2017). Typically, species are grouped into two decomposition classes using C/N rate value to separate them in those with rapid decomposition and with slow decomposition rate, which is accepted a C/N value close to 25 as references to separate these groups (Wider and Lang, 1982). Leguminous species, mainly due to its high N content and nutrient cycling, presents rapid decomposition and release of nutrients (Talgre et al., 2017). However, besides C/N values, other factors have an influence on decomposition process, which are lignin and polyphenols contents and their relationships associated with weather, the microorganisms action and soil conditions (Carvalho et al., 2015).
In semi-arid regions, where high temperatures and scarcity of water in drought periods prevail, is acceptable that plant residues decomposition is very low. However, research carried out by litterbag-method in a semi-arid of Enthiopia showed a high plant residue decomposition even when litterbags were placed on the soil surface (Abera et al., 2014).
Several species of cover crops plants can be used to improve the cycling of nutrients in the soil (Isbell et al., 2017). The mix of species, with the use of legumes and non-legumes, should take a biomass production with intermediate C/N, providing soil cover for more time and a better synchronization between nutrients released by cover crops residues and nutritional demand of the commercial crop in succession (de Sá Pereira et al., 2017). The favorable effects of cover crops on soil properties and crop yields are recognized, due to biomass production, nutrient cycling capacity, especially from deeper layers, and subsequent nutrients release by plant residues decomposition (Mendonça et al., 2015).
Multifunctional agriculture is attracting more and more attention among scientists and is considered as a way to obtain sustainability, since it focuses on the complex correlation and interactions between the agroecosystem, natural environment and socioeconomic development (Zhen et al., 2017). Plant cocktails used as cover crop presents a significant multifunctional resource compared to monoculture, which may increase functional diversity within crop rotation, and is particularly effective for designing mixtures with characteristics that increase the multifunctionality and sustainability of the agroecosystem (Blesh, 2017). The design of polycultures that maximize functional diversity may lead to agroecosystems with greater multifunctionality (Finney and Kaye, 2017).
A practical method was assessed with the use of mixed cropping as cover crops and green manure as a technological strategy for nutrients cycling in irrigated systems in semiarid regions. However, few are the studies about the dynamics of nutrients and information on the effect of mixed cropped as green manures on the nutrient cycling (Tarui et al., 2013). This study presents the nutrient cycling potential of multifunctional Agroecosystems when plant mixtures are cultivated before melon crop produced in irrigated areas from semiarid. It provides an approach to identify the critical processes and opportunities for improvements in the conventional crop system that may support producers in the task to improve soil quality and reduce cost with fertilizers.
The objective was to evaluate sustainable agroecosystems models that improve the efficiency of nutrient cycling for crops.
MATERIALS AND METHODS
Area descriptions
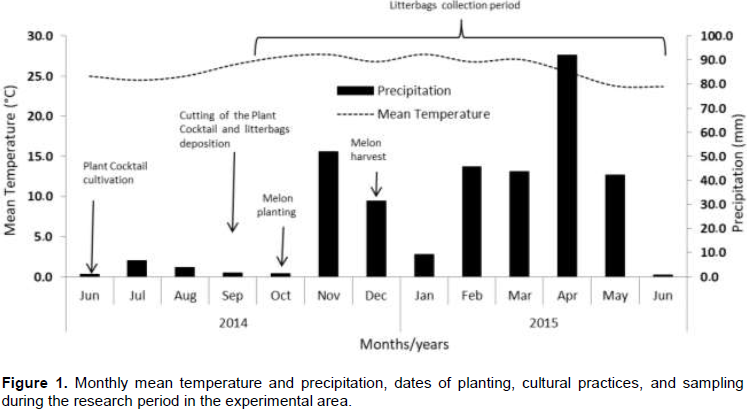
The study was conducted in an experimental field located at geographical coordinates 9°08’S, 40°18’W, 365.5 m a.s.l from June 2014 to June 2015 in the initial phase of a long-term experiment. The soil is classified as ultisol eutrophic red-yellow plinthic (EMBRAPA, 2018), with the following physical and chemical characteristics at the depth of 0 to 0.20 m: 83.1% of sand; 11.9% of silt; 4.9% of clay; pH (H2O) 6.6; organic matter 5.82 g dm-3 (wet oxidation-diffusion); P 47.34 mg dm-3 (Mehlich-1); H+Al 1.21 mmol dm-3 (KCl-extraction); K (Mehlich-1); Ca (KCl-extraction); Mg (KCl-extraction); and Na (Mehlich-1) exchangeable, 11.0, 18.5, 7.8, and 0.7 mmol dm-3, respectively, the sum of bases (S) 31.8 mmol dm-3; capacity cation exchange (CEC) 43.4 mmol dm-3 and base saturation (V) 71.6% (EMBRAPA, 2011). The climate is classified as BSwh’ according to the Köppen classification system, with an average annual temperature of 26.8°C, the average annual rainfall of 360 mm, and the climax vegetation called Caatinga (xeric shrubland and thorn forest). Data of mean temperature and precipitation were measured at the agrometeorological weather station located at experimental farm (Figure 1).
Experimental design and plant
The PCs cultivation preceded a yellow melon crop (Cucumis melo L.). The successor culture planted on the PCs and NV residues. The treatments were arranged in four blocks in a split-plot design. Two tillage treatments as main plots had dimensions of 30 × 10 m. Conventional tillage (T) comprised plowing and disking compared to no soil disturbance in NT plots. Sub-plots treatments, 10 × 10 m, comprised three cropping systems, two different compositions of plant cocktail and one natural vegetation cover.
Mixes of plants used as cover crops present greater multifunctionality than monocultures. The greater the diversity of cover crops, the greater the services from agroecosystems, mainly due to the specific ecological characteristics inherent to each species used (Blesh, 2017). Different from other studies, which use few species as cover crops, this work contemplated a larger number of plants to compose the plant cocktail, with potential already known in semiarid region. The PCs composition used with conventional tillage and no-till treatments are: PC1 - 75% legumes + 25% non-legumes; PC2 - 25% of legumes + 75% non-legumes; and NV - natural vegetation. Fourteen species included in the composition of PCs, comprised legumes, oilseeds and grasses, including the following species: (A) Legumes - calopo (Calopogonium mucunoide), velvet bean (Stizolobium aterrimum L.), grey-seeded mucuna (Stizolobium cinereum Piper and Tracy), Crotalaria (Crotalaria juncea), rattlebox (Crotalaria spectabilis), jack beans (Canavalia ensiformes), pigeon pea (Cajanus cajan L.), lab-lab bean (Dolichos lablab L.); (B) no legumes: sesame (Sesamum indicum L.), corn (Zea mays), pearl millet (Pennisetum americanum L.), milo (Sorghum vulgare Pers.), sunflower (Helianthus annuus), andcastor oil plant (Ricinus communis L). The natural vegetation composed of the predominant species: benghal dayflower (Commelina benghalensis L.), purple bush-bean (Macroptilium atropurpureum), florida beggarweed (Desmodium tortuosum) and goat’s head (Acanthorpermun hispidum DC).
Data collection
Plant cocktails were cultivated in the second half of June (Figure 1) .The seeds were sown in furrows at a spacing of 0.50 m. The irrigation system used was plastic pipes distributed between the rows with drip emitters spaced at 0.5 m and a flow rate of 4.0 L h-1. In order to ensure the seed germination, initially the smaller seeds were distributed, followed by the intermediate size and then the larger, avoiding the segregation effect. Seventy days after sowing, end of September , when most species were in the flowering stage, the PCs species were cut in the spacing of 1 m², with three replicates for each treatment, to evaluate the total aerial biomass production. PC effective as a cover crop was maintained and the other parts were incorporated by a disc harrow to 0.4 m depth. Subsamples of plant cocktails from each treatment were weighted and sent to the Laboratory of Soil (Embrapa - semiarid), stored in a greenhouse at 65 to 70°C for 72 h, and weight again (g kg-1) to estimate the dry matter yield (t ha-1), lignin content (g kg-1) and C/N ratio. Natural vegetation, which grows spontaneously in the area, had a similar procedure for evaluation.
For analysis of residues decomposition and nutrient release by PCs and NV were used for only three experimental blocks. Samples of the aerial biomass of plants cocktails and natural vegetation were used for the confection of litterbags (size 0.3 × 0.3 m, mesh 0.5 mm) with 250 g of wet biomass. There is a clear difference in decomposition rates between shoots and roots, which may also be explained by the differences in the chemical composition of the residue (Talgre et al., 2017). Thus, shoots and roots from the different plants were cut and mixed. According to the treatments design, the litterbags were buried in 0.2 m soil depth or deposited on the soil surface. Only one block was used for litterbags deposition, which was arranged in rows (between the melon crop lines) and identified by stakes with nameplates. After plant residues deposition and during the first 3 months (melon culture lifespan), drip irrigation system was used with drip emitters distributed on the rows with 2.0 m width. For each treatment type, 30 litterbags were used. Sampling was carried out at 0, 14, 21, 50, 78, 105, 141, 172, 208, 236, and 258 days after deposition (DAD) (Figure 1) , which 3 litterbags from each treatment were removed from the field. The biomass of the litterbags was cleaned manually, by removing the plant roots which have grown into the inside of the bags, dried at 65°C in a forced air oven until a constant weight and weighed. To eliminate soil contamination, the technique of determining the ash residues suggested by Potthoff and Loftfield (1998) was used. Samples of dried biomass were ground in a stainless steel mill type "Wiley" with 1.0 mm mesh sieve for analysis of nitrogen (N), phosphorus (P), potassium (K), sodium (Na), calcium (Ca) and magnesium (Mg) and total carbon.
For analysis of total C and N was used for the Elemental Analyzer Truspec (LECO USA). P and K were determined by nitric/perchloric acid digestion (Bataglia et al., 1983). The P determination was done colorimetrically by forming the blue-colored complex from molybdate in the presence of ascorbic acid and K, by flame photometry (EMBRAPA, 2011). Ca and Mg were analyzed by atomic absorption spectrophotometry (Bataglia et al., 1983). The lignin content was determined by the acid detergent fiber method (Van Soest and Wine, 1968).
The obtained values were converted into percentage concerning the mass and nutrient content of the beginning of decomposition (To). With those data, the biomass decomposition rate and nutrients release for each treatment were determined. The residue decomposition and nutrient release follow the simple exponential model used by Olson (1963):

where 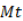 are the remaining percentages of DB and macronutrients after T days and
are the remaining percentages of DB and macronutrients after T days and 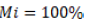 when T is equal to zero, that is, the deposition day of the residues on the soil. With decomposition, constant k was calculated with the time required to release 50%
when T is equal to zero, that is, the deposition day of the residues on the soil. With decomposition, constant k was calculated with the time required to release 50% 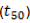 and 95%
and 95% 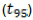 of the nutrients,
of the nutrients,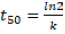 and
and 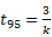 (Shanks and Olson, 1961). Reorganizing the equation, it was possible to calculate the decomposition constant, or -value:
(Shanks and Olson, 1961). Reorganizing the equation, it was possible to calculate the decomposition constant, or -value:

Statistical analysis
The results were statistically analyzed for variance (ANOVA), using the SISVAR® Statistical Software (Ferreira, 2011). The difference between treatment means was assessed by the Tukey test, at 5% probability.
RESULTS AND DISCUSSION
Residues production, content, and accumulation of macronutrients
In a previous work carried out in the same area showed that PCs do not significantly differ among themselves, with mean value of 8.3 t ha-1 for PC1 and 9.20 t ha-1 for PC2 composition, and NV with a DB representing about 50% of the PCs with 4.50 t ha-1 (Giongo et al., 2016).
The biomass production and nutrients accumulation in 2014 by PC1, PC2, and NV are shown in Table 1. Despite not significant, the PC1, with the highest percentage of legumes, showed the highest biomass production (8.86 t ha-1). The NV presented almost 50% less DB production (4.35 t ha-1) than PC1 and PC2. However, the PC2 DB (7.55 t ha-1) was not significantly different from PC1 DB, may be related to the high photosynthetic capacity and carbon-richness biomass of the grasses and the nutrient cycling efficiency of the oilseed plants as well (Giacomini et al., 2003)
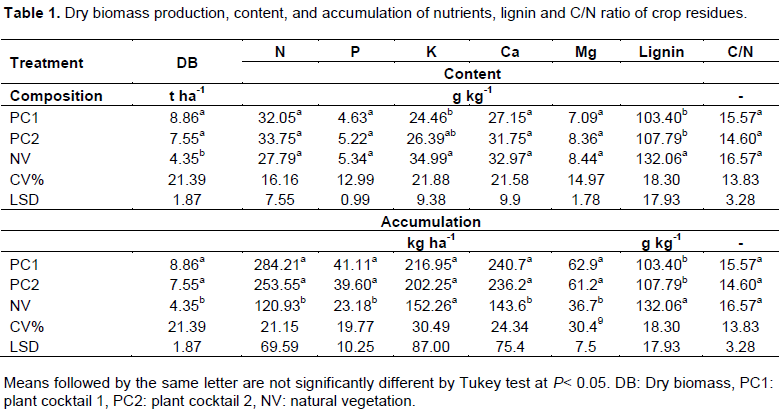
In relation to the quantity of residues required to maintain soil protection, the DB production by CPs was higher than the values suggested by Alvarenga et al. (2001), that is, 6.0 t ha-1. In semiarid multifunctional agroecosystems to increase the half-life, to promote the gradual release of the nutrients and the time of cover of the soil it is fundamental that the biomass is not incorporated (Blanco-Canqui et al., 2015).
The DB production can vary significantly with rainfall index, temperature, location and soil fertility (Torres and Pereira, 2014). In relation to NV, must be considered that it holds a varied seed bank, and diverges with soil management, availability of rainfall and photoperiod that could reflect directly on the annual DB production (Borgy et al., 2015).
The plant residues contents of N, P, Ca, and Mg did not differ significantly among the different cover crops tested. Although the content of K was higher in NV, the amount of this nutrient was lower due to the low DB production in comparison to the PCs (Table 1).
With regard to accumulation of nutrients, with exception of K, PC1 and PC2 differ significantly (P <0.05) from NV on the amount of these elements added to the soil. Although it was not statistically significant, the PC1, with a higher proportion of legumes, presented an accumulation slightly larger for all elements. Thus, for PC1 the values of N, P, K, Ca, and Mg reached 284.21, 41.11, 216.95, 240.7, and 62.9 kg ha-1, respectively.
Therefore, the N was the nutrient with the highest accumulation followed by K, different from NV that accumulated more K than N. Hence, the plant mixture cultivated PC1 and PC2 reached the follow accumulation order N > Ca > K > Mg > P while NV got the follow order K > Ca > N > Mg > P (Table 1). The presence of oilseeds could have contributed to N accumulation from PC2 which is not different significantly from PC1 (Liu et al., 2018). Both plant mixtures (PC1 and PC2) and spontaneous vegetation (NV) were efficient in K uptake and cycling, indicating that cover crop cultivation is an important strategy to mitigate losses of K by leaching and erosion. It is possible to observe that the performance of NV in nutrient uptake in relation to plant mixtures (PC1 and PC2), without considering phytomass production, was very efficient in nutrient cycling, since there were no significant differences between them.
The amount of nutrient uptake depends on the species, soil fertility, phenological stage in dehydrating the C/N ratio, the planting season, besides the climatic conditions of each study (Fontana et al., 2018).
Decomposition rate of plant residues
The study of plant residues decomposition rate allows checking when and how much of each nutrient is released in the soil , consequently to promote the adjustment of fertilization. Table 2 shows the decomposition constant , the 50 and 95% decomposition of plant residues. The soil management system changed the decomposition rate of PC and NV. In no-till treatments, the absence of plowing promoted a decrease of the plant residue decomposition kinetics and the maintenance of soil cover for more time. The decomposition of 50% of PCs DB takes approximately 115 days when is not incorporated (NT) in the soil, and around 98 days when is incorporated (T), showing a small difference between the types of soil management. To decompose 95% of the PCs and NV, DB takes around 497 and 424 days, respectively, in NT, and 400 and 500 days, respectively, in T. Although not high, there was a difference in residues decomposition from the surface deposited litterbags than those that were buried. In tropical soils, the higher amplitude of diurnal temperature variation results in higher decomposition than under constant soil temperature condition (Séna Koglo et al., 2017). Unfortunately, we did not record the soil temperature, but high mean air temperature was registered (>25°C) in most months of this experiment (Figure 1). However, differences in soil moisture content probably may also be favoring the decomposition process. Previous studies in the same area showed that soil moisture content in 0.2 m depth was higher in all treatments with PCs under NT than T (Pereira Filho et al., 2016).
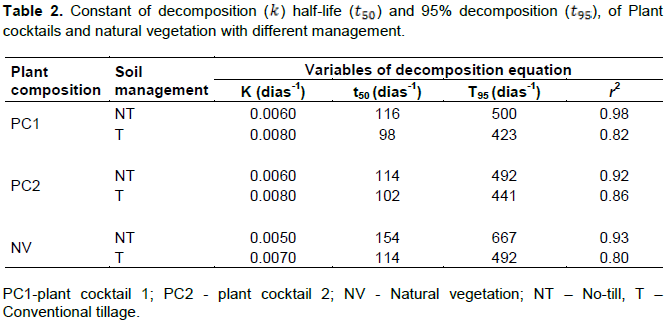
C/N values showed no significant differences in all treatments. In addition, those values presented below of the threshold that separate the faster and slower decomposition group (C/N = 25), certainly related to the promotion of intermediate values by PCs. Combined with C/N values, highlight the lignin contents that were significantly higher in NV (p>0.01) than PCs. Lignin inhibits the decomposition of plant residues, accentuating the maintenance of soil cover (Xia et al., 2017). NV had a lower decomposition rate with the half-life of 154 days when not incorporated and 114 days when incorporated in the soil. This can be explained by the fact that NV has higher values of both C/N ratio (16.57) and lignin content (132.06 g kg-1) becoming more resistant to decomposition. In addition, the smaller the N value and the higher the lignin value, the lower is the decomposition rate (Xia et al., 2017). Consequently, C/N and lignin values corroborated with t50 and t95 values found.
The dynamics and decreasing loss of relative dry biomass by PCs and NV are as shown in Figure 2. Based on the exponential model equation, the plant residues presented the relative loss of dry matter in the end evaluated period: PC1, PC2, and NV decomposed 78.73, 79.23 and 68.57%, respectively, when not incorporated and 83.91, 82.71 and 79.25%, respectively, when incorporated in the soil .The kinetics of the decomposition process showed a similar pattern in all treatments, with a fast initial phase followed by a slower one. The initial and rapid phase is related to leaves and other materials less lignified, and the next and slower phase is related to the decomposing of more resistant compounds (Xavier et al., 2017). In addition, the use of drip irrigation during the successor crop life cycle may promote an increase in the decomposition process This result was also observed by Giongo et al. (2011)in previous studies in the same area.
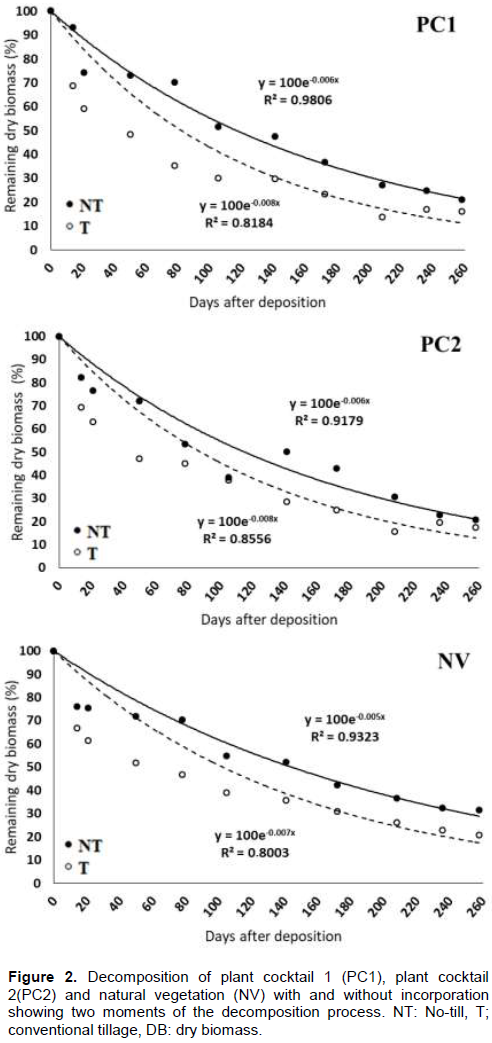
It was observed that the quantity of biomass produced by PCs did not decompose completely during the study period, essentially for not incorporated treatments, with higher DB quantity compared with values recommended by Alvarenga et al. (2001)for Cerrado regions (Brazilian Savanna) where the rainfall indices are higher than semi-arid regions. It is important that PCs produce a satisfactory amount of mulch for adoption, maintenance or continuation of similar production systems in the region. In semi-arid multifunctional agroecosystems, it is fundamental that the biomass is not incorporated into the soil in order that it increases its half-life, promote the gradual nutrients release and increase the time of soil cover. In addition, the PCs have been demonstrated to be alternatives to supply part of the successional crop nutritional demand, hence contributing to the reduction of production cost by reducing the quantity of fertilizers.
Nutrients released from plant residues
The intensity and duration of rainfall are factors that influence the biomass decomposition and nutrient release (Mendonça et al., 2015). During the study period, with low monthly rainfall averages, typical of the study area, were less than 50 mm, except for April 2015 with 90 mm (Figure 1). The low rainfall observed may not have influenced the nutrients release from residues, considering the presence of water in the soil, due to irrigation during the experiment, and also the overlapped wet bulbs observed.
Table 3 shows the analysis of variance among the types of plant cocktails and types of soil management with regard to decomposition constant k and the release time of 50 and 95% of nutrients by plant residues. It was observed that the treatments with soil tillage there was a higher rate of decomposition, consequently leading to less time of nutrients release. In treatments with soil tilling no significant difference occurred between the soil cover types for k and time of nutrient release.
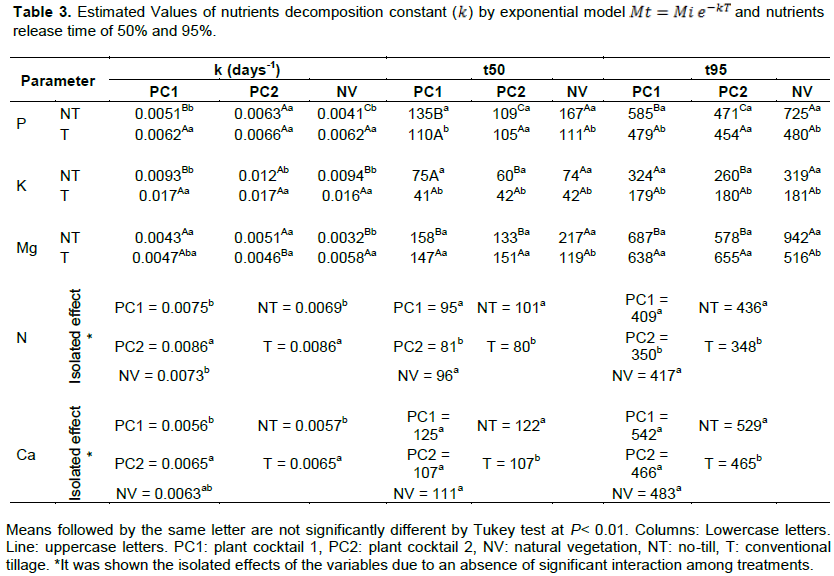
The results of analysis of variance showed significant results (p>0.01) for the variables time, cover crop type and the interaction of time/cover crop type. The kinetics of nutrients mineralization process in all treatments showed a similar pattern, with an initial rapid phase during the first months, when it was released great part of nutrients mainly by incorporated treatments, and followed by a slower release phase.
N is one of the most affected nutrient by no-till system, since the maintenance of residues on the soil surface modifies the processes of immobilization, mineralization, and leaching (Zhang et al., 2016), promoting the reduction of residues decomposition process and causing changes in the availability and loss of N (Frasier et al., 2017). The N mineralization occurred differently between incorporated and non-incorporated treatments, showing a slightly faster release in conventional tillage.
No significant interaction was observed among the types of treatments (Table 3). However, when evaluated separately, the types of plant cocktails and soil management showed a significant interaction for the Tukey test at 5% probability (p<0.05). In relation to the constant of decomposition (k), plant cocktail 2 with a mean of 0.0086 day -1 and treatments with T also with a mean of 0.0086 day-1 were highlighted. PC1 and NV take longer time to release 50% of nitrogen than PC2, with means of 95 days ( =0.0075 day-1) and 96 days ( =0.0073 day-1), respectively. As expected, the treatments in T system were those who take less time (80 days; =0.0086 day-1) to release half of the amount of N from the residues dry mass. To release 95% of the nitrogen from the residues it takes between 350 to 417 days, depending on the plant composition and soil management type (Table 3). Half of the nitrogen is released in the first 3 months after cutting the plant mixtures used as a cover crop. The amount of N released quickly in the early decomposition stage is associated with loss of water-soluble fractions of this nutrient (Zhang et al., 2016). Although the loss of N occurs by volatilization, leaching, etc., the amount of N that returns to the soil in the form of plant residues represents a considerable portion of the total N absorbed by plants in succession (Costa et al., 2012).
The cover crops cultivation promotes the increase of microbial P and labile P forms, in addition, the slow and gradual residue mineralization in the no-till system causes the release and redistribution of organic P forms, more stable and less susceptible to adsorption reactions (Malhi et al., 2018). Different from T system, the P decomposition rates (k) and times of 50 and 95% nutrients release were significantly different between PCs and NV (P<0.01) in NT. To release 50% of P in NT could range from 109 to 135 days for PCs and 167 days for NV. Treatments in T could range from 105 to 111 days to release 50% of P. The release of 95% of the P can take about 725 days (k = 0.0041 day-1) in NT and 480 DAD (k = 0.0062 day-1) in T for NV. Among the plant cocktails, the NPC1 takes 585 days in NT and 479 days in T to release 95% of P (Table 3).
The quantity of P released by NV, which was close to half of P accumulated by PC, is directly related to the average amount of this nutrient accumulated by NV biomass (Table 1).
The higher release of P during the initial period of decomposition is related to the loss of soluble P accumulated in the plant tissues vacuoles (Yang et al., 2017). Usually, the intensity and duration of rain are responsible for the amount of P that returns to the soil from the crop residues (Costa et al., 2012), but the phosphorus released have a tendency to be associated with the irrigation process than rainwater due to the incidence of low rainfall during this period.
Among the nutrients, K was the faster-released nutrient by plant residues, mainly in T system when the dry biomass residues were incorporated into the soil, which presented significant differences between the types of soil management (p<0.001). However, there were no significant differences among the treatments in T related to decomposition rates (k) and times of 50 and 95% of nutrients release (p <0.001). The residues required between 60 and 75 days ( =0.0093 day-1) in NT treatments and around 42 days ( =0.017 day-1) (for all the treatments) in T to release 50% of K. 95% of K was released between 260 and 324 days in NT treatments and between 179 and 181 DAD in T treatments (Table 3).
The highest speed of K release from plant residues can be attributed to the fact that K is an element that is not associated with any structural component of vegetal tissue, and found in ionic form with easily extracted from the plant tissue (Karthika et al., 2018)without necessarily having a biological decomposition and mineralization (Bernardes et al., 2010). Consequently, it is essential to reduce the implantation period of the successor culture to minimize the loss of K.
Different from the K, Ca has more difficulties to be mineralized by the fact that it makes part of the middle lamella of the cell wall constituents, forming one of the most recalcitrant components of plant tissues, and cofactor of some enzymes involved in the hydrolysis of ATP and phospholipids, and also secondary messenger in metabolic regulation (Karthika et al., 2018). Like N, no significant difference was observed among the treatments (Table 3). However, when evaluated separately, the types of plant cocktails and soil management showed a significant interaction for the Tukey test at 5% probability (p<0.005). To release 50% of Ca from dry biomass residues, the different cover crop could take 111 to 125 days. The treatments with NT took more time (122 days; = 0.0057 day-1) than T (107 days = 0.0065 day-1) to release half of the amount of Ca from the residues dry biomass. To release 95% the plant mixtures (PCs and NV) residues could take a mean of 500 days (Table 3). On average, like P and N, plant residues takes approximately 3 months to release half of the amount of Ca accumulated. The fast release of Ca at the beginning of the process is related to the participation of this element with ionic compounds and water-soluble molecules (de Freitas et al., 2015).
Both 50 and 95% of Mg release from PCs residues were not significantly different between NT and T systems , except for NV (P<0.01) (Table 3). To release 50% of Mg, the PCs treatment could range from 133 (PC2, k = 0.0051 day-1) to 158 days (PC1, k = 0.0043 day-1). However, in NV, which was significantly different between soil management systems , the plant residues to release 50% of Mg could take 119 days (k = 0.0058 day-1) in T and 217 days (k = 0.0032 day-1) in NT. To release 95% of Mg could take about 942 days (k = 0.0032 day-1) for NV in NT and 687 days (k = 0.0043 day-1) for PC1 in NT. Similar to Ca, the higher Mg release early in the process is due to the participation of this element in ionic compounds and soluble molecules (Gransee and Führs, 2013).
NT treatments were significantly different from T treatments (P<0.01) and showed that the incorporation of plant residues increased decomposition and as a result the faster release of nutrients. To release 95% of nutrients from plant residues could take more than a year, except for K which could take about 324 days ( =0.0093 day-1) when not incorporated in the soil, but 181 days ( =0.016 day-1) when incorporated. This suggests that plant residues are still decomposed and nutrients are released along with the residue of the subsequent PC culture.
The order of release 50% of the nutrients accumulated in Plant Cocktails residues was K> N> P > Ca > Mg. 50% of the amount of N, P, Ca and Mg nutrients are released from the residues between the third to the fourth month after plant cocktails deposition, while K may take only 2 months to release into the soil. A management of successor culture can be done knowing the release time of the nutrients by the residues of the cover plants. The use of PCs could replace largely the need to supply nutrients via chemical fertilization, especially when it promotes synchronization between the nutrient released by plants residues with the demand of the successional crop.
Therefore, the PC1 and PC2 could be indicated for use as cover crops and green manure in similar agricultural areas, which demonstrated high potential to produce dry biomass and nutrients accumulation, different from NV that had lower dry biomass production that promoted less soil protection. In a no-till system, those plant mixture could minimize nutrient leakage and maximize nutrient cycling, interfaced with integrated weed and pest management options that require less agrochemicals (Rockström et al., 2017).
The lack of significant differences related to nutrient accumulation between PC1 and PC2 may be linked to the complexity of species selection to compose of the PC and the success depends heavily on interactions between components species, management practices available and environmental conditions (Lithourgidis et al., 2011). Tropical grasses usually have higher photosynthetic ability to grow and produce biomass (Taiz and Zeiger, 2013)whereas legumes can have relatively slower growth, however, with N incorporation function to the production system that may result in plants dominated by the slower species (Kaye and Quemada, 2017), disfavoring the balance in the green manure composition and damaging the desired benefits (Calvo et al., 2010).
The use of any evaluated PC is more efficient in nutrient accumulation and subsequent release into the soil than NV, considering that the amount of nutrient uptake by PC1 and PC2 showed no statistically significant differences. However, it can not disregard the potential of NV to accumulate nutrients and its recalcitrant character, which have the potential to cycle nutrients and maintain soil cover and that need to be better studied.
Cash crops in semi-arid regions, as melon, are characterized by the intense use of agricultural inputs, especially synthetic fertilizers which increase the cost production and environmental impacts. Therefore, PC used as green manure intercropped with melon can improve melon yield and soil quality, by nutrient cycling and biological nitrogen fixation. Thus, they were identified as positive effects in delivered nutrient to the melon when different leguminous and grass plants were cultivated as green manure, before melon production.
Accordingly, the cash crop intercropped with PCs as cover crops and green manure, with or without predominance of legumes and use of a no-tillage system, could be a technological strategy for adding biomass and nutrients to the soil, as well as increasing crop yields in semi-arid regions.
CONCLUSIONS
The data presented support the following conclusions:
(1) PCs had a DB production twice higher than NV, essential for adoption of NT in semi-arid region;
(2) There was a difference in residues decomposition from the surface deposited litterbags than those that were buried ;
(3) DB production and nutrient accumulation by PC1 and PC2 had no significant differences;
(4) Nitrogen was the nutrient with the highest accumulation by PCs followed by K;
(5) The order of release 50% of the nutrients accumulated in PCs and NV residues was K> N> P > Ca > Mg and occurred less than the first four months;
(6) PCs could be indicated for use as cover crops and green manure in similar agricultural areas, which demonstrated high potential to produce dry biomass and nutrients accumulation and they could replace largely the need to supply nutrients via chemical fertilization;
(7) Use of PCs as cover crops and green manure, with or without predominance of legumes and use of a no-tillage system, could bea technological strategy in agroecosystems for nutrients cycling in semi-arid regions.
ACKNOWLEDGEMENTS
The first author cordially acknowledges the National Council for Scientific and Technological Development (CNPq) for granting scholarship. He also is grateful to Empresa Brasileira de Pesquisa Agropecuária (Embrapa Semi-arid) for funding this research via "Strategies for reducing carbon emissions and efficient use of water in irrigated and rainfed production systems in the Brazilian semi-arid” project and provide laboratory facilities and experimental field.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abera G, Wolde-Meskel E, Bakken LR (2014). Unexpected high decomposition of legume residues in dry season soils from tropical coffee plantations and crop lands. Agronomy for Sustainable Development 34:667–676. |
|
|
Almeida DO, Bayer C, Almeida HC (2016). Fauna e atributos microbiológicos de um Argissolo sob sistemas de cobertura no Sul do Brasil. Pesquisa Agropecuária Brasileira 51:1140–1147. |
|
|
Alvarenga RC, Cabezas WAL, Cruz JC, Santana DP (2001) Plantas de Cobertura de Solo para Sistema Plantio Direto. In: Informe Agropecuário. Embrapa Milho e Sorgo, Belo Horizonte, MG. Brasil 22(208):25-36 |
|
|
Bataglia OC, Furlani AMC, Teixeira JPF, Gallo JR (1983). Métodos de análises química de plantas. Instituto Agronômico de Campinas, IAC, Boletim técnico, 78, Campinas-SP |
|
|
Bernardes TG, da Silveira PM, Mesquita MAM, de Aguiar RA, Mesquita GM (2010). Decomposição da biomassa e liberação de nutrientes dos capins braquiária e mombaça, em condições de cerrado. Pesquisa Agropecuária Tropical 40:370-377. |
|
|
Blanco-Canqui H, Shaver TM, Lindquist JL, Elmore RW, Francis CA (2015). Cover Crops and Ecosystem Services: Insights from Studies in Temperate Soils. Agronomy Journal 107:2449-2474. |
|
|
Blesh J (2017). Functional traits in cover crop mixtures: Biological nitrogen fixation and multifunctionality. Journal of Applied Ecology 55:38-48. |
|
|
Borgy B, Reboud X, Peyrard N, Sabbadin R, Gaba S (2015). Dynamics of Weeds in the Soil Seed Bank: A Hidden Markov Model to Estimate Life History Traits from Standing Plant Time Series. PLoS One 10:e0139278. |
|
|
Calvo LC, Foloni JSS, Brancalião SR (2010). Produtividade de fitomassa e relação C/N de monocultivos e consórcios de guandú-anão, milheto e sorgo em três épocas de corte. Bragantia 69:77-86. |
|
|
Carvalho AM De, Coser TR, Rein TA, Dantas RA, Rafael RS, Kleberson WS (2015). Manejo de plantas de cobertura na floração e na maturação fisiológica e seu efeito na produtividade do milho. Pesquisa Agropecuária Brasileira 50:551-561. |
|
|
Costa NR, Andreotti M, de Aquino Gameiro R, Pariz CM, Buzetti S, Lopes KS (2012). Adubação nitrogenada no consórcio de milho com duas espécies de braquiária em sistema plantio direto. Pesquisa Agropecuária Brasileira 47:1038-1047. |
|
|
de Freitas ST, Handa AK, Wu Q, Park S, Mitcham EJ (2015). Role of pectin methylesterases in cellular calcium distribution and blossom-end rot development in tomato fruit. The Plant Journal 71:824-835. |
|
|
de Sá Pereira E, Duval ME, Galantini JA (2017). Decomposition from legume and non-legume crop residues: Effects on soil organic carbon fractions under controlled conditions. Spanish Journal of Soil Science 7:86-96. |
|
|
EMBRAPA (2018). Sistema brasileiro de classificação de solos, 5th edn. Embrapa Soils, Brasilia, DF. Brasil |
|
|
EMBRAPA (2011). Manual de métodos de análises de solos, 2nd edn. Embrapa Solos - Dados eletrônicos, Rio de Janeiro, Brasil 230 p. |
|
|
Ferreira DF (2011). Sisvar: a computer statistical analysis system. Ciência e Agrotecnologia 35:1039-1042. |
|
|
Finney DM, Kaye JP (2017). Functional diversity in cover crop polycultures increases multifunctionality of an agricultural system. Journal of Applied Ecology 54(2):509-517. |
|
|
Fontana M, Labrecque M, Messier C, Bélanger N (2018). Permanent site characteristics exert a larger influence than atmospheric conditions on leaf mass, foliar nutrients and ultimately above ground biomass productivity of Salix miyabeana 'SX67.' Forest Ecology and Management 427:423-433. |
|
|
Frasier I, Noellemeyer E, Amiotti N, Quiroga A (2017). Vetch-rye biculture is a sustainable alternative for enhanced nitrogen availability and low leaching losses in a no-till cover crop system. Field Crops Research 214:104-112. |
|
|
Giacomini SJ, Aita C, Vendruscolo ERO, Cubilla M, Nicoloso RS, Fries MR (2003). Matéria seca, relação C/N e acúmulo de nitrogênio, fósforo e potássio em misturas de plantas de cobertura de solo. Revista Brasileira de Ciência do Solo 27:325-334. |
|
|
Giongo V, Mendes AMS, Cunha JTF, Galvão SRS (2011). Decomposição e liberação de nutrientes de coquetéis vegetais para utilização no Semiárido brasileiro. Revista Ciência Agronômica 42:611-618. |
|
|
Giongo V, Salviano AM, Santana MS, Costa ND (2016). Soil management systems for sustainable melon cropping in the submedian of the são francisco valley. Revista Caatinga 29:537-547. |
|
|
Gransee A, Führs H (2013). Magnesium mobility in soils as a challenge for soil and plant analysis, magnesium fertilization and root uptake under adverse growth conditions. Plant Soil 368:5-21. |
|
|
Isbell F, Adler PR, Eisenhauer N, Fornara D, Kimmel K, Kremen C, Letourneau D, Liebman M, Polley HW, Quijas S, Scherer-Lorenzen M (2017). Benefits of increasing plant diversity in sustainable agroecosystems. Journal of Ecology 105:871-879. |
|
|
Karthika KS, Rashmi I, Parvathi MS (2018). Biological Functions, Uptake and Transport of Essential Nutrients in Relation to Plant Growth. In: Hasanuzzaman M, Fujita M, Oku H, Nahar K, Hawrylak-Nowak B. (eds) Plant Nutrients and Abiotic Stress Tolerance. Springer Singapore, Singapore pp. 1-49. |
|
|
Kaye JP, Quemada M (2017). Using cover crops to mitigate and adapt to climate change. A review. Agronomy for Sustainable Development 37:4. |
|
|
Liebig MA, Hendrickson JR, Archer DW, Scholljegerdes EJ, Hendrickson JR, Tanaka DL (2015). Short-Term Soil Responses to Late-Seeded Cover Crops in a Semi-Arid Environment. Agronomy Journal |
|
|
Lithourgidis AS, Dordas CA, Damalas CA, Vlachostergios DN (2011). Annual intercrops: An alternative pathway for sustainable agriculture. Australian Journal of Crop Science 5:396-410. |
|
|
Liu T, Ren T, White PJ, Cong R, Lu J (2018). Storage nitrogen co-ordinates leaf expansion and photosynthetic capacity in winter oilseed rape. Journal of Experimental Botany 69:2995-3007. |
|
|
Liu Y, Tian F, Jia P, Zhang J, Hou F, Wu G (2017) Leguminous species sequester more carbon than gramineous species in cultivated grasslands of a semi-arid area. Solid Earth 8(1):83-91. |
|
|
Malhi SS, Légère A, Vanasse A, Parent G (2018). Effects of long-term tillage, terminating no-till and cropping system on organic C and N, and available nutrients in a Gleysolic soil in Québec, Canada. The Journal of Agricultural Science 156:472-480. |
|
|
Mendonça VZ De, Mello LMM De, Andreotti M, Pariz CM, Yano EH, Pereira FCBL (2015). Liberação de nutrientes da palhada de forrageiras consorciadas com milho e sucessão com soja. Rev Bras Ciência do Solo 39:183-193. |
|
|
Neto JF, Alexandre C, Crusciol C, et al (2011). Plantas de cobertura, manejo da palhada e produtividade da mamoneira no sistema plantio direto. Revista Ciência Agronômica 42:978-985. |
|
|
Olson JS (1963). Energy Storage and the Balance of Producers and Decomposers in Ecological Systems. Ecology 44:322-331. |
|
|
Pereira Filho A, Teixeira Filho J, Giongo V, Simões W, Lal R (2016). Nutrients dynamics in soil solution at the outset of no-till implementation with the use of plant cocktails in Brazilian semi-arid. African Journal of Agricultural Research 11:234-246. |
|
|
Pittelkow CM, Linquist B, Lundy ME, Liang X, van Groenigen KJ, Lee J, Gestel N, Six J, Venterea RT, Kessel C (2015). When does no-till yield more? A global meta-analysis. Field Crops Research 183:156-168. |
|
|
Potthoff M, Loftfield N (1998). How to quantify contamination of organic litter bag material with soil? Pedobiologia (Jena) 42:147-153. |
|
|
Roberts DP, Mattoo AK (2018). Sustainable Agriculture—Enhancing Environmental Benefits, Food Nutritional Quality and Building Crop Resilience to Abiotic and Biotic Stresses. Agriculture 8:8. |
|
|
Rockström J, Williams J, Daily G, Noble A, Matthews N, Gordon L, Wetterstrand H, DeClerck F, Shah M, Steduto P, de Fraiture C, Hatibu N, Unver O, Bird J, Sibanda L, Smith J (2017). Sustainable intensification of agriculture for human prosperity and global sustainability. Ambio 46:4-17. |
|
|
Séna Koglo Y, Abdulkadiri A, Dodzi Bissadu K, Kokouvi Akamah A (2017). Mitigating droughts effects on tropical agriculture systems: The role of improved soil management practices in regulating soil moisture, temperature and carbon losses. Journal of Soil Science and Environmental Management 8:104-112. |
|
|
Shanks R, Olson JS (1961). First year breakdown of leaf litter in Southern Appalachia Forest. Science 80(134):194-195. |
|
|
Taiz L, Zeiger E (2013) Fisiologia vegetal, 5th edn. Artmed, Porto Alegre, RS, Brazil. |
|
|
Talgre L, Roostalu H, Mäeorg E, Lauringson E (2017). Nitrogen and carbon release during decomposition of roots and shoots of leguminous green manure crops. Agronomy Research 15:594-601. |
|
|
Tarui A, Matsumura A, Asakura S, Yamawaki K (2013). Evaluation of Mixed Cropping of Oat and Hairy Vetch as Green Manure for Succeeding Corn Production. Plant Production Science 1008:1-10. |
|
|
Torres JLR, Pereira MG (2014). Produção e decomposição de resíduos culturais antecedendo milho e soja num latossolo no cerrado mineiro. Comunicata Scientiae 5:419-426. |
|
|
Van Soest PJ, Wine RH (1968). Determination of lignin and cellulose in acid detergent fiber with permanganate. Journal of the Association of Official Analytical Chemists 51:780-785. |
|
|
Wider RK, Lang GE (1982). A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology 63:1636-1642. |
|
|
Wutke EB, Calegari A, Wildner LP (2014). Espécies de adubos verdes e plantas de cobertura e recomendações para seu uso. In: Lima Filho OF, Ambrosano EJ, Rossi F, Carlos JAD (eds) Adubação verde e plantas de cobertura no Brasil: Fundamentos e práticas, 2nd edn. Embrapa Informação Tecnológica, Brasilia, DF. Brasil pp. 59-168. |
|
|
Xavier FA, Oliveira JI, Silva MR (2017). Decomposition and Nutrient Release Dynamics of Shoot Phytomass of Cover Crops in the Recôncavo Baiano. Revista Brasileira de Ciência do Solo. |
|
|
Xia M, Talhelm AF, Pregitzer KS (2017). Chronic nitrogen deposition influences the chemical dynamics of leaf litter and fine roots during decomposition. Soil Biology and Biochemistry 112:24-34. |
|
|
Yang SY, Huang TK, Kuo HF, Chiou TJ (2017). Role of vacuoles in phosphorus storage and remobilization. Journal of Experimental Botany 68:3045-3055. |
|
|
Zhang H, Zhang Y, Yan C, Liu E, Chen B (2016). Soil nitrogen and its fractions between long-term conventional and no-tillage systems with straw retention in dryland farming in northern China. Geoderma 269:138-144. |
|
|
Zhen L, Sheng W, Wang C, Zhang L (2017). Multifunctional Agriculture and the Relationship Between Different Functions. In: Zhang L, Schwärzel K (eds) Multifunctional Land-Use Systems for Managing the Nexus of Environmental Resources. Springer International Publishing, Cham pp. 53-67. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


