ABSTRACT
The increase in water scarcity, now made the researchers to look for various ways to decrease water use in rice production and increase the water use efficiency. One of the approaches that lead to a considerable amount of savings in water use by rice is aerobic cultivation, which minimizes the methane emission also. In this light of view, an investigation was carried out to study the morpho-physiological traits and yield potential of six rice genotypes viz., PMK 3, ASD 16, MDU 3, MDU 5, CO 47 and RM 96019 under aerobic condition. The parameters viz., plant height, total drymatter production, leaf area index, R:S ratio were found to be higher in the rice cultivar PMK 3. The physiological and biochemical traits viz., proline content, total phenolics, catalase and superoxide dismutase enzymes activities were found to be significantly higher in PMK 3 rice cultivar. Yield and yield components such as panicle length, number of productive tillers, number of spikelets per panicle, fertility co-efficient, panicle harvest index, grain yield and harvest index were recorded. The genotype PMK 3 recorded higher yield (4298 kg/ha) followed by ASD 16 (3068 kg/ha) and CO 47 recorded lower yield (1407 kg/ha) under aerobic condition.
Key words: Aerobic rice, morphology, physiological traits and yield.
Rice occupies the enviable prime place among the food crops cultivated around the world and it is the most important food crop in Asia. Water is a looming crisis due to competition among agricultural, industrial, environmental and domestic users. By 2025 AD, 30% of the human population would be threatened by water scarcity because of the fact that, 70% of water withdrawals is used in irrigated agriculture worldwide (IRRI, 2001). In Asia, more than 50% of irrigation water is used for irrigating rice. Rice is a heavy consumer of water requiring some 4000 to 5000 L to produce 1 kg of rice (Shen et al., 2001). A growing scarcity of fresh water will pose problems for rice production in future years. No wonder as the water shortage crisis has already reached parts of China as well as central and west parts of India, Pakistan and some parts of Bangladesh. Hence, shifting gradually from traditional rice production system to growing rice aerobically, especially in water scarce irrigated lowlands, can mitigate occurrence of water related problems. China has pioneered in the development of aerobic rice and so the IRRI is studying Chinese methods. The rice is called aerobic rice where the plants are grown under non-puddled and non-flooded soil condition. Growing rice aerobically saves water by: (i) eliminating continuous seepage and percolation (ii) reducing evaporation and (iii) eliminating wetland preparation (Castaneda et al., 2002). Aerobic rice cultivation is one of the ways to minimize the methane emission wherein rice is grown under non-flooded condition and it also saves 55 to 60% of water. Nevertheless, varieties adopted for aerobic condition are being tested in several countries particularly India, but are yet to be widely adopted. Hence, plant physiologists and breeders have to address the challenges in breeding varieties with better physiological adaptations for higher yields under aerobic conditions and also developing new ideotype concept for rice under aerobic environment.
The field experiment was carried out at Tamil Nadu Agricultural University, Coimbatore (11°N, 77°E, elevation of 426.72 m). The soil of the experiment site is clay loam. The available soil nitrogen, phosphorus and potassium were 247.5, 15.0 and 527.0 kg ha-1 respectively (Soil pH 7.3; EC 0.5 dSm-1). Six rice varieties viz., PMK 3, ASD 16, MDU 3, MDU 5, CO 47 and RM 96019 were directly sown at a spacing of 20 x 10 cm (Appendix 1).
The experiment was laid out in randomized block design with six treatments (genotypes) and four replications. Irrigation was given once in five days based on IW/CPE ratio (= 0.8) and hence the aerobic condition was maintained. Fertilizers at 120:38:38 N P K kg / ha was applied along with irrigation. The full dose of phosphorus applied as basal and N, K applied at three splits as topdressing. Proper weed management and plant protection measures were carried out at the appropriate time as per the recommendation. Plant samples were drawn at maximum tillering (50 to 55 DAS), panicle initiation (60 to 65 DAS), flowering (80 to 85 DAS) and maturity (110 to 115 DAS) stages for recording various morphological, physiological, biochemical and yield characters.
The height of the plant was measured from the base of the shoot to the longest leaf at different stages and mean worked out for the four plants in each replication and expressed in cm. For estimating total dry matter production (TDMP), the entire plant was pulled out with the root system intact, at different stages. The shoot and root portions were weighed separately after drying the plants at 80°C for 24 h and the TDMP was expressed in g plant-1. The leaf area index (LAI) was calculated by employing the formula of Williams (1946). Using shoot and root dry weight the root- shoot ratio (R:S ratio) was arrived. Stomatal conductance and transpiration rate were measured between 10.30 am to 11.30 am using a Steady State Porometer (PP systems, EGM 4 - PMR 5, version 5.02) at different phenological stages. The measurements were recorded on the topmost fully expanded leaf from five plants. The average values were computed and expressed as mmol m-2 s-1 for both the parameters. Proline content of the leaves was estimated by the method described by Bates et al. (1973). Phenol content of leaf extract was determined by using the method of Malik and Singh (1980) and expressed in mg g-1 fresh weight. Catalase activity was determined by titrimetric method using KMnO4 (Gopalachari, 1963) and expressed as μg H2O2 g-1 min–1. Super oxide dismutase activity was determined using Nitro Blue Tetrazolium (NBT) by the method of Beau Champ and Fridovich (1971) and expressed as enzyme units mg-1 of protein. The yield components viz., number of productive tillers, panicle length, fertility co-efficient, panicle harvest index (ratio of grain weight to panicle weight) and 1000 grain weight were determined. The data collected was subjected to statistical analysis in randomized block design (Gomez and Gomez, 1984).
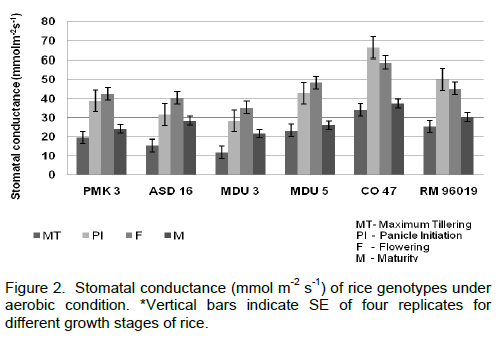
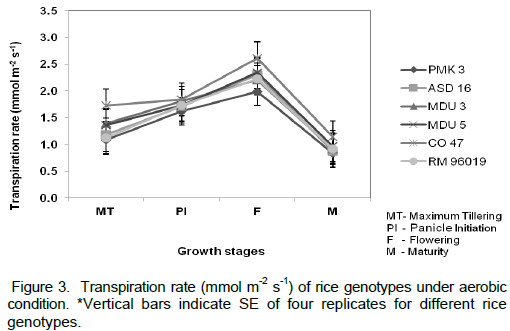
The morphological characters viz., plant height, TDMP, Root: Shoot ratio and LAI were measured. Irrespective of the genotypes, the plant height increased significantly from maximum tillering to maturity stage of the crop (Figure 1). Among the genotypes, PMK 3 showed higher plant height under aerobic condition. It has grown up to 84.15 cm followed by RM 96019 (74.55 cm) at maturity stage. The shortest plant height was recorded in MDU 5 (43.99 cm). TDMP showed significant variation in all the six rice genotypes (Table 1). The variety PMK 3 exhibited 22.75 g and CO 47 recorded the lowest value of 14.04 g at maturity stage. Highest LAI was observed at flowering stage for all the genotypes (Table 1). Among the genotypes, PMK 3 recorded higher LAI (4.92) and CO 47 (3.08) exhibited the least value. The maximum R:S ratio was noticed in PMK 3 (1.62) followed by ASD 16 (1.57) and CO 47 recorded lower value (1.25) when compared to other genotypes (Table 1). Data on stomatal conductance revealed that the cultivar CO 47 recorded higher value in all the stages and lower stomatal conductance was observed in MDU 3 under aerobic condition (Figure 2). Generally an increasing trend in transpiration rate was observed upto flowering for all the rice genotypes (Figure 3). PMK 3 recorded less transpiration rate (1.89 mmol m-2 s-1) followed by MDU 3 (2.20 mmol m-2 s-1) and RM 96019 (2.22 mmol m-2 s-1) at flowering stage and there after declining trend was observed at maturity stage in all genotypes. CO 47 showed higher transpiration rate in all the stages.


The data on proline content at various stages are furnished in Figure 4. Among the genotypes, PMK 3 recorded higher proline content of 987.50, 1223.75, 1768.75 and 843.25 µg g-1 at maximum tillering, panicle initiation, flowering and maturity stages respectively. The genotype CO 47 recorded the least value from tillering to maturity with a value of 652.50 to 982.40 µg g-1. At maturity stage, all the genotypes showed on par with each other except CO 47 for proline content. The data showed increasing trend in total phenolics from tillering to flowering stage after that significant reduction was observed. Accumulation of total phenolics was observed in all the genotypes under aerobic condition and the highest accumulation being recorded in PMK 3 and the lowest in CO 47 (Figure 5). In general, all the six rice genotypes recorded maximum value at flowering stage for total phenolics. The enzyme catalase activity exhibited increasing trend from tillering to flowering and afterwards declining trend was noticed (Figure 6). Among all the genotypes, PMK 3 expressed higher enzyme activity followed by ASD 16 and MDU 5. The genotypes MDU 5 and RM 96019 are on par with each other.



All the genotypes recorded higher superoxide dismutase enzyme activity during flowering stage and the least activity was observed in CO 47. This enzyme activity showed similar response like catalase activity. It exhibited increasing trend from tillering to flowering then gradual decline was noticed at maturity stage. PMK 3 recorded higher enzyme activity compared to other genotypes at all the stages of observation followed by ASD 16 and MDU 3 (Figure 7).

In related to yield and yield components (Table 2), the genotype MDU 3 recorded more numbers of productive tillers (516.24) followed by ASD 16 (504.36) and PMK 3 (468.25). The genotype CO 47 recorded the least number of productive tillers among all the genotypes. The rice genotypes PMK 3, RM 96019, ASD 16, MDU 5, MDU 3 and CO 47 recorded the panicle length as 22.83, 21.35, 19.38, 18.80, 18.78 and 17.51 cm respectively. Among them, PMK 3 recorded maximum panicle length. The data on panicle harvest index exhibited significant difference among all the genotypes. Between the genotypes, PMK 3 registered higher value of 79.65% followed by ASD 16 (77.82%), RM 96019 (76.84%) and MDU 3 (76.28%). The genotype CO 47 recorded lower value of 63.13%. PMK 3 recorded more number of spikelets per plant (116.12) followed by ASD 16 (111.26) and MDU 3 (106.02). The genotype CO 47 recorded the least value (98.96) among all the genotypes for number of spikelets per plant. The genotype PMK 3 recorded the spikelet fertility co-efficient of 85.31% followed by RM 96019 (84.79%) (Table 2). The genotype PMK 3 recorded the maximum 1000 grain weight of 21.12 g followed by ASD 16 (20.06g), RM 96019 (19.54g), MDU 3 (19.46 g), MDU 5 (19.42 g) and CO 47 (19.18 g).
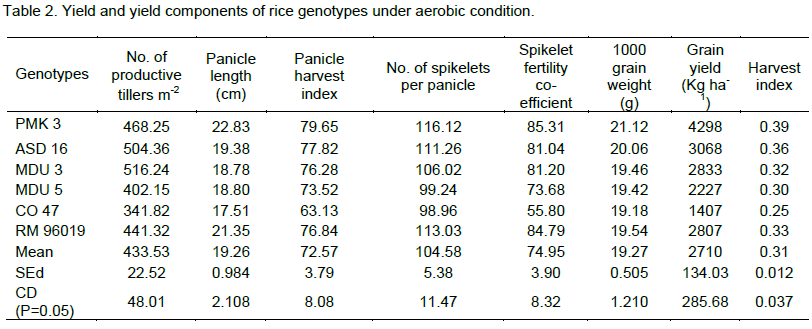
Among the six rice genotypes, PMK 3 recorded the highest yield of 4298 kg ha-1 followed by ASD 16 (3068 kg ha-1). The other genotypes, MDU 3, MDU 5, RM 96019 and CO 47 registered 2833, 2807, 2227 and 1407 kg ha-1 respectively for grain yield. The data on harvest index showed significant differences among the rice genotypes. PMK 3 recorded higher harvest index of 0.39 followed by ASD 16 (0.36) and RM 96019 (0.33). The genotype CO 47 recorded the least value of 0.25 due to lower grain yield.
Irrespective of the genotypes, the plant height increased significantly from maximum tillering to maturity stage of the crop (Figure 1). Among the six rice genotypes, PMK 3 showed higher plant height under aerobic condition when compared with other genotypes. The present findings were in accordance with the findings of Russo (2000) and Gowri et al. (2005). Insalud et al. (2004) reported that the genotype Kae Noi had increased root length and decreased shoot length in well drained soil. Tomar and Prasad (1996) reported that the plant height in rice was associated with thick and deep root system. The total dry matter production showed significant variation in all the rice genotypes (Table 1). The variety PMK 3 accumulated more dry matter at maturity stage followed by ASD 16 and the genotype CO 47 recorded the lowest value. Lilley and Fukai (1994) reported that water stress reduced the biomass production in rice varieties and the degree of reduction depended upon the severity of moisture stress. The maintenance of higher biomass during stress may be viewed as one of the criteria to decide drought tolerance. Drymatter yield of rice genotypes were reduced by 11 to 37% and 30 to 65% under mild and severe moisture stress, respectively (Yang et al., 1995). Sritharan et al. (2004) also reported similar results in rice under stress condition. Gowri (2005) observed that under aerobic condition drymatter production was reduced drastically compared to flooded condition. Nieuwenhuis et al. (2002) reported that continuous aerated condition produced 32% lower biomass than continuously flooded treatment. It may be because of low moisture stress induced impaired tillering (Yoshida, 1981) or due to accelerated leaf senescence and hence decreased photosynthetic area under moisture stress (Simane et al., 1993) leading to lower drymatter production.
Leaf area index (LAI) particularly at reproductive stage has a specific role in deciding drymatter production and grain yield of rice (Yoshida, 1981). In the present investigation, PMK 3 recorded higher LAI (4.92) but CO 47 (3.08) exhibited the least value (Table 1). Gowri (2005) also reported the reduction in LAI under aerobic condition in ADT 43 compared to PMK 3. The reduction in LAI might be due to rapid decline in leaf elongation as reported by Lilley and Fukai (1994). It was observed that, the root to shoot ratio (R:S ratio) of rice plant increases with drought (Yoshida, 1981). Root characteristics in rice are genetically controlled but they are also strongly affected by soil conditions and crop management practices (Sharma et al., 1987 and Thangaraj and Sivasubramanian., 1990). The R:S ratio was high in drought resistant upland rice cultivars than susceptible cultivars (Lu and Hua, 1994). In the present study, PMK 3 registered higher R:S ratio (1.62) than other genotypes (Table 1). Similar findings were also recorded by Gowri (2005) on PMK 3 rice variety under aerobic condition.
Stomata are minute pores through which gases diffuse out and into leaves. In the present study, MDU 3 recorded lower stomatal conductance under aerobic condition compared to other genotypes (Figure 2). Higher diffusive resistance in response to water stress is a characteristic behaviour of upland varieties (O’ Toole and Cruz, 1980) because of their higher stomatal sensitivity (Dingkuhn et al., 1989) and it is one of the drought avoidance mechanisms in rice. However, the higher stomatal conductance was recorded in CO 47 as this variety might have suffered severely from water deficit due to aerobic condition. Increased diffusive resistance was found to be associated with accumulation of phenols suggesting that phenolics might lead to stomatal closure. This is corroborated by the reports of Manthe et al. (1992). The cultivar PMK 3 showed lower transpiration rate than other genotypes (Figure 3). Thus, this genotype might have reduced the water loss. Similar observations were also reported by Dingkuhn et al. (1989) and Chauhan et al. (1996).
Proline accumulation under water stress may occur in different ways viz., stimulation of synthesis from glutamic acid perhaps due to loss of feedback inhibition of the synthesis of the intermediate 4’ – pyroline - 5 - carboxylate; inhibition of proline oxidation; impaired incorporation of free proline into protein and reduced export of proline via phloem. Reasonable hypothesis based on the possible role of proline accumulation under water stress condition has been advanced as a nitrogen store, energy store, NH3 – detoxification product and cytosolic osmoregulation. Results of the present study indicated that the cultivar PMK 3 recorded higher proline accumulation than the other genotypes due to aerobic condition (Figure 4). Similar finding was reported by Gowri (2005) stated that PMK 3 accumulated higher amount of proline than ADT 43 under aerobic condition. Sritharan and Mallika (2006) also reported that higher proline content under moisture stress condition in the tested rice genotypes.
There are diverse groups of phenolic compounds found in all plant parts. Phenols are known to regulate a range of growth and development processes in plants. Accumulation of total phenolics was observed in all the genotypes under aerobic condition and the highest accumulation was recorded in PMK 3 and the lowest in CO 47 (Figure 5). Similar results were reported by Ashraf et al. (1994) under water stress in wheat. Manthe et al. (1992) opined that phenolic compounds at higher concentration reduced the transpiration rate by closing the stomata. This observation could be confirmed in the present study as the tolerant cultivar PMK 3 recorded more phenolics content and lower transpiration rate. Characterization of the accumulated phenolic compounds might provide clear information regarding the role played by phenolics in unsaturated soil moisture condition.
Decreasing water potential and stomatal closure resulted in the accumulation of activated oxygen species (Sgherri et al., 1993). Increased photorespiration under water stress also elevate glycolate oxidase activity resulting in H2O2 production (Mittler and Zilinskas, 1992). Allen (1995) reported that generation of activated oxygen species under water stress might be responsible for most of the damage caused to cellular components. Shigenaga et al. (1994) further reported that superoxide, peroxide and hydroxyl radicals were highly reactive and were linked to degenerative diseases and aging. The present study revealed that the antioxidant enzymes viz., catalase and superoxide dismutase activity were over expressed under aerobic condition regardless of genotypes (Figures 6 and 7). Further, the activity of these enzymes in tolerant genotype PMK 3 was much higher than in susceptible cultivar CO 47. This result was in conformation with the findings of Zelitch (1990) who reported that drought induced damage was negated with increasing activities of SOD and catalase. The lesser activity of these enzymes in the susceptible cultivar CO 47 indicated that these enzymes degraded at higher rate under aerobic condition, while their synthesis might have been inhibited. Similar observations were also reported by Zhang and Kirkam (1994) and Ranjita et al. (1994) in wheat species and Srivalli et al. (2003) in upland rice.
Yield and yield attributing parameters are the ultimate manifestation of a plant’s ability to survive, grow and produce yield under drought regardless of the tolerance mechanisms involved. Aerobic condition significantly reduced grain yield in CO 47, mainly through reduction in filled grain percentage, spikelets number per panicle and number of productive tillers per hill, but the cultivar PMK 3 recorded higher yield components and grain yield (Table 2). Fukai et al. (1991) reported close relationship of grain yield under water stress with grain number and grain weight was relatively stable across environments. Further grain number was determined by spikelet number and filled grain percentage.
The reduced spikelet number per panicle may probably be due to reduced LAI. Increased diffusive resistance might have reduced photosynthesis leading to reduced assimilate supply as suggested by Cruz and O’Toole (1984) Boonjung (1993).
The present study revealed that the cultivar CO 47 showed reduced panicle harvest index (PHI), poor fertility co-efficient and harvest index. These observations were supported by earlier reports of Boonjung (1993) and Lilley and Fukai (1994). Limitations of assimilate supply particularly during reproductive stage affected spikelet number per panicle and filled grain percentage (Cruz and O’Toole, 1984). Reduction in leaf water potential at anthesis caused poor panicle exertion (Ekanayake et al., 1989) and this might be the reason for less number of grains due to pollination abnormalities. This is in conformity with the findings of Gowri (2005) in rice under aerobic condition.
The variety PMK 3 has showed higher root:shoot ratio, dry matter production as well as antioxidant enzymes activities which leads to more yield under aerobic condition. These findings are much important for identification of traits for the wider adaptation of aerobic rice under water limited conditions. Hence, shifting gradually from traditional rice production system to growing rice aerobically, especially in water scarce irrigated lowlands, can mitigate occurrence of water related problems and there by lesser green house gas emission.
The authors have not declared any conflict of interest.
REFERENCES
Allen RD (1995). Dissection of oxidative stress tolerance using transgenic plants. Plant Physiol. 107:1049-1054.
CrossRef
|
|
|
|
Ashraf MY, Azmi AR, Khan AH, Ala SA (1994). Effect of water stress on total phenols peroxidase activity and chlorophyll content in wheat (Triticum aestivum L.). Acta Physiol. Plant. 16(3):185-191. |
|
|
Bates LS, Waldren RP, Teak TD (1973). Rapid determination of free proline for water stress studies. Plant Soil. 39:205-207.
CrossRef |
|
|
Beau-Champ C, Fridovich I (1971). Superoxide dismutase: Improved assays and assay applicable to acrylamide gels. Anal. Biochem. 44:276-287.
CrossRef |
|
|
|
Boonjung H (1993). Modelling growth and yield of upland rice under water limiting conditions. Ph.D. Thesis, University of Queensland. |
|
|
|
Castaneda AR, Bouman BAM,Peng S Visperas RM (2002). The potential of aerobic rice to reduce water use in water- scarce irrigated low lands in the tropics. In: Water wise rice production.(Ed.). Bouman, B.A.M., H. Hengsdijk, B.Hardy, P.S. Bindraban, T.P. Tuong and J.K.Ladha. IRRI., Losbanos, Philippines. |
|
|
|
Chauhan JS, Moya TB, Singh RK Singh CV (1996). Growth and development under different soil moisture regimes in upland rice (Oryza sativa L.). Indian J. Plant. Physiol. 1:270-272. |
|
|
Cruz RT, O'Toole JC (1984). Dryland rice response to an irrigation gradient at flowering stage. Agron. J. 76:178-182.
CrossRef |
|
|
Dingkuhn M, Cruz RT, O'Toole JC, Dorffling K (1989). Net photosynthesis, water use efficiency, leaf water potential and leaf rolling as affected by water deficit in tropical upland rice. Aust. J. Agric. Res. 40:1183-1192.
CrossRef |
|
|
|
Ekanayake IJ, Datta SK, Steponkus PL (1989). Spikelet sterility and flowering response of rice to water stress at anthesis. Ann. Bot. 63:257-264. |
|
|
Fukai S, Li L, Vizmonte PT, Fischer KS (1991). Control of grain yield by sink capacity and assimilate supply in various rice cultivars. Exp. Agric. 27:127-135.
CrossRef |
|
|
|
Gomez KA, Gomez AA (1984). Statistical procedures for Agricultural Research. An IRRI book, Wiley Inter Science publication, John Wile and Sons, New York, USA. P. 680. |
|
|
|
Gopalachari NC (1963). Changes in the activities of certain oxidizing enzymes during germination and seedling development of Phaseolus mungo and Sorghum vulgare. Indian J. Exp. Biol. 1:98-100. |
|
|
|
Gowri S (2005). Physiological studies on aerobic rice (Oryza sativa L.). M.Sc., thesis submitted to Tamil Nadu Agricultural University, Coimbatore, India. |
|
|
|
Gowri S, Vijayalakshmi C, Sritharan N (2005). Impact of aerobic condition on growth parameters and yield in rice. J. Agric. Res. Manage. 4:117-118. |
|
|
|
Insalud N, Bell RW, Rerkasem B (2004). Response of rice cultivars to aerobic condition. In: Fourth International Crop Congress, Sep 26-Oct 1, Brishane, Australia. |
|
|
|
IRRI (2001). Enhancing water productivity in rice-based production systems In: Enhancing productivity and sustainability of favorable environments project summary reports. pp .25-27. International Rice Research Institute, Manila, Philippines. |
|
|
Lilley JM, Fukai S (1994). Effect of timing and severity of water deficit on four diverse rice cultivars. III. Phenological development, Crop growth and grain yield. Field Crops Res. 37:225-234.
CrossRef |
|
|
|
Lu F, Hua HJ (1994). A study on the main drought resistant index of land rice. Acta. Agric. Boreali Sinica. 9:7-12. |
|
|
|
Malik CP, Singh MB (1980). In: Plant enzymology and Histoenzymology. Ludhiana, Kalyani Publishers. |
|
|
Manthe B, Schulz M, Schnable H (1992). Effect of salicylic acid on growth and stomatal movements of Vicia faba: evidence for salicylic acid metabolization. J. Chem. Ecol. 18:1525-1539.
CrossRef |
|
|
Mittler R, Zilinskas BA (1992). Regulation of pea cytosolic ascorbate peroxidase and other antioxidant enzymes during the progression of drought stress and following recovery from drought. Plant J. 5:397-405.
CrossRef |
|
|
|
Nieuwenhuis J, Bouman BAM, Castaneda A (2002). Crop -water responses of aerobically grown rice: preliminary results of pot culture experiments. In: BAM Hengdijik, H Hardy, PS Bindrabab, STP Toung, JK Ladha (eds.), Water- Wise Rice Production. IRRI. Pp. 177-185 |
|
|
|
Ranjita B, Rana D, Patel BB, Monoranjan A (1994). Alterations in the activities of active oxygen scavenging enzymes of wheat leaves subjected to water stress. Plant Cell Physiol. 35:489-495. |
|
|
|
Russo S (2000). Preliminary studies on rice varieties adaptability to aerobic irrigation. Cashiers Options Méditerranéennes. 15:35-39. |
|
|
Sgherri CL, Pinzino MC, Navari-Izzo F (1993). Chemical, changes and O2 production in thylakoid membranes under water stress. Physiol. Plant. 87:211-216.
CrossRef |
|
|
Sharma PK, De Datta, SK, Redulla, CA (1987). Root growth and yield response of rainfed lowland rice to planting method. Exp. Agric. 23:305-313.
CrossRef |
|
|
Shen L, Courtis B, McNally KL, Robin S, Li Z (2001). Evaluation on near isogenic lines of rice introgressed with QTL for root depth through marker aided selection. Theor. Appl. Genet. 103:75-83.
CrossRef |
|
|
Simane B, Pea Cock JM, Struik PC (1993). Differences in developmental plasticity and growth rate among drought resistant and susceptible cultivars of durum wheat (Triticum turgidum var. durum). Plant Soil. 157: 155-166.
CrossRef |
|
|
|
Sritharan N, Annie Sheeba J, Mallika Vanangamudi (2004). Mitigating drought in rice by foliar spray of chemicals and plant growth regulators. J. Agric. Res. Manage. 3(2):45-47. |
|
|
|
Sritharan N, Mallika Vanangamudi (2006). Screening of rice genotypes for salt tolerance. Plant Archives. 6(2): 815-818. |
|
|
Srivalli B, Sharma G, Chopra RK (2003). Antioxidative defense system in an upland rice cultivar subjected to increasing intensity of water stress followed by recovery. Physiol. Plant. 119:503-512.
CrossRef |
|
|
|
Thangaraj M, Sivasubramanian V (1990). Physiological response of rice to water stress at reproductive phase. Oryza, 27:493-495. |
|
|
|
Tomar JB Prasad SC (1996). Relationship between inheritance and linkage for drought tolerance in upland rice (Oryza sativa L.) varieties. Indian J. Agric. Sci. 66:459-465. |
|
|
|
Williams RE (1946). The physiology of plant growth with special reference to NAR. Ann. Bot. 10:21-27. |
|
|
|
Yang JC, Zhu QS, Wang ZQ (1995). The effects of soil moisture on the yield and physiological characteristics of rice. Acta Agron. Sinica. 21:110-114. |
|
|
|
Yoshida S (1981). Fundamentals of rice crop science. International Rice Research Institute, Los Banos, Philippines. |
|
|
Zelitch I (1990). Further studies on O2 resistant photosynthetic and photorespiration in a tobacco mutant with enhanced catalase activity. Plant Physiol. 92:352-357.
CrossRef |
|
|
|
Zhang J, Kirkam MB (1994). Drought stress induced changes in activities of SOD, catalase and peroxidase or proline oxidase in wheat species. Plant Cell Physiol. 35:785-91. |
|
|
|
Zhang KF, Qing M, Huan HK (1997). Effect of water deficit on physiological activities of paddy rice and upland rice seedlings. J. Shandong Agric. Univ. 28: 53-55. |



