Full Length Research Paper

ABSTRACT
At the application time of sugarcane ripeners, most times, morning glory plants, that have escaped, survived or emerged, are entwined on sugarcane stalks and these plants are at this time in different reproductive stages (and it becomes more difficult to control) causing a mechanical and crop yield damage. The main aim of this research was to evaluate sugarcane crop ripeners effect on the seeds germination, seedling development and the abscission of the reproductive structures of morning glory. The treatments consist of five ripeners: ethyl-trinexapac (300 ga.i.ha-1), glyphosate (216 ga.i.ha-1), fluazifop-p-butyl (75 ga.i.ha-1), sulfometuron-methyl (15 ga.i.ha-1), ethephon (480 ga.i.ha-1) and one control plot (without ripener application). They were applied during the reproductive stages of morning glory. The experimental design used was completely randomized design with four replications and six treatments, to a total of 24 plots. After ripeners application, abscised reproductive structures were counted to determine the percentage of abscission. The seeds of each phenological stage were collected when it reached the point of physiological maturity and it was set to germinate in Petri dishes to evaluate the germination rate, germination mean time, germination velocity, initial growth and dry mass weight. Trinexapac-ethyl, glyphosate and fluazifop-P-butyl application when plants are with open flowers can reduce seed viability. Glyphosate showed the smallest length value and it can affect seedling growth. Glyphosate and ethephon caused the highest percentages of abscission of morning glory reproductive structures and they can contribute to reduce the seeds inflow to the soils.
Key words: Abscission, Ipomoea hederifolia, plant growth regulator, reproductive structure
INTRODUCTION
Sugarcane became one of the main crops of Brazilian economy (Mapa, 2016). In the last decades the system passed through major changes due the prohibition of burning the sugarcane areas before harvesting in São Paulo state, mainly because of environmental and social impacts derived from the crop (Monqueiro et al., 2011). With the adoption of this practice, harvesting after burning is being substituted with raw sugarcane harvesting, following progressive legal restrictions. As a consequence, the raw sugarcane system provided important modifications for weeds (Kuva et al., 2011). The remaining straw above the surface that originates from the harvesting process changes the light, temperature and humidity conditions, and these factors favor the development of weeds with voluble stems in those areas, such as the morning glory (Azania et al., 2011).
The morning glory develops at the same time with the biggest sugarcane growth phase, and adult specimens become intertwined in stalks and leaves, hindering the crop development, the agricultural practices and harvesting operations (Azania et al., 2002). Silva et al. (2009) verified that the presence of Ipomoea hederifolia during the entire crop cycle of RB855536 sugarcane reduces 46% of its productivity. The same authors also wrote that during harvesting, its fruits and seeds may be found attached to the mother-plant, thus allowing the dissemination of these seeds by the harvester machines to medium and long distances.
After the biggest sugarcane growth phase, the stabilization of its growth and the maturation starts. During this process, the morning glory plants show various flowering fluxes that, usually, coincide with the sugarcane ripener application. Most of these chemicals are used to provide a consistent ripening effect on sugarcane, to suppress flowering (which can reduce sugar content) and to enhance sucrose concentration (Dalley and Richard Jr., 2010). Among traditional products used as sugarcane ripeners that belong to growth inhibitors group or herbicide compounds group are glyphosate, sulfometuron-methyl, ethephon, trinexapac-ethyl, and fluazifop-P-butyl (Leite et al., 2008).
This way, it is expected that ripeners applied on these plants may reduce the fruit production and, consequently, seeds. This abortion induction and or seed reduction may happen depending on the reproductive stage of the plant. New alternatives which egress blockage of viable weed seeds on the ground, eliminating the production and/or viability of these seeds still on the mother-plant as well as hinder the development of seedlings represent promising implements for weed management.
Given all that has been explained above, the aim of this work is to evaluate the usage of sugarcane ripeners on the seed germination, seedling development andreproductive structures abscission of I. hederifolia.
MATERIALS AND METHODS
The experiment was developed at Universidade Estadual Paulista “Júlio de Mesquita Filho”, Jaboticabal Campus, São Paulo State, located at 21°14’05” S, 48°17’09” W coordinate and 615 m altitude. I. hederifolia seeds were seeded on raised beds of 50 cm width and 50 cm length.
As soon as the species had reached the four stages (or phenophases) needed for the experiments, five ripeners were applied: ethyl-trinexapac (300 ga.i.ha-1), glyphosate (216 ga.i.ha-1), fluazifop-p-butyl (75 ga.i.ha-1), sulfometuron-methyl (15 ga.i.ha-1) and ethephon (480 ga.i.ha-1) on the I. hederifolia plants, and a control plot was left without application.
On I. hederifolia, those stages were defined following Bleiholder et al. (1991), through the BBCH-Code scale, which enables one to identify weeds development stages (Table 1). Each stage was marked with different colored ribbon strip, to identify which flowering of fructification stage that branch was on the application day. The experimental design used was completely randomized design with four replications and six treatments, to a total of 24 plots. For each plot, 30 reproductive structures per stage were marked, to a total of 124 reproductive structures per plot.

Ripener application was performed with a CO2 pressurized backpack sprayer equipped with a XR 110.02 nozzle. The equipment was regulated at 2.2 bar pressure applying 200 L.ha-1 of herbicide mix.
After the ripener application, daily observations were carried out on the number of marked structures on which abscission was carried out in each phenophase during the period between ripener application and seed harvesting, to determine the percentage of structures that suffered abscission. After that, the fruits of each treatment, when they have reached maturation point, were harvested for their seeds (right before the dehiscence).
For the next part of the experiment, the seeds were tested through tetrazolium chloride test for each batch (Brasil, 2009). Seeds were then put to germinate on Petri dishes with filter paper as medium, embedded with 2.5 times the weight of paper on water and arranged on BOD (Biological Oxygen Demand) germination chambers at 27°C temperature. The experiment was conducted on completely randomized design, with four replications and 25 seeds per Petri dish. Daily observations were carried for 15 days after the first seed germinated (radical > 2 mm), counting the number of germinated seeds.
Thereafter, the seeds from each stage were evaluated by the germination test, through analyzing the germination rate, germination velocity index (Maguire, 1962), mean germination time (Labouriau, 1983), length and dry matter of seedlings aerial parts and roots.
The results were analyzed on completely random design on a 6x4 split-plot design (treatments x stage), being the whole-plot A and split-plot B. Results were submitted to analysis of variance and means compared through the Tukey test at 5% probability.
RESULTS AND DISCUSSION
Abscission of reproductive structures
Analyzing the treatments among each stage, all the applied products provoked increase on abscission percentage when compared to the control plot (Table 2).
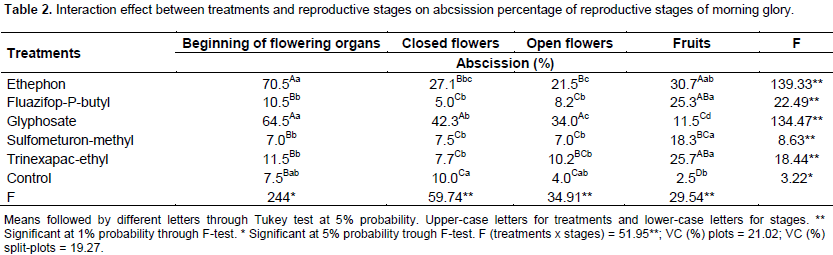
The results obtained with glyphosate show a more harmful effect on the beginning of flowering structures, still closed flowers and open flowers, at 57, 32 and 30%, respectively, compared to the control plot. Viator et al (2003) showed increased number of cotton bolls abscised per plant with the increasing of glyphosate rates.
When the fruits were formed, the abscission due to glyphosate application is 9% higher. In this stage, the abscission was very low (35% lower than previous stages).
Ethephon application caused an increase of 63, 17, 28 and 28% on beginning of flowering structures, closed flowers, open flowers and fruits stages, when compared to the control plot. On the remaining treatments (trinexapac-ethyl, Fluazifop-P-butyl and sulfometuron-methyl), the results differ only on the fruit stages, on a 20% higher mean value. Hole and Hardwick (1978), Iglesias et al. (2006), Ninot and Romero (2012) reported that ethephon may promote the abscission of different plant organs such as leaves, flowers, and fruits.
Among the stages, it is possible to see the application on the beginning of flowering organs, closed and open flowers promoted the highest abscission rates when glyphosate and ethephon were applied. The application on the fruits, all treatments provided increase on the abscission rates.
The results found for glyphosate and its effects on the abscission rates in the reproductive stages may be explained by the raise of ethylene and the consequential cellulase activity raise on the abscission zones, diminishing the resistance to abscission. That occurs because amino acid and ureid biosynthesis branches into tryptophan, phenylanine and tyrosine synthesis, which is inhibited by glyphosate, or in glycine, serine, cysteine and methionine. When the first branch is inhibited, the second one is more induced. It is possible to find the presence of methionine in it, which is the precursor of ethylene synthesis (Yamada and Castro, 2007).
The abscission of flowering structures; abscission after ethephon application may occur due to auxin transport inhibition caused by ethylene liberation that happens when the product contact the plant tissue. Auxin is produced in the leaves and an auxin gradient from the leaf blade to the stem keeps the petiole abscission zone insensitive to ethylene. The ethylene diminishes the auxin activity for the reduction of its synthesis and transport as well as raise of its destruction (Taiz and Zeiger, 2013). The physiological effect of IAA, in this process, is to reduce the sensibility of the abscission zone to ethylene. On low IAA concentrations, the “active” ethylene, after the application, result in abscission (Bangerth, 2000).
Germination percentage
At the beginning of flowering stages, it can be noted that with or without application, there are no significant differences, however, the germination being lower than in the later stages (Table 3).

Analyzing the stage closed flowers; the ripeners have results similar to the control. On open flowers, the treatments as a whole, caused reduction related to the control plot, of 26%. At fruit stage, the ripeners do not differ among themselves or the control plot.
Germination on the beginning of the flowering structures may have occurred due the dormancy mechanism characteristic of some weed species, e.g. primary dormancy, which, following Viana et al. (2008), is intrinsic of weed seeds, developed while still attached to the mother-plant, remaining after the dispersion. Likewise, dormancy is a perpetuation and defense mechanism, producing seed lots with variation on the germination times (Marcos Filho, 2005).
It is notable that in the fourth stage the mean germination percentage was higher than in the initial stages. Knowing that after the fecundation, seeds grow fast in size and reaches a maximum growth in a short period (Carvalho and Nakagawa, 2012), the usage of cell division inhibitors, which molecules are ready for translocation to the meristem tissues (Shaner et al., 1984) and highly movable to the meristematic areas with high growth rates (Rodrigues, 1995) explain, the efficiency of ripeners on the initial stages of reproductive development.
Velocity index
For the beginning of flowering structures, sulfometuron-methyl and ethephon are the ones that presented the highest values among all treatments, taking a longer time for germination processes (Table 4).

On the closed flowers stage, the ripeners showed results similar to those obtained from the control plot, while the open flowers stage showed mean values higher than the control, as it happened to the germination percentage. The effect of treatments on the fruits stage shows no significant difference.
The germination velocity index was similar to the pattern attained in the germination percentage for every treatment, except for ethephon on the closed flowers stage, being the lowest values for velocity for the also lowest values of germination percentage. On the open flowers stage, the values obtained from ripener application treatments also don’t go over the control plot’s. The vigor level of the seed may affect the plant development and proportionate lower competition capacity of these for light, water and nutrients with neighboring plants. According to Oliveira et al. (2009) the germination velocity test is based on the principle of seeds that have lesser velocity are less vigorous.
Mean germination time
The effect of treatments on the mean germination time shows no significant difference, except in the beginning of the flowering structures stage there is difference for glyphosate, Fluazifop-P-butyl, sulfometuron-methyl e ethephon, which exhibit the lowest time to germination (Table 5). The comparison among the stages regarding the treatments show the beginning of flowering structures stages has a slower germination than the other treatments.

Aerial parts and root length
The interaction results between stages and treatments upon the aerial parts length are on Table 6. Glyphosate treatment showed the lowest length when applied on closed and open flowers, with a 24% variation, when it was compared to control.
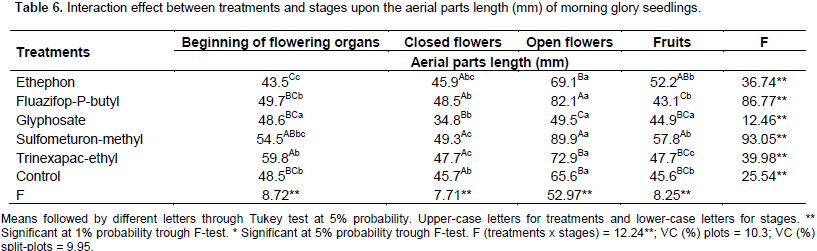
It is valid to point that for trinexapac-ethyl, fluazifop-P-butyl and sulfumeturon-methyl, these treatments added to the increase to in length, in some stages. Trinexapac-ethyl increases 9.0% when applied in the beginning of flowering structures while fluazifop-P-butyl and sulfumeturon-methyl increased 25 and 37% in the open flowers. These treatments did not showed significant differences on the others stage compared to the control plot. For ethephon treatment, in all stages, the results were similar to the control plot results.
Table 7 revealed the interaction results between treatment and stages for root length. Glyphosate shows lower results for every stage applied than any other. When set against the control plot, the reductions are of 68, 10, 80 and 87%, for each of the treatments, respectively.
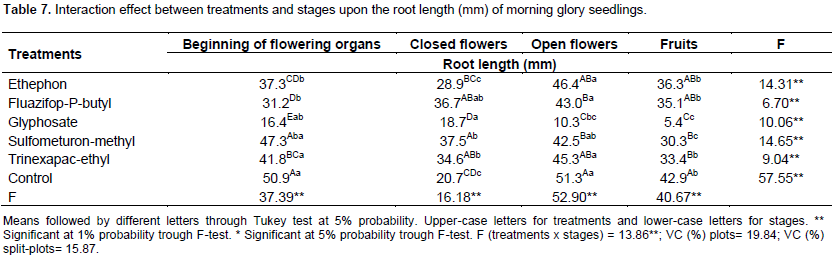
Trinexapac-ethyl treatment, when compared to the control plot, shows reduction of 18% (beginning of flowering structures) and 22% (fruit). However, when in the closed flowers, the root length was 67% higher.
As for the effect of fluazifop-p-buthyl on the length, it is noted that, when applied in the beginning on flowering structures and open flowers, there were 39 and 16% lower than the control plot. Yet, when applied on the closed flowers, the length was 77% higher. The results from sulfometuron-methyl and ethephon were similar to the control.
During the root length evaluation (third and fourth stages), the seedlings from glyphosate application did not show primary root development and didn’t emit secondary roots either. Pline et al. (2001) reported that 30% of absorbed glyphosate may be retained in the reproductive tissues. Viator et al. (2003) reported that squares and bolls of resistant cotton retained 0.2 to 3.7% of the applied glyphosate. Funguetto et al. (2004) presented results alike, studying the glyphosate effects on non GMO soy plants, which had reduction in the root structures due the inhibition of primary roots and the secondary roots emission, being the hypocotyls proportionally bigger than the primary root, characterizing an anomaly.
The limitation on the root length may be explained by the diminishing of IAA production during the seedling development. As the translocation of glyphosate through the plant occurs through symplastic way, the application of agrochemicals on source organs (mature leaves) enable the translocation of said product to sink organs (growing organs) on the rest of the plant, simultaneously to the photoassimilates (Peterson et al., 1978; Caseley and Coupland, 1985; Franz et al., 1997), and so, glyphosate would be translocated to the forming seeds in the mother-plant, and by accumulating on the seeds, may cause the inhibition of IAA hormone (indoleacetic acid auxin) during the plant development. The IAA, hormone produced on the tip regions of a plant (roots and stem), is a substance related to the growth regulation, derived from triptofan through many phases, and, also, independent of triptofan, as does its precursor the indol-3-glycerol phosphate, which depends on chorismate for its formation. Glyphosate inhibits the synthesis of chorismate as well as triptofan synthesis (Yamada and Castro, 2007).
Aerial parts and roots dry matter content
On Table 8 are the interaction results between treatments and stages upon aerial parts dry matter. For all treatments, in fruits stage, the results did not differ from control plot. When flowers were open, the dry matter content for all treatments, except for ethephon, decreased around 25% (glyphosate) and 50% (fluazifop-P-butyl, sulfometuron-methyl and trinexapac-ethyl).
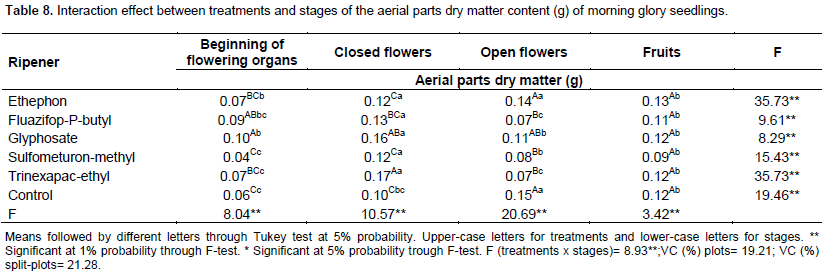
In the beginning of the flower structures and in the closed flowers ethephon and sulfometuron-methyl treatments showed the same results to control plot. As for glyphosate and fluazifop-P-butyl, it increases the mass when applied to the beginning of the flower structures (about 58%) and in closed flowers, where both accumulated 60% (glyphosate) and 30% (fluazifop-P-butyl) more than the control.
Trinexapac-ethyl, in the beginning of the flower structures, did not differ from control plot, while in the closed flowers, it accumulated 70% more dry matter content than control plot.
Table 9 shows the interaction results between treatments and stages of roots dry matter. For the fruit stage, as it happened in the aerial parts dry matter content, no results were different. In the beginning of the flowering stages, trinexapac-ethyl, fluazifop-P-butyl, sulfumeturon-methyl and etephon have more dry matter than the control. Fluazifop-P-butyl is the treatment that most increased this parameter.
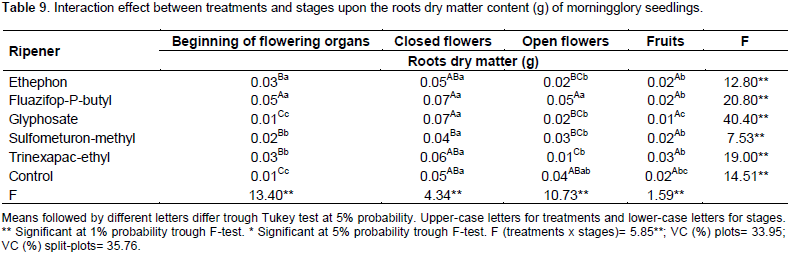
The application of these ripeners on the closed and open flowers have similar results to the control plot, except to trinexapac-ethyl treatment, on the open flowers application, that provided the least dry matter accumulation between the treatments, 75% lower than control. The glyphosate results were similar to the control in all stages.
The difference between the results of dry matter may be related to the product accumulation in the seeds when it receives the photoassimilates upon the seed formation stage, and, though, the malformation of these during the development of the mother-plant, causing negative effects on the seed quality. It is also important to highlight that the application of the products was before the fruits reached the physiological maturity; therefore, the photoassimilate transport was not finished (Carvalho and Nakagawa, 2012).
The data found for the ripeners’ applications on the reproductive stage indicate that those ripeners may affect the initial development of the seedlings which originated from the mother-plant, and, hence, provide lesser competition characteristics for light, water and nutrients with the desirable crop. Besides, in having reduced fruit and seed production, as well as increasing the abscission of reproductive structures diminishes the weeds seed bank resupplying.
CONCLUSION
Glyphosate (216 g a.e ha-1) and ethephon (480 g a.i ha-1) may reduce the egress of I. hederifolia to the ground. Trinexapac-ethyl (300 g a.i.ha-1), glyphosate (216 g a.e ha-1) and fluazifop-P-butyl (75 g a.i.ha-1) applied when flowers are open, reduced the seeds viability.
The application of glyphosate (216 g a.e ha-1) during the reproductive stages of Ipomoea hederifolia may affect the seedlings development.
CONFLICT OF INTERESTS
The author(s) have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors are grateful to CAPES for financing this Project and to CNPq for the concession of research funding for Alves.
REFERENCES
|
Azania CAM, Hirata ACS, Azania AAPM (2011). Technical Bulletin: Biology and chemical management of morninglory in sugarcane. IAC (Technical Bulletin IAC 209). |
|
|
Azania AAPM, Azania CAM, Gravena R, Pavani MCMD, Pitelli RA (2002). Sugar cane (Saccharum spp.) straw interference in emergence of weed species of the convolvulaceae family. Planta Daninha 20(2):207-212. |
|
|
Bangerth F (2000). Abscission and thinning of young fruit and their regulation by plant hormones and bioregulators. Plant Growth Regulation 31(1):43-59. |
|
|
Bleiholder H, Kirfel H, Langeluddeke P, Stauss R (1991). A uniform code for the phenological stages of crops and weeds. Pesquisa Agropec. Bras. 26(9):1423-1429. |
|
|
Brasil Ministério da Agricultura e Reforma Agrária (2009). Regras para análise de sementes. Brasília, SNAD/DNDV/CLAV P 365. |
|
|
Carvalho NMDE, Nakagawa J (2012). Sementes, ciência, tecnologia e produção. 5. Ed. Jaboticabal; FUNEP. 590 p. |
|
|
Caseley JC, Coupland D (1985). Environmental and plant factors affecting glyphosate uptake movement and acidity. In: Grossbard, E, Atkinson DA (Ed.). The herbicide glyphosate. London: Butterworths, pp. 92-123. |
|
|
Dalley CD, Richard Jr EP (2010). Herbicides as ripeners for sugarcane. Weed science 58(3):329-333. |
|
|
Franz JE, Mao MK, Sikorski JA (1997). Glyphosate: A unique global herbicide. American Chemical Society. Washington, DC: American Chemical Society pp. 521-605. |
|
|
Funguetto CI, Tillmann MAA, Villela FA, Dode LBD (2004). Bio assays for detection of soybean seeds tolerant to glyphosate herbicide. Revista brasileira de sementes 26(1):130-138. |
|
|
Pline WA, Price AJ, Wilcut JW, Edmisten KL, Wells R (2001). Absorption and translocation of glyphosate in glyphosate-resistant cotton as influenced by application method and growth stage. Weed Sci. 49:460-467. |
|
|
Hole CC, Hardwick RC (1978). Chemical aids to drying seeds of beans (Phaseolus vulgaris) before harvest. Ann Appl. Biol. 88(3):421-427. |
|
|
Iglesias DJ, Tadeo FR, Primo-Millo E Talon M (2006). Carbohydrate and ethylene levels regulate citrus fruitlet drop through the abscission zone A during early development. Trees: Structure and Function 20(3):348-355. |
|
|
Kuva MA, Pitelli RA, Alves PLCA, Salgado TP, Pavani MCMD (2008). Weed seedbank and its correlation with the established flora in no-burn sugarcane area. Planta Daninha 26(4):735-744. |
|
|
Labouriau LG (1983). A germinação das sementes. Organização dos Estados Americanos. Programa Regional de Desenvolvimento Científico e Tecnológico. Série de Biologia. Monografia 24:174. |
|
|
Leite GPH, Crusciol CAC, Lima GPPL, Silva MDEA (2008). Growth regulators and technological quality of sugarcane in the middle of the cropping season. Ciência e Agrotecnologia 32(6):1843-1850. |
|
|
Marcos Filho J (2005). Fisiologia de sementes de plantas cultivadas. Piracicaba: FEALQ. 495 p. |
|
|
Maguire JD (1962). Speed of germination - aid in selection and evaluation for seedling emergence and vigor. Crop Sci. 2:176-177. |
|
|
MAPA - Ministério da Agricultura, Pecuária e Abastecimento (2016). Citros. Available in: <View>. |
|
|
Monqueiro PA, Silva PV, Hirata ACS, Martins FRA (2011). Weed Infestation Maps under Different Sugarcane Harvest Systems. Planta Daninha 29(1):107-119. |
|
|
Ninot A, Romero A, Tous J, Batlle I (2012). Effect of loosening agent sprays on the efficiency of the mechanical harvesting of 'Arbequina' Olives. Hortscience 47(10):1419-1423. |
|
|
Oliveira ACS, Martins GN, Silva RF, Vieira HD (2009). Testes de vigor em sementes baseados no desempenho de plântulas. Inter Science Place 2. |
|
|
Peterson CADE, Wildt PPO, Edgington CV (1978). A rationale for the ambimobile translocation of the nematicide oxamyl in plants. Pest. Biochem.Physiol. 8:1-9. |
|
|
Rodrigues J D (1995). Fisiologia da cana-de-açúcar. Botucatu: Instituto de Biociência 101 p. |
|
|
Shaner DL, Anderson PC, Stidham MA (1984). Imidazolinones: Potent inhibitors of acetohydroxyacid synthase. Plant Physiol. 76(2):545-546. |
|
|
Silva IAB, Kuva MA, Alves PLCA, Salgado TP (2009). Interference of a Weed Community with Predominance of Ipomoea hederifolia on Sugar Cane Ratton. Planta Daninha 27(2):265-272. |
|
|
Taiz L, Zeiger E (2010). Plant physiology. 5. ed. Sinauer Associates. Sunderland, MA 782 p. |
|
|
Viana RS, Silva PH, Mutton MA, Mutton MJR, Guimarães ER, Bento M (2008). Effects on the application of chemical ripeners on the culture of sugarcane (Saccharum spp.), SP81-3250 variety. Acta Sci. Agron. 30(1):65-71. |
|
|
Viator RP, Senseman SA, Cothren JT (2003). Boll abscission responses of glyphosate-resistant cotton (Gossypium hirsutum) to glyphosate. Weed Technol. 17:571-575. |
|
|
Yamada T, Castro PRC (2007). Efeitos do glifosato nas plantas: implicações fisiológicas e agronômicas. International Plant Nutrition Institute. Piracicaba: Informações Agronômicas 24 p. (Technical Bulletin 119). Available in: |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


