ABSTRACT
A parasite study was conducted from August 2004 to July 2005 to highlight the distribution of gill Monogenean parasites from Oreochromis niloticus. A total of 370 fish collected from Ayamé man-made Lake, Côte d’Ivoire were examined for Monogenean species using routine parasitological techniques. The study revealed the presence of six species of Monogeneans (Cichlidogyrus thurstonae, Cichlidogyrus halli, Cichlidogyrus tilapiae, Cichlidogyrus rognoni, Cichlidogyrus cirratus and Scutogyrus longicornis). These species showed an aggregate pattern within the host population. Four species of parasites showed positive and significant correlation with parasitic abundance and the relative condition factor. The prevalence and intensity of the infection of these parasites were highest in the long rainy season and lowest in the dry season. The large sized fishes were more subjected to parasitic infection than smaller ones. In contrast, no sex effect was observed on the infestation of O. niloticus by the Monogeneans. No preference for the right or left side of the fish host was observed and the parasite species mostly colonized the middle arches (II and III). This could lead to more informed aquacultural practises, and maximization of the production of O. niloticus.
Key words: Disease, season, sex, infestation, gill.
Monogenean trematodes belong to one of the most species-rich classes of fish parasites. They are commonly found on fish gills and skin. Nevertheless, some Monogenean species invade the rectal cavity, ureter, body cavity, nostrils, intestine, stomach and even the vascular system (Rohde et al., 1992; Pariselle and Euzet, 1998; Whittington et al., 2000). Their life cycle involves only one host and they mostly spread by way of egg releasing and free-swimming infective larvae (Öztürk and Özer, 2014). In the natural environment, Monogeneans are often in equilibrium with their hosts, and can cause serious morbidity and mortality during fish breeding (Buchmann and Lindenstrom, 2002; Kayis et al., 2009; Strona et al., 2010) resulting in significant economical losses (Ghittino et al., 2003; Johnson et al., 2004; Boungou et al., 2008; Velloso and Joaber, 2010). The Nile tilapia, Oreochromis niloticus, indigenous to the Nile basin, plays an important role in aquaculture globally(Akoll et al., 2011) could face such problem. This fish has a great economic interest in Côte d’Ivoire (Gole-Bi et al., 2005). Indeed, the total annual yield of inland fisheries estimated at 18 000 tons are essentially dominated by Cichlidae, notably O. niloticus which represents between 50 to 70% of the total catches (Gole-Bi et al., 2005).
The Food and Agricultural Organization of the United Nations (FAO, 2009) reported that, to satisfy an increasing demand in freshwater fish, extensive research must include studies of their parasites for optimal production levels. The knowledge of fish parasites is of particular interest in relation not only to fish health but also to understanding ecological problems (Sures, 2001; Dudgeon et al., 2006). Baseline data, collected from infections within natural water systems, serve as reference point for management strategies.
In tropical Africa, Monogeneans parasites are mostly known in their taxonomical aspect (Pariselle and Euzet, 2009) and specificity and biological cycle (Aloo, 2002; Tombi and Bilong Bilong, 2004). Studies concerning eco-biological aspects of these parasites are scarce (Nwani et al., 2008; Keremah and Inko-Tariah, 2013, Tombi et al., 2014). To the best of our knowledge, there has only been two ichthyoparasitological studies (Blahoua et al., 2009, 2015) on infection dynamics of gill Monogenean parasites of Ivoirian fish.
Here we present some eco-biological aspects of Monogeneans’ gill parasites of Oreochromis niloticus in Lake Ayamé I, Côte d’Ivoire.
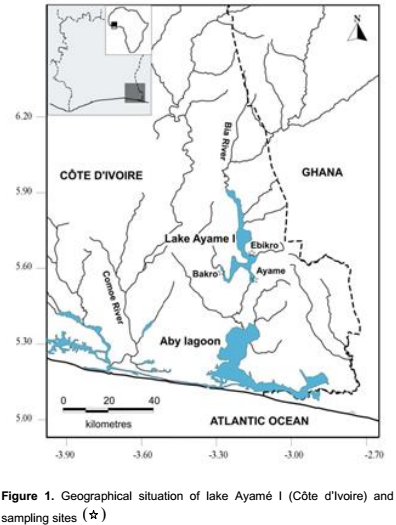
Study area
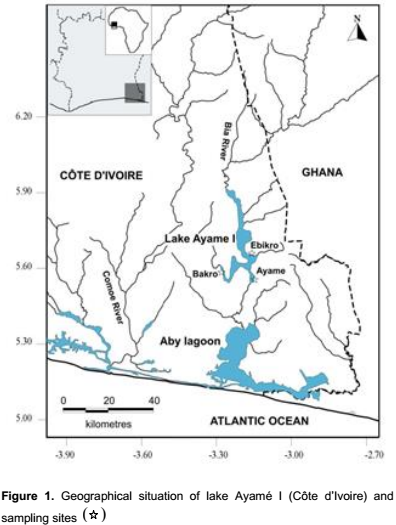
Lake Ayamé I, the oldest hydroelectric dam of Côte d’Ivoire being commissioned for hydroelectric generation in 1959 (Reizer, 1967) has an average surface of 90 km2 and its maximum depth is 20 m (Gourene et al., 1999) (Figure 1). The reservoir is situated at 3°-3°5’ W, 5°30’-6°N in the Southeast of the country. The climate of lake area in the Bia watershed is an equatorial transition zone with two rainy seasons separated by a short dry period in August and September and a more pronounced dry season from December to March. The lake is characterized by two high water level periods, April to June and September to November (with maximum discharge of about 500 m3 s-1). Conversely, low water level periodsare December to March and July to August (with a minimal discharge of about 0.13 m3 s-1). Mean annual water surface temperature in the reservoir is 28°C (Ouattara et al., 2007). Water surface temperature varied from 25.9 to 26.2°C during the small rainy season and from 29.8 to 32.7°C in the long rainy season (Blahoua et al., 2009). The average dissolved oxygen and water transparency ranged between 2.8 to 15.4 mgl-1 and 550 to 1371 mm respectively (Nobah et al., 2008).
Fish sampling and parasitological analyses
From August 2004 to July 2005, a total of 370 O. niloticus individuals were collected from the described three main landing sites every month, during 3 to 4 days per site. Fishing was done with multi-meshed nylon gillnets (8 to 90 mm mesh sizes). Each gill net measures 30 m long by 1.5 m deep. Nets were set overnight (17 to 07h) and during the following day (07 to 12 h).
Once out of the water, the fish were immediately identified following Teugels and Thys van den Audernaerde (2003) keys. The weight of the fish were taken using digital weighing balance and recorded. The standard length (SL) of each specimen was measured to the nearest 1 mm on a measuring board. The effect of the parasites on the health status of the fish host was investigated from Fulton’s condition factor (K-factor) with the following Formula: Kc= W x 105/L3, where W is the weight (grams) and L the standard length of fish (millimeters) (Klemm et al., 1992). The fish sample were divided into four different length classes of 50 mm amplitude, which are: Class I (50 ≤ SL < 100), class II (100 ≤ SL < 150), class III (150 ≤ SL < 200) and class IV (200 ≤ SL ≤ 250). Fishes were killed by a single cut through the spinal cord and dissected to determine their sex. Subsequently, gill arches on each side were removed fresh from bucco-pharyngeal cavity by dorsal and ventral sections and then stored in ice (0°C).
At the laboratory, each gill arch was placed in a separate Petri dish containing water, and examined for Monogenean parasites under a binocular microscope (Olympus SZ 60). Gill arches from each side of the fish were numbered I-IV from the anterior portion of the gill arch below the operculum to the posterior. Individual worms were collected and mounted on a slide in a drop of ammonium picrate-glycerine mixture (Malmberg, 1957). The identification of the parasite species observed were done with a microscope magnification of 400 and 1000X, on the basis of available taxonomic characters as described by Pariselle and Euzet (2003, 2009).
The total number of parasites counted from the entire gill section was expressed in terms of prevalence, abundance and mean intensity following the formula proposed by Bush et al. (1997). Aggregation was calculated using dispersion index (S2/x) to determine distribution pattern of parasites (Poulin, 1993; Shaw and
Dobson, 1995).
Statistical analysis
The Sperman’s correlation coefficient “rs” was used to investigate correlations of abundances of parasites with the relative condition factor of the hosts (Zar, 1996). The Chi-square (χ2) test was used to compare two or more proportions (prevalence) of samples. Analysis of variance and Student U test were used to determine the existence of any meaningful difference in mean intensity of the parasites species. Statistical significance level was evaluated at of 5% using STATISTICA 6.0.
Monogenean parasite species composition
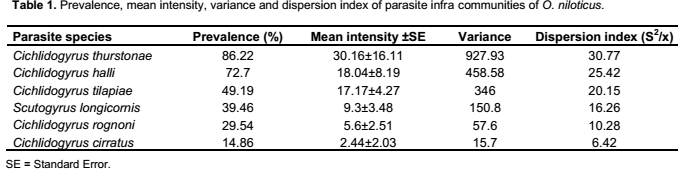
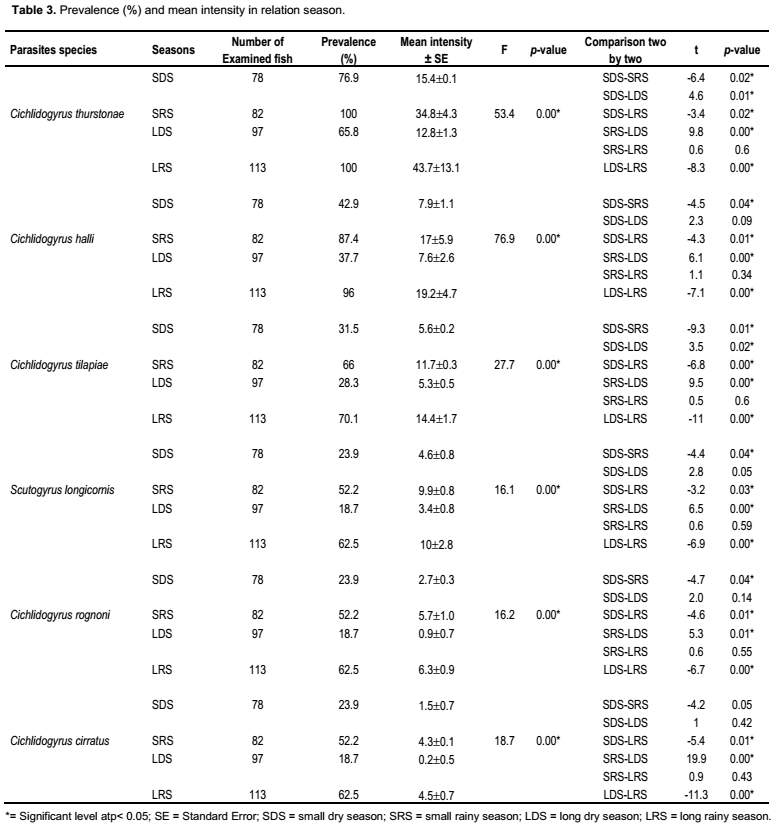
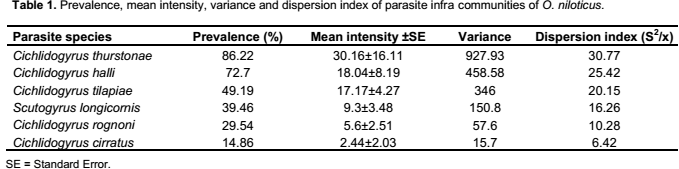
Six species belonging to two different genera: Cichlidogyrus (C. thurstonae Ergens, 1981; C. halli (Price and Kirk, 1967); C. tilapiae Paperna, 1960; C. rognoni Paperna, 1964; C. cirratus Paperna, 1964) and Scutogyrus (S. longicornis (Paperna and Thurston, 1969)), were collected on the gills of Oreochromis. niloticus. Prevalence, mean intensity and the aggregation index are reported (Table 1).These data revealed that all the Monogenean parasites adopted an aggregation distribution (variance is greater than mean intensity).
Four of the six parasites species (C. thurstonae, C. halli, C. tilapiae and S. longicornis) studied have an abundance significantly and positively correlated with the condition factor of the fish (rs = 0.73, 0.69, 0.59 and 0.38, respectively, p˂ 0.05) (Table 2).
Temporal variation of the occurrence of the parasites
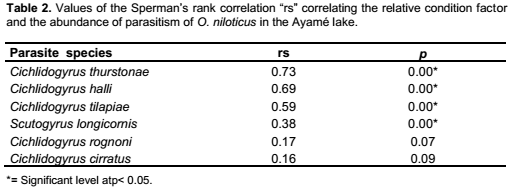
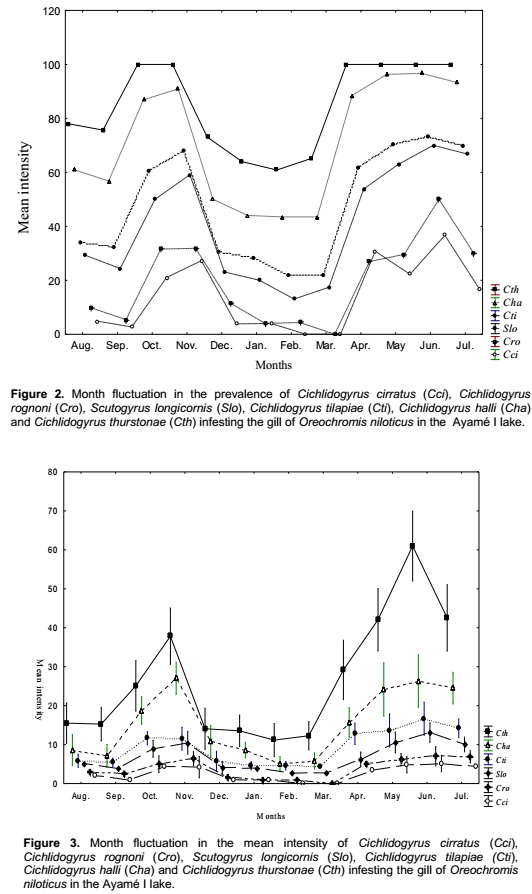
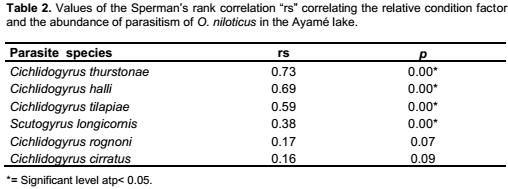
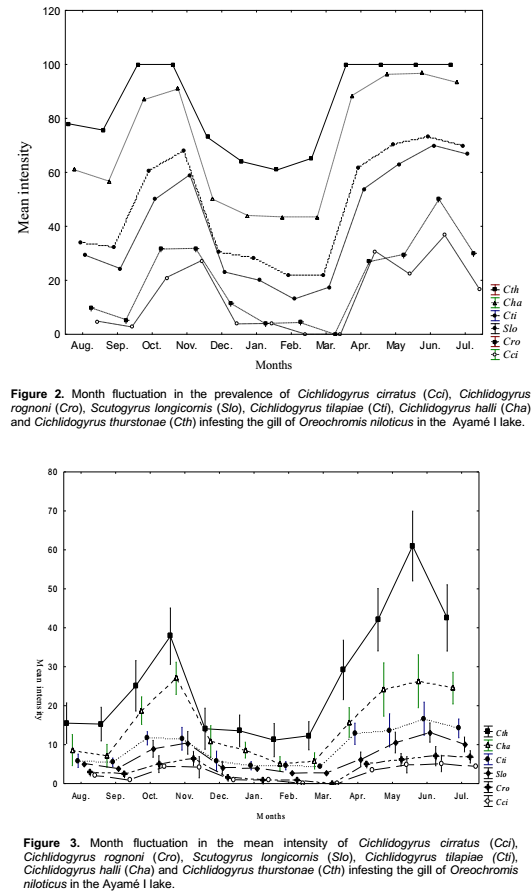
C. thurstonae occurred on the host population during all the study period. A bimodal curve showed its maximum prevalence (100%) in October and November 2004 (small rainy season) and from April to July 2005 (long rainy season). The lowest infestation rate (60.9%) was recorded in February 2005 (long dry season) (Figure 2). The prevalence variations during the four seasons were significant (χ2 = 70.3, df = 3, p < 0.05). Mean intensity values varied from 11.2 ± 1 in February 2005 (long dry season) with a peak of 61 ± 2.1 in June 2005 (long rainy season) (Figure 3). This variation curve was similar to the prevalence one. Significant difference was found in intensity of infection between seasons (Analysis of variance, F = 53.4). Parasitism changed from one season to another (Student, p < 0.05) (Table 3).
Scutogyrus longicornis, Cichlidogyrus tilapiae and C. halli were also present during the year of study. The prevalence variations of these parasites were similar to that of C. thurstonae but the values remained a little lower compared to the first one. For S. longicornis and C. tilapiae, the lowest prevalences (13 and 21.7%) were found in February 2005 (long dry season) and the highest (70 and 73.3%) in June (long rainy season), respectively. The number of fish infested by C. halli ranged from 43.5% in February and March 2005 (long dry season) to 96.7% in June (long rainy season) (Figure 2). The infestation showed a clear seasonal pattern (χ2 = 56.6 and 54.7 and 80.3, df = 3, respectively, p < 0.05). The mean intensity reached the highest values 10.5 ± 0.7, 11.9 ± 0.8 and 26.4 ± 1 in November 2004 (small rainy season) and 13.1 ± 0.5, 16.8 ± 1.1 and 27 ± 1.6 in June (long rainy season), respectively (Figure 3). The statistical analyses revealed that the infection of this host with these three parasites differed significantly between seasons (Analysis of variance, F = 16.1 and 27.7 and 76.9, respectively, df = 3, p< 0.05). There was also significantly difference from one season to another (Student test, p < 0.05) (Table 3).
For Cichlidogyrus cirratus and C. rognoni, the prevalence of infestation (varied from 3.8% in December (long dry season) to 36.7% in June (long rainy season) and from 4% in January (long dry season) to 50% in June (long rainy season), respectively. There was a lack of parasites in February and March 2005 (Figure 2). The Chi-square (χ2) applied to the temporal variations of the occurrence of these parasites showed that these values were statistically significant at 5% (χ2 = 38.1 and 41.7, df = 3, respectively). In both species, the highest mean intensities were recorded in November 2004 (small rainy season) and June 2005 (long rainy season). The values of mean intensity of C. cirratus were 4.4 ± 0.8 and 5.2 ± 0.5 then 6.4 ± 0.4 and 7.1 ± 0.6 for C. rognoni, respectively (Figure 3). Seasonal changes of mean intensity of infection followed the seasonal pattern of prevalence (Analysis of variance, F = 18.7 and 16.2, respectively, df = 3, p < 0.05). Intensity of infection changed from one season to another except from the small to the long dry season (p < 0.05) (Table 3).
Spatial distribution of Monogeneans on gills
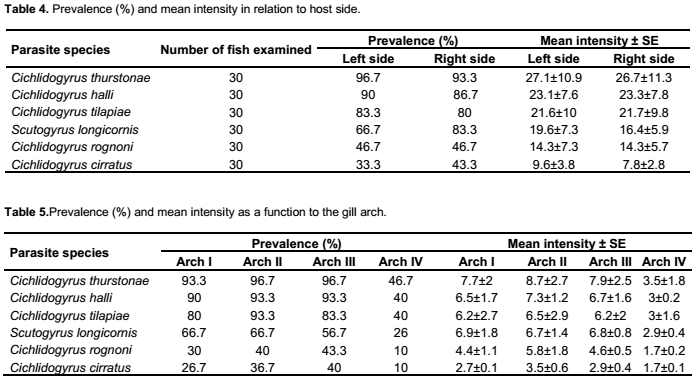
The prevalence and mean intensity values of Cichlidogyrus thurstonae were 96.7% and 27.1 ± 10.9 on the left side and 93.3% and 26.7 ± 11.3 on the right side of fish, respectively (Table 4). These values were not statistically significant at 5% (χ2 = 0.3, df = 1; analysis of variance, F = 0.06, p > 0.05). This species was more frequent and concentrated on arches II and III (χ2 = 38.9; df = 3, analysis of variance, F = 5.5, p < 0.05) (Table 5).
Scutogyrus longicornis, Cichlidogyrus tilapiae and C. halli were present in 66.7, 83.3 and 90% on the left side and 83.3, 80 and 86.7% on the right one, respectively. There was no significant difference between the Monogenean prevalence of left and right pairs of fish (χ2 = 2.2, 0.1 and 0.2 df = 1, p ˃ 0.05). Mean intensity values were 19.6 ± 7.3, 21.6 ± 10 and 23.1 ± 7.6 on the left set of gill arch of the fish while they were 16.4 ± 5.9, 21.7 ± 9.8 and 23.3 ± 7.8 on the right one, respectively (Table 4). In these three cases the differences were not statistically significant (Analysis of variance, F = 0.23, 0.03 and 0.02, df = 1, respectively, p ˃ 0.05). The data analysis showed that S. longicornis, C. tilapiae and C. halli were more frequently found on the median arches while the arch IV was the least infected (respectively χ2 = 13, 25.9 and 37.3, df = 3, p < 0.05). These species were more concentrated on the median arches (Analysis of variance, F = 7.1, 5.7 and 5.7 respectively, df = 3, p < 0.05) (Table 5).
The prevalence values of Cichlidogyrus cirratus and C. rognoni were 33.3 and 46.7% on the left side and 43.3 and 46.7% on the right one, respectively. These Monogeneans did not show preference for the left or right side of the gills (respectively (χ2 = 0.6 and 0.1, df = 1, p > 0.05). Intensity of infection was 9.6 ± 3.8 on the left side and 7.8 ± 2.8 on the right side for C. cirratus while was 14.3 ± 7.3 on the left and 14.3 ± 5.7 on the right side for C. rognoni (Table 4). There was no significant difference between the intensity of infection (Analysis of variance, F = 0.04 and 0.69, df = 1, respectively, p ˃ 0.05). In these two cases the significance difference was observed between the fourth and the rest of the gill arches (respectively, χ2 = 8 and 9.5, df = 3, analysis of variance F = 1.7 and 9.4, p < 0.05) (Table 5).
Relationship between body length and degree of infection
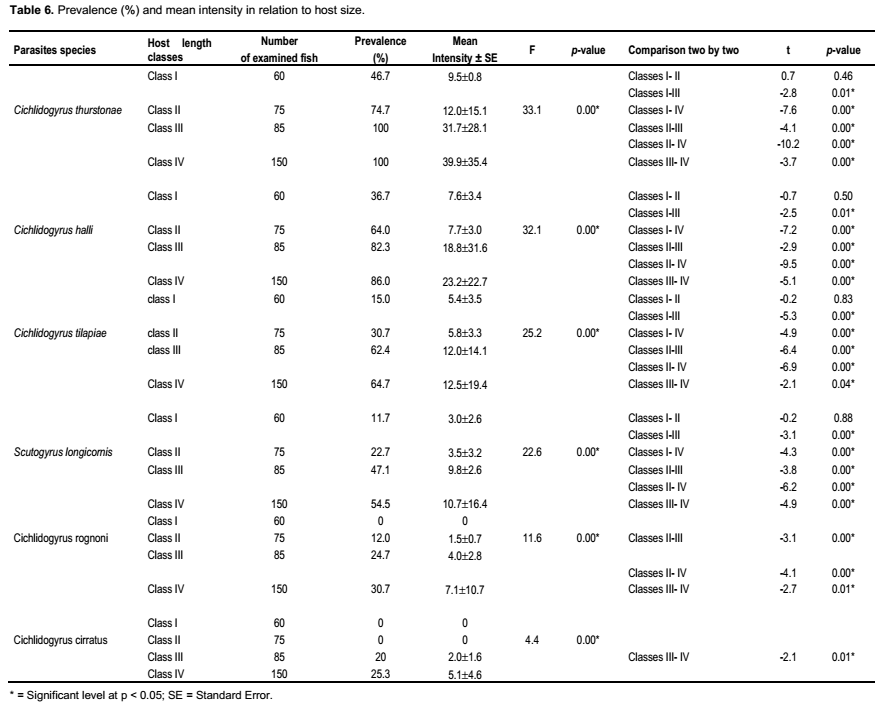
All the Monogenean species infested individuals of all any length class, except for C. cirratus and C. rognoni. For each parasite, the prevalence was lowest in the smallest length class (standard length ranges 50 to 100 mm) (Table 6).
The highest prevalence 100% of C. thurstonae was observed in the fish of the classes with standard length ranges greater than 150 mm. Statistical tests showed significant difference in prevalence according to the size classes (χ2 = 124.96, df = 3, p < 0.05). The values of mean intensity were 9.5 ± 0.8 in the first class and 12.0 ± 15.1, 31.7 ± 28.1 and 39.9 ± 35.4 in the other ones respectively (Analysis of variance, F = 33.1, dl = 3, p < 0.05). Globally, there was also a significant difference from one size class to another (Student test, p < 0.05) (Table 6).
The prevalence of Scutogyrus longicornis, Cichlidogyrus tilapiae and C. halli increased progressively with host size and reached the highest values 54.5, 64.7 and 86% in the largest individuals, respectively (χ2 = 43.7 and 58.6 and 59.5, df = 3 ; p < 0.05).
The values of mean intensity of S. longicornis were 3.0 ± 2.6, 3.5 ± 3.2 and 9.8 ± 2.6 in the first three classes respectively and 10.7 ± 16.4 in the latter class, while they were 5.4 ± 3.5 and 5.8 ± 3.3 in the z first two classes then 12.0 ± 14.1 and 12.5 ± 19.4 in the two latter classes for C. tilapiae. The lowest rates infestation (prevalence and mean intensity) of C. halli were observed in the smallest length class (36.7%, 7.6 ± 3.4) respectively. The highest values were observed in the largest individuals (standard length ranges 200 to 250 mm) (86%, 23.2 ± 22.7). Intensity of infection was varied significantly by host length class (Analysis of variance, F = 22.6 and 25.2 and 32.1, respectively, df = 3, p < 0.05). Student test revealed that there was significant difference from one size class to another (p < 0.05) (Table 6).
For C. cirratus and C. rognoni, the lowest prevalence 20 and 12% were recorded in the fish of the classes with standard length range 150 to 200 mm and 100 to 150 mm, respectively and the highest values 25.3 and 30.7% in the latter class. The Chi-square (χ2) indicated that the prevalence of these two species depends on the size of the host (χ2 = 38.3 and 79.6, respectively, df = 3; p < 0.05). The values of mean intensity varied from 2.0 ± 1.6 to 5.1 ± 4.6 for C. cirratus whereas varied from 1.5 ± 0.7 to 7.1 ± 10.7 for C. rognoni. The difference was statistically significant (Analysis of variance, F = 4.4 and 11.6, respectively, df = 3, Student, p < 0.05) (Table 6).
Relationship between host sex and degree of infection
The prevalence of C. thurstonae was 85% for male fish and 87.7% for female one. On female fish the mean intensity was slightly higher (31 ± 0.6) than on male fish (29.4 ± 0.3). Host sex no significantly affects the infection (χ2 = 0.5, df = 1; Analysis of variance, F = 3.7, p > 0.05).
The species S. longicornis, C. tilapiae and C. halli were present in 38.5, 52.8 and 69% of male host and 46.5, 50.6 and 77.1% of the female host, respectively. The intensities of infection were 7.8 ± 0.6, 11.2 ± 1.03 and 18.7 ± 0.01 in the male and 9.5 ± 0.3, 11.2 ± 1.5 and 17.4 ± 0.3 in the female. These values were not statistically significant at 5% (χ2 = 2.8, 0.1 and 3.01 respectively, df = 1; Analysis of variance, F = 0.12, 0.01 and 0.02, respectively, p > 0.05).
Prevalence and mean intensity of C. cirratus were 15% and 3.8 ± 0.9 for male fish and 14.7% and 4.4 ± 0.7 for female fish. The prevalence of C. rognoni was 20% in the male and 21.2% in the female. Intensity of infection was 5.7 ± 0.3 for male fish and 21.2% and 5.6 ± 0.7 for female fish. These parasites were also partitioned equally between males and females (χ2 = 0.08 and 0.01, respectively, df = 1, p > 0.05; Analysis of variance, F = 0.1 and 0.16, respectively, dl = 1, p > 0.05).
The present study recorded six species of Monogenea community on Oreochromis niloticus: Cichlidogyrus thurstonae, C. tilapiae, C. halli, C. rognoni, C. cirratus and Scutogyrus longicornis. This species richness is different of those reported by the previous studies. Boungou et al. (2008) found out five on the six Monogenean species in the same fish from the dam of Loumbila (Burkina Faso) whereas Tombi et al. (2014) reported four species in Melen station in Yaoundé (Cameroon). Usually, the number of Monogenean species per fish host varied (Pariselle, 1996) and this variability of parasite richness has been associated to various factors related to experimentation (Walter et al., 1995), the phylogeny of hosts and parasites (Bush et al., 1997; Sasal et al., 1997), host (Morand et al., 1999), ecology (Zharikova, 2000) and water quality (Galli et al., 2001; El-Seify et al., 2011). Our result suggests that the water of Lake Ayamé I may provide better eco-climatic conditions for the development of these parasites and can facilitate the contact between the infesting stages of these Monogenean and the host fish.
The exploitation of a host fish by several genera of Monogeneans has been already reported. Blahoua et al. (2009) mentioned the presence in Sarotherodon melanotheron, the genus Scutogyrus and several species of Cichlidogyrus. Ibrahim (2012) showed that eight Monogenean species colonized the gills of Tilapia zillii. Moreover, the colonization of hosts by several congeneric species was also reported by Bittencourt et al. (2014) and Tombi et al. (2014). This simultaneous infection could be explained by the fact that in natural environment, the parasitic densities are generally weak and therefore, the niches are always available on the gill biotope (Buchmann and Lindenstrom, 2002; Simkova et al., 2006).
Regarding parasite dispersion, in most cases, the parasites are almost universally aggregated between their hosts (Krasnov and Poulin, 2010).
Indeed, most of the hosts have few if any parasites, while a small number of hosts are infected with many parasites (Poulin, 1993).
This pattern is expected in most animals in nature, as observed within Monogeneans (C. thurstonae, C. tilapiae, C. halli, C. rognoni, C. cirratus and S. longicornis) parasitizing the gills of O. niloticus in the present study. This trend has been also evidenced for C. thurstonae, C. tilapiae, C. halli and S. longicornis Monogenean gill parasites of the same host by Tombi et al. (2014). According to Combes (1995), an aggregative distribution may indicate heterogeneity in the relationship between the host and the parasite populations. The probability for the parasite to meet its host and its chances of surviving in the latter, it may vary from one host to another. In addition, Kennedy (1977) stated that aggregative distribution increase the opportunities for parasites to meet a partner in order to reproduce.
In several studies, there has been a growing interest on the total number of parasites per host fish with the condition factor of the fish. Yamada et al. (2008) found significant and positive correlation of the condition factor of cichlids with the abundance of a species of Monogeneans. Lizama et al. (2007) found negative and significant correlation when relating the Piaractus mesopotamicus (Holmberg, 1887) condition factor of fish farming with the levels of parasitism of two species of Monogeneans. Tozato (2011) found no differences in condition factor of Corydoras aeneus (Gill, 1858) parasitized and not parasitized by Monogeneans, and concludes that these parasites did not affect the welfare of the host. The author also stated that the parasite community of fish studied has low pathogenicity to the host in the different environments of the basin, indicating the proper response of fish against parasitism. In this study, it was observed that the condition factor of O. niloticus had significant and positive correlation with the abundance of parasitic species C. thurstonae, C. tilapiae, C. halliand S. longicornis. Cone (1995) stated that larger fish and a better condition factor can withstand higher intensities of infection by Monogeneans parasites despite being pathogenic. This is possibly the reason for the significant and positive correlation between condition factor studied fish and abundance of parasitism by Monogeneans.
Concerning the temporal variation of the occurrence of parasites, except for C. rognoni and C. cirratus absent during in February and March, all other parasite species were present throughout the year in the host fish O. niloticus. Thus, this host is vulnerable at any period. This is consistent with the results of Blahoua et al. (2015) who observed the omnipresence of Tilapia zillii gill parasites in Lake AyaméI. The occurrence of Monogenean presented its maximal prevalence and mean intensity during the rainy seasons (LRS and SRS) with peaks in June and November, respectively. Seasonal variations in Monogenean abundances observed in this study have also been reported by various authors.
For example, Bilong and Tombi (2005) showed that two Monogeneans namely Dactylogyrus in solitus and D. bopeleti gills parasites from Barbus martorelli reached their highest prevalence and their maximum intensity during the rainy seasons. Blahoua et al. (2009) also observed that C. acerbus, C. halli and Scutogyrus minus appeared to be more prevalent and more abundant in long rainy season. Our results can be explained by water temperature which is assumed to be an important factor in controlling the occurrence of Monogeneans (Koskivaara et al., 1991; Simkova et al., 2001). In fact, higher water temperature must have been harmful on the adult worms (Bilong and Tombi, 2005). It also appears from the work of Bilong (1995) that temperature is not the only factor, and that other abiotic factors (concentration of suspended solids, conductivity, water transparency) might influence the seasonal abundance of parasites.
Several authors have investigated the relationship between the level of Monogenean infection and the size of the host fish. Mierzejwska et al. (2006) found that the infection intensities of white bream and roach with Dactylogyrus sphyrna and D. crucifer, respectively, increased with fish body length. Tombi et al. (2014) also agreed with them reporting positive relationship between the prevalence and mean intensity of C. thurstonae, C. halli, C. tilapiae and S. longicornis and the host, O. niloticus. In contrast to these results, Boungou et al. (2008) found that the size of O. niloticus in the dam of Loumbila (Ouagadougou) had no influence on the prevalence of its Monogenean gill parasites. The increase of parasitism with the size of O. niloticus from Ayamé Lake can be attributed to the fact that, larger fish offer large colonized surfaces area to parasites (Cable et al., 2002, Bilong Bilong and Tombi, 2004). In fact, an increase in the branchial surface can provide a larger area of infestation, and thus more potential sites for attachment of these parasites (AydoÄŸdu et al., 2003; Tekin-Ozan et al., 2008). This result can also dueto the strongest water current passing through the gill of larger fish, thus creating convenient conditions for parasites settlements (Guttiérrez and Martorelli, 1999; Turgut et al., 2006).
We found no significant variation on the prevalence and the mean intensity by host sex in Monogenean species. Boungou et al. (2008) had made the same observation with all Monogeneans of O. niloticus, except for C. halli. Tombi et al. (2014) also observed no sex influence on the infestation of the same host gill filaments by C. thurstonae, C. halli and C. tilapiae. This result suggests that very few parasites species have a preference in relation to the sex of the host (Rohde, 1993).
In this study, no significant preferences were found in the distribution of Monogenean species on the gill arches between the left and right sides of its host. These results are similar to those of Le Roux et al. (2011), who found no significant difference between the number of Cichlidogyrus philander on the left and right gill arches of the Pseudocrenilabrus philanderphilander. Similarly, Tombi et al. (2014) reported that C. thurstonae, C. halli, C. tilapiae and Scutogyrus longicornis colonized the two parts of the gill system of the Nile tilapia, O. niloticus in the same way. This could be due to the fact that similar volumes of water flowing through the left and right side of the gill might have brought equal amount of infective larval stages to the gill (Raymond et al., 2006). On the contrary, Hendrix (1990) found an asymmetrical distribution of Bothithrema bothi (Monogenea) on Scophtalmus aquosus (Bothidae). Bilong (1995) interpreted this phenomenon to be linked to Bothidae morphology. For Dessoutter (1992), Bothidae has a flat and dissymmetrical body. According to Rohde (1993) a preference for one side of the host body is associated to body asymmetry of some parasites. This suggests that equitable distribution of parasites on both sides obtained in this study is due to the bilateral symmetry of Nile tilapia associated with that of its gill Monogeneans.
In O. niloticus, the Monogeneans were most preferentially attached to the middle arches II and III. In this same host, Tombi et al. (2014) found that C. halli and S. longicornis were more concentrated on arches II and III, respectively. Le Roux et al. (2011) had reported that Cichlidogyrus philander mostly colonized the median arches of Pseudocrenilabrus philander philander. A number of factors have been recognized as determinants of microhabitat choice among monogeneans. Some authors include intrinsic factors such as the need to increase chances of locating a mate by having a narrow microhabitat niche and environmental conditions. Among the environmental factors, water current has been identified as among the most important factor determining site specificity (Madanire-Moyo et al., 2010; Le Roux et al., 2011) with the suggestion that middle arches offer a hydrodynamically protected site (Lo et al., 2001).
It is an established fact that many parasites have free living stages (eggs or larvae) which have complimentary behavioral reactions to the ambient environment (Echi and Ezenwaji, 2009), and therefore, the settling of oncomiracidia is likely to be influenced by water currents. Indeed, the strongest water current flowing through the middle part of the gill arches and the high respiratory water can provide a high number of parasites (Gutiérrez and Martorelli, 1994; Lo and Morand, 2001). For Buchmann (1989) and Koskivaara and Valtonen (1991), the median preference arches may also be related to the large colonized surfaces they offer to parasites. Other factors such as parasite load (Lo and Morand, 2000) and the host immunity can also influence microhabitat selection (Koskivaara and Valtonen, 1992; Gutiérrez and Martorelli, 1994).
This study adds to a further understanding of Monogeneangill parasitizing O. niloticus in a natural water system. This could lead to more informed aquacultural practises, and maximization of the production of O. niloticus.
The authors have not declared any conflict of interests.
REFERENCES
|
Akoll P, Konecny R, Mwanja WW, Nattabi KJ, Agoe C, Schiemer F (2011). Infection patterns of Nile tilapia (Oreochromis niloticus L.) and African catfish (Clarias gariepinus) in Uganda. Parasitol. Res.110:315-323.
Crossref
|
|
|
|
Aloo PA (2002).A comparative study of helminth parasites from the fish Tilapia zillii and Oreochromis leucostictus in Lake Naivasha and Oloidien Bay, Kenya. J. Helminthol. 76:95-102.
Crossref
|
|
|
|
|
AydoÄŸdu A, Kostadinova A, Fernandez M (2003). Variations in the distribution of Parasites in the common carp, Cpyrinus carpio, from Lake Iznik, Turkey: population dynamics related to season and host size. Helminthologia 40:33-40.
|
|
|
|
|
Bilong Bilong CF (1995). Les Monogènes parasites des poissons d'eau douce du Cameroun: Biodiversité et spécificité; biologie des populations inféodées à Hemichromis fasciatus. Th. Doc. d'Etat, Université de Yaoundé I, P. 341.
|
|
|
|
|
Bilong Bilong CF, Tombi J (2004). Hétérogénéité du système branchial de Barbus martorelli Roman, 1971 (Poisson Cyprinidae) et modèle de croissance. J. Cam. Acad. Sci. 4(3):211-218.
|
|
|
|
|
Bilong Bilong CF, Tombi J (2005). Temporal structure of a component community gill parasites of Barbus martorelli Roman, 1971 (Freshwater Cyprinidae) in the centre province, Cameroon. Cam. J. Biol. Biochem. Sci. 13(1):9-18.
|
|
|
|
|
Bittencourt LS, Pinheiro DA, Cárdenas MQ, Fernandes BM, Tavares-Dias M (2014). Parasites of native Cichlidae populations and invasive Oreochromis niloticus (Linnaeus, 1758) in tributary of Amazon as River (Brazil). Rev. Bras. Parasitol. Vet. 23(1):44-54.
Crossref
|
|
|
|
|
Blahoua KG, N'Douba V, Koné T, Kouassi NJ (2009). Variations saisonnières des indices épidémiologiques de trois Monogènes parasites de Sarotherodon melanotheron (Pisces: Cichlidae) dans le lac d'Ayamé (Côte d'Ivoire). Sci. Nature 6(1):39-47.
Crossref
|
|
|
|
|
Blahoua KG, Yao SS, Etilé RN, N'Douba V (2015). Infection dynamics of four gill Monogenean species from Tilapia zillii (Gervais, 1848) in man-made Lake Ayame I, Côte d'Ivoire. Int. J. Biol. Chem. Sci. 9(1):12-23.
Crossref
|
|
|
|
|
Boungou M, Kabré GB, Marques A, Sawadogo L (2008). Dynamics of population of five parasitic Monogeneans of Oreochromis niloticus Linné, 1757 in the dam of Loumbila and possible interest in intensive pisciculture. Pak. J. Biol. Sci. 11(10):1317-1323.
Crossref
|
|
|
|
|
Buchmann K (1989). Relationship between host size of Anguilla Anguilla and the infection level of the Monogeneans Pseudodactylogyrusspp. J. Fish Biol. 35(4):599-601.
Crossref
|
|
|
|
|
Buchmann K, Lindenstrom T (2002). Interactions between monogenean parasites and their fish hosts. Int. J. Parasitol.32:309-319.
Crossref
|
|
|
|
|
Bush AO, Kevin DL, Jeffrey ML, Allen WS (1997). Parasitology meets ecology on its own terms. J. Parasitol. 83:575-583.
Crossref
|
|
|
|
|
Cable J, Tinsley RC,Harris PD (2002). Survival and embryo development of Gyrodactylus gasterostei (Monogenea: Gyrodactylidae). Parasitology124:53-68..
|
|
|
|
|
Combes C (1995). Interactions durables. Ecologie et évolution du parasitisme. Masson.
|
|
|
|
|
Cone DK (1995). Monogenea (Phylum Platyhelminthes) cited by Woo, PTK (1995) Fish Diseases Disorders.1; Protozoa, metazoan, infections CAP International.
|
|
|
|
|
Dessoutter M (1992). Bothidae. In: Faune des poissons d'eaux douces et saumâtres de l'Afrique de l'Ouest. Tome 2. Levêque C, Paugy D, Teugels, GG (eds) Faune Tropicale, pp. 858-859.
|
|
|
|
|
Dudgeon D, Arthungton AH, Gessner MO, Kawabata Z, Knowloer DJ, Lévêque C, Naimon JR (2006). Freshwater biodiversity: important, threats, status and conservation challenge. Biol. Rev. 81:163-182.
Crossref
|
|
|
|
|
Echi PC, Ezenwaji MG (2009).The parasite fauna of characids (Osteichthyes: Characidae) Anambra river, Nigeria. Afr. J. Ecol. 48:1-4.
Crossref
|
|
|
|
|
El-Seify MA, Zaki MS, Desouky ARY, Abbas HH, Abdel Hady OK, Abou Zaid (2011). Seasonal variations and prevalence of some external parasites affecting freshwater fishes Reared at Upper Egypt. Life Sci. J. 8(3):397-400.
|
|
|
|
|
FAO, Fisheries, Aquaculture Department (2009). The state of world fisheries and aquaculture 2008.Food and agriculture organization of the United Nations, Rome.
|
|
|
|
|
Galli P, Crosa G, Mariniello L, Ortis M, D'Amelio S (2001). Water quality as a determinant of the composition of fish parasite communities. Hydrobiologia 452:173-179.
Crossref
|
|
|
|
|
Ghittino C, Latini M, Agnetti F, Panzieri C, Lauro L, Ciappelloni R, Petracca G (2003). Emerging pathologies in aquaculture: Effects on production and food safety. Vet. Res. Commun. 27:471-479.
Crossref
|
|
|
|
|
Golé Bi GG, Koffi KJD, Dadi SG (2005). Contribution socio-économique de la pêche artisanale en Côte d'Ivoire. FAO/DFID/PMEDP, Projet Pilote Pêche, Kossou. GCP/INT/735/UK, Rapport Expertise, 49p.
|
|
|
|
|
Gourène G, Teugels GG, Hugueny B, Thys Van Audernaerde DFE (1999). Evaluation de la diversité ichtyologique d'un bassin ouest africain après la construction d'un barrage. Cybium 23(2):147-160.
|
|
|
|
|
Gutiérrez PA, Martorelli SR (1994). Seasonality, distribution, and preference sites of Demidospermus valenciennesi Gutiérrez et Suriano, 1992 (Monogenea: Ancyrocephalidae) in catfish. Res. Rev. Parasitol. 54(4):259-261.
|
|
|
|
|
Guttiérrez PA, Martorelli SR (1999).The structure of the monogenean community on the gills of Pimelodus maculates from Río de la Plata (Argentina). Parasitology 119(02):177-182.
Crossref
|
|
|
|
|
Hendrix SS (1990). Attachment and gill microhabitat of Bothitrema bothi (Monogenea). ICOPA VII, Paris, S6. A. Dynamique des populations parasitaires. Abstract of paper.
|
|
|
|
|
Ibrahim MM (2012). Variation in parasite infracommunies of Tilapia zillii in relation to some biotic and abiotic factors. Int. J. Zool. Res. 8(2):59-70.
Crossref
|
|
|
|
|
Johnson SC, Treasurer JW, Bravo S, Nagasawa K, Kabata Z (2004).A review of the impact of parasitic Copepods on marine aquaculture. Zool. Stud. 43(2):229-243.
|
|
|
|
|
Kayis S, Ozceplep T, Capkin E, Altinok I (2009).Protozoan and metazoan parasites of cultured fish in Turkey and their applied treatments. Isr J. Aquacult. Bamid. 61:93-102.
|
|
|
|
|
Kennedy CR (1977). The regulation of fish parasite populations. In: Regulation of parasite populations. G.W. Esh (Ed.). New York: Academic Press P. 253.
Crossref
|
|
|
|
|
Keremah RI, Inko-Tariah MB (2013). Comparative study of ectoparasites on Nile tilapia (Oreochromis niloticus) cultured under integrated and unintegrated pond systems. Afr. J. Biotechnol. 12(19):2711-2714.
|
|
|
|
|
Klemm O, Talbot RW, Klemm KI (1992). Sulfur dioxide in New England Fog, Atmospheric Environment 26, Part A, 2063-2075.
|
|
|
|
|
Koskivaara M, Valtonen ET (1991).Paradiplozoon homoion (Monogenea) and some other gill parasites on roach Rutilus rutilus in Finland. Aqua. Fen. 21:137-143.
|
|
|
|
|
Koskivaara M, Valtonen ET (1992). Dactylogyrus (Monogenea) communities on the gills of roach in three lakes in Central Finland. Parasitology 104:263-272.
Crossref
|
|
|
|
|
Koskivaara M, Valtonen ET, Prost M (1991). Dactylogyrids on the gills of roach in central Finland: features of infection and species composition. Int. J. Parasitol.21:565-572. http://dx.doi.org/10.1016/0020-7519(91)90061-B.
Crossref
|
|
|
|
|
Krasnov BR,Poulin R (2010). Ecological properties of a parasite: species specific stability and geographical variation. In: The biogeography of host-parasite interactions. Morand S, Krasnov BR. New York: Oxford University Press Inc. pp. 99-114.
|
|
|
|
|
Le Roux LE, Avenant-Oldewage A,Walt van der FC (2011). Aspects of the ecology of Cichlidogyrus philander collected from Pseudocrenilabrus philander philander from the Padda Dam, Gauteng, South Africa. Afr. Zool. 46:103-116.
Crossref
|
|
|
|
|
Lizama MAP, Takemoto RM, Ranzani-Paiva MJT, Ayroza LMS, Pavanelli GC (2007).RelaÒ«ãoparasite hospedeiro em peixes de pisciculturas da região de Assis, Estado de São Paulo, Brasil. Piaractus mesopotamicus (Holmberg, 1887). Acta Sci. Biol. Sci. Maringá 29:437-445.
|
|
|
|
|
Lo CM, Morand LS, Galzin R (2001).Gill parasites of Cephalopholis argus (Teleostei: Serranidae) from Moorea (French Polynesia): site selection and coexistence. Folia Parasitol. 48:30-36.
Crossref
|
|
|
|
|
Lo CM, Morand S (2000). Spatial distribution and coexistence of Monogenean gill parasites inhabiting two damselfishes from Moorea Island in French Polynesia. J. Helminthol. 74(4):329-336.
Pubmed
|
|
|
|
|
Lo CM, Morand S (2001). Gill parasites of Cephalopholis argus (Teleostei: Serranidae) from Moorea (French Polynesia): site selection and coexistence. Folia Parasitol. 48:30-36.
Crossref
|
|
|
|
|
Madanire-Moyo GN, Malta MM, Olivier PAS, Luus-Powell WJ (2010). Population dynamics and spatial distribution of Monogeneans on the gill of Oreochromis mossambicus (Peters, 1852) from two lakes of the Limpopo river system, South Africa. J. Helminthol. 27:1-7.
|
|
|
|
|
Malmberg G (1957). On the occurrence of Gyrodactylus on Swedish fishes. In: Swedish, with description of species and a summary in English. Skrifterutgivna av Sodra Sveriges Fiskeriforening: pp. 19-76.
|
|
|
|
|
Mierzejwska K, Wlasow T, Dzika E (2006). Fish Monogeneans from a shallow, eutrophic Oswin lake in Poland. Wiad. Parazytol. 52(1):37-47.
|
|
|
|
|
Morand S, Poulin R, Rohde K, Hayward C (1999).Aggregation and species coexistence of ectoparasites of marine fishes. Int. J. Parasitol. 129:663-672.
Crossref
|
|
|
|
|
Nobah KSC, Koné T, Ouattara NI, Kouamélan EP, N'Douba V, Snoeks J (2008). Etude des performances de croissance de deux tilapias (Tilapia zillii et T. guineensis) et de leurs hybrides encage flottante. Cybium 32(2):131-136.
|
|
|
|
|
Nwani CD, Oti EE, Odoh GE, Ezenwaji HMG (2008). Endo-parasitic helminthes of four Mormyrid species (Osteichthyes : Mormyridae) from a west African flood river system. Pak. J. Biol. Sci. 11(2):202-207.
Crossref
|
|
|
|
|
Ouattara A, Gourène G, Podoor N (2007). Activité photosynthétique du phytoplancton dans le barrage hydroélectrique d'Ayamé I (Côte d'Ivoire). Agron. Afr. 19:63-70.
|
|
|
|
|
Öztürk T, Özer A (2014). Monogenean fish parasites, their host preferences and seasonal distributions in the lower Kızılırmak Delta (Turkey).Turk.J. Fish. Aquat. Sci. 14:367-378.
|
|
|
|
|
Pariselle A (1996). Diversité, spéciation et évolution des monogènes branchiaux de Cichlidae en Afrique de l'ouest. PhD Thesis: University of Montpellier II, France. P. 199.
|
|
|
|
|
Pariselle A, Euzet L (1998). Five new species of Cichlidogyrus (Monogenea: Ancyrocephalidae) from Tilapia brevimanus, T. buttikoferi and T. cessiana from Guinea, Ivory Coast and Sierra Leone (West Africa). Folia Parasitol. 45:275-282.
|
|
|
|
|
Pariselle A, Euzet L (2003). Four new species of Cichlidogyrus (Monogenea: Ancyrocephalidae), gill parasites of Tilapia cabrae (Teleostei: Cichlidae), with discussion on relative length of haptoral sclerites. Folia Parasitol. 50:195-201.
Crossref
|
|
|
|
|
Pariselle A, Euzet L (2009). Systematic revision of dactylogyridean parasites (Monogenea) from cichlid fishes in Africa, the Levant and Madagascar. Zoosystema 31(4):849-898.
Crossref
|
|
|
|
|
Poulin R (1993). The disparity between observed and uniform distributions: a new look at parasite aggregation. Int. J. Parasitol. 23:937-944.
Crossref
|
|
|
|
|
Raymond KM, Chapman LL, Lanciani CA (2006).Host, macrohabitat, and microhabitat specificity in the gill parasite Afrodiplozoon polycotyleus (Monogenea). J. Parasitol. 92:1211-1217.
Crossref
|
|
|
|
|
Reizer C (1967). Aménagement piscicole du Lac d'Ayamé. C.T.F.T., P 46.
|
|
|
|
|
Rohde K (1993). Ecology of marine parasites.An introduction to marine parasitology, 2nd Ed. Cab International, Wallingford, Oxon.
|
|
|
|
|
Rohde K, Heap M, Hayward CJ, Graham KJ (1992).Calitotyle australiensis n.sp.andCalitotyle sp. (Monogenea, Monopisthocotylea) from the rectum and rectal glands and Rugogaster hycholagi Shell, 1973 (Trematoda, Apisdogastrea from the rectal glands of holocephalans off the coast of southeastern Autralia. Syst. Parasitol. 21:69-79.
Crossref
|
|
|
|
|
Sasal P, Morand S, Guegan JF (1997).Parasite species richness for fish of the Mediterranean Sea. Mar. Ecol.-Prog. Ser. 149:61-71.
Crossref
|
|
|
|
|
Shaw DJ, Dobson AP (1995). Patterns of macroparasite abundance and aggregation in wildlife populations: A quantitative review. Parasitology111:S111-S133. http://dx.doi.org/10.1017/S0031182000075855.
Crossref
|
|
|
|
|
Simkova A, Gelnar M, Sasal P (2001). Aggregation of congeneric parasites (Monogenea: Dactylogyrus) among gill microhabitats within one host species (Rutilus rutilus). Parasitology 123:599-607.
Crossref
|
|
|
|
|
Simkova A, Verneau O, Gelnar M, Morand S (2006). Specificity and specialization of congeneric Monogenans parasiting Cyprinid.Evolution60:1023-1037. http://dx.doi.org/10.1554/05-521.1.
Crossref
|
|
|
|
|
Strona G, Stefani F, Galli P (2010). Monogenoidean parasites of Italian marine fish: an updated checklist. Ital. J. Zool.77(4):419-437.http://dx.doi.org/10.1080/11250001003614841.
Crossref
|
|
|
|
|
Sures B (2001). The use of fish parasites as bioindicators of heavy metals in aquatic ecosystems; A Review.Aquat.Ecol. 35:245-255.
Crossref
|
|
|
|
|
Tekin-Özan S, Kir I,Barlas M (2008). Helminth parasites of common carp (Cyprinus carpio L., 1758) in Beysehir lake and population dynamics related to month and host size. Turk.J. Fish. Aquat. Sci. 8:201-205.
|
|
|
|
|
Teugels GG,Thys van den Audenaerde DFE (2003). Cichlidae. In Faune des poissonsd'eaux douces et saumâtres de l'Afrique de l'Ouest,Lévêque C, Paugy D, Teugels GG (eds). Tome 2. Musée Royal d'Afrique Centrale : Tervuren, ORSTOM (Paris), pp.521-600.
|
|
|
|
|
Tombi J, Bilong Bilong CF (2004). Distribution of gill parasites of the freshwater fish Barbus martorelli Roman, 1971 (Teleostei: Cyprinidae) and tendency to inverse intensity evolution between Myxosporidia and Monogenea as a function of the host age. Rev. Elev. Méd. Vét.Pays Trop. 57(1/2):71-76.
|
|
|
|
|
Tombi J, Akoumba JF, Bilong Bilong CF (2014). The monogenean community on the gills of Oreochromis niloticus from Melen fish station in Yaounde, Cameroon. Int. J. Mod. Biol. Res. 2:16-23.
|
|
|
|
|
Tozato HC (2011). Influência do parasitismo na condição de Corydoras aeneus (Gill, 1858) (Osteichthyes: Siluriformes) da bacia do ribeirão do Feijão, São Carlos, SP. Revista científica eletrônica de Medicina Veterinária, Garça/SP, 16:1-15.
|
|
|
|
|
Turgut E, Shinn A, Wootten R (2006). Spatial distribution of Dactylogyrus (Monogenean) on the gills of the host fish.Turk.J.Fish. Aquat.Sci. 6:93-98.
|
|
|
|
|
Velloso AL, Joaber PJ (2010). Influence of ectoparasitism on the welfare of Micropogonias furnieri. Aquaculture310:43-46.
Crossref
|
|
|
|
|
Walter BA, Clayton DH, Cotgreave PC, Gregory RD, Price RD (1995).Sampling effort and parasite species richness.Parasitol.Today11:306-310.
Crossref
|
|
|
|
|
Whittington ID, Cribb BW, Hamwood TE, Halliday JA(2000). Host-specificity of Monogenean (Platyhelminth) parasites: a role for anterior adhesive areas? Int. J.Parasitol. 30:305-320.
Crossref
|
|
|
|
|
Yamada FH, Takemoto RM, Pavanelli GC (2008). Relação entre o fator de condição relativo (Kn) e abundância de ectoparasitos de brânquias, em duas espécies de ciclideos da bacia do rio Paraná, Brasil. Acta Sci. Biol. Sci. 30:213-217.
Crossref
|
|
|
|
|
Zar JH (1996). Biostatistical analysis, 3 Ed. Prentice-Hall Inc.: Upper saddle river New Jersey, P 662.
|
|
|
|
|
Zharikova TI (2000). The adaptative reactions of the gill ectoparasites of the bream (Abramis brama) and the white bream (Blicca bjoerkna) onto the anthropologenic factor influence in the Ivan'kovo reservoir. Parasitology 34(1):50-55.
|
|