ABSTRACT
The dynamics of rangeland vegetation are products of spatial and temporal land use that determine rangeland productivity and conservation of biodiversity. This study examined effects of site, elevation, land use and season on vegetation attributes at three sites in Dida-Hara, southern Ethiopia. Herbaceous plant attributes (that is, above-ground herbaceous biomass, basal cover, plants’ density, species richness, diversity index and evenness), as well as woody plants characteristics such as density and species richness were measured. Herbaceous and woody vegetation variables were examined by season and land use types (that is, enclosures vs. open-grazed areas) across three sites and two altitudes. The results showed that all vegetation attributes were greatly affected by site, land use type and season. Herbaceous vegetation attributes such as biomass, basal cover and herbaceous species richness were more affected by land use types and season. Elevation affected herbaceous vegetation characteristics such as basal cover, herbaceous species diversity and woody richness. Effects of site differences in terms of herbaceous biomass were common during the dry season. Grass diversity was significantly affected by site, elevation, land use type, season and altitude across spatial and temporal scales. Herbaceous biomass was significantly higher in enclosure than in the communal land use type whereas herbaceous biomass showed a declining trend with increased density of woody plants.
Key words: Vegetation attributes, rangeland, grazing pressure, elevation, site, land-use, season.
Knowledge of vegetation dynamics at different spatial and temporal scales in relation to different land use practices, elevation and season is essential to consider for proper management of arid rangelands (Fernandez-Gimenez and Allen-Diaz, 1999; Snyder and Tartowski, 2006). This is particularly important in arid and semi-arid rangelands where forage productivity is highly variable across sites, between seasons and altitudinal gradient (Gibbens et al., 2005; Oba et al., 2000; Briske et al., 2003; Peters and Havstad, 2006). We need to isolate the natural drivers of rangeland vegetation dynamics from human induced impacts (Fernandez-Gimenez and Allen-Diaz, 1999; Bestelmeyer et al., 2006). Among the drivers of rangeland vegetation dynamics, land use practices (Coppock, 1994; Angassa and Oba, 2010), fire suppression (Oba et al., 2000; Angassa and Oba, 2008), site differences and altitudinal variation (Sheuyange et al., 2005; Wu and Archer, 2005; Bestelmeyer, et al., 2006; Dime et al., 2012) and impact of recurrent drought (Peters and Havstad, 2006) can affect the dynamics of rangeland vegetation.
Peters and Havstad (2006) have shown that the spatial distribution of rangeland vegetation across site can be greatly affected by the physical environments such as soil differences, amount and distribution of rainfall. Others (Angassa and Oba, 2010; Teka et al., 2012; Angassa, 2014) indicated that season and land use types have a significant effect on the dynamics of rangeland vegetation. Management of grazing lands thus requires resource planners to be aware of the effects of site differences and land use types on rangeland productivity (Peters and Havstad, 2006; Riginos and Grace, 2008). Wu and Archer (2005) have argued that the extent to which variation among sites may favorably maintain soil moisture and nutrients can be a crucial element of vegetation dynamics. Hence, patterns of vegetation dynamics on rangeland ecological sites can be expressed in terms of climate variability, soil types, soil moisture and nutrient availability (Briske et al., 2003). Although a number of factors might be responsible for the dynamics of rangeland vegetation, the combined effects of site, elevation and land use in relation to season have remained unclear. Thus, how site differences, altitudinal gradient and land use types influence vegetation dynamics on rangelands needs better understanding (Oba et al., 2000; Angassa, 2005).
In the case of arid and semi-arid rangelands, one would envisage, the temporal variability being critically important for herders as they use grazing lands in rotation based on seasonal mobility. This means that the temporal variation acts on the spatial variability of grazing lands to influence the dynamics of land use patterns between seasons of the year (Fernandez-Gimenez and Allen-Diaz, 1999; Bestelmeyer et al., 2006; Snyder and Tartowski, 2006). How gazing or lack of disturbance by livestock influences the products of the interactions between spatial and temporal scales in arid and semi-arid rangelands is important to note. For example, do we have the same outcome as the communal rangelands if livestock grazing is excluded for longer period from a given rangeland site? What would be the responses of vegetation variables if grazing pressure is removed on the one hand, and continuous grazing is allowed on adjacent rangeland areas? Understanding the effects of site, altitudinal gradient and land use types has important implications for proper management and conservation of rangeland biodiversity.
The present study evaluates responses of vegetation attributes to site, altitudinal variation and land use types in Borana rangelands of southern Ethiopia. To achieve the objective of this study, the following questions were asked: (1) How do spatial and temporal variability of rangelands influence herbaceous vegetation attributes (that is, herbaceous biomass, basal cover, density, species richness, diversity and evenness) and woody vegetation variables (that is, density and richness)? (2) What is the effect of land use types, site differences, altitudinal gradient and season on vegetation variables?
It was predicted that the impacts of spatial scales are more positive in facilitating woody vegetation than human induced land use types, while the temporal scales influence herbaceous biomass production, keeping the land use type constant. It was also predicted that the interactions between season and grazing pressure would show greater influence on herbaceous vegetation than on woody vegetation attributes.
Description of the study sites
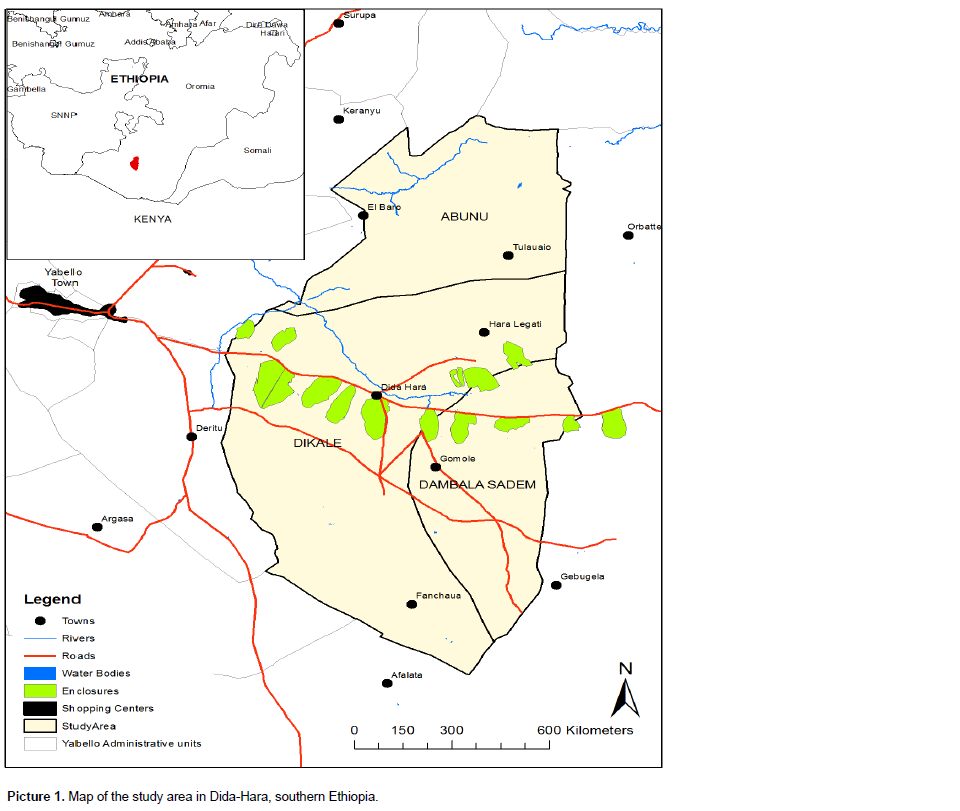
This study was conducted in semi-arid rangelands of southern Ethiopia across three locations (Dikale, Dembi and Siiquu) in Dida-Hara (Picture 1). The study locations hereafter referred to as sites based on two types of land use systems (enclosures vs. communal land use) where cattle are the dominant livestock type. Mean annual rainfall from the nearest weather stations over the last two decades was 500 mm (Angassa and Oba, 2007). Soil types of the three study sites vary from red in the uplands to ‘‘black cotton’’ soils in the bottomlands as in all the three sites, land use types were replicated twice by altitudinal gradient for data collection (Coppock, 1994). Commiphora africana, Acacia drepanolobium and other Acacia species dominate the woody cover, while grass species such as Chrysopogon aucheri, Cenchrus ciliaris, Aristida adscensionis, Eragrostis papposa, Heteropogon contortus, Panicum coloratum, Pennisetum mezianum, Pennisetum stramineum and Themeda triandra make up the majority of herbaceous vegetation in the study sites (Angassa and Baars, 2000).
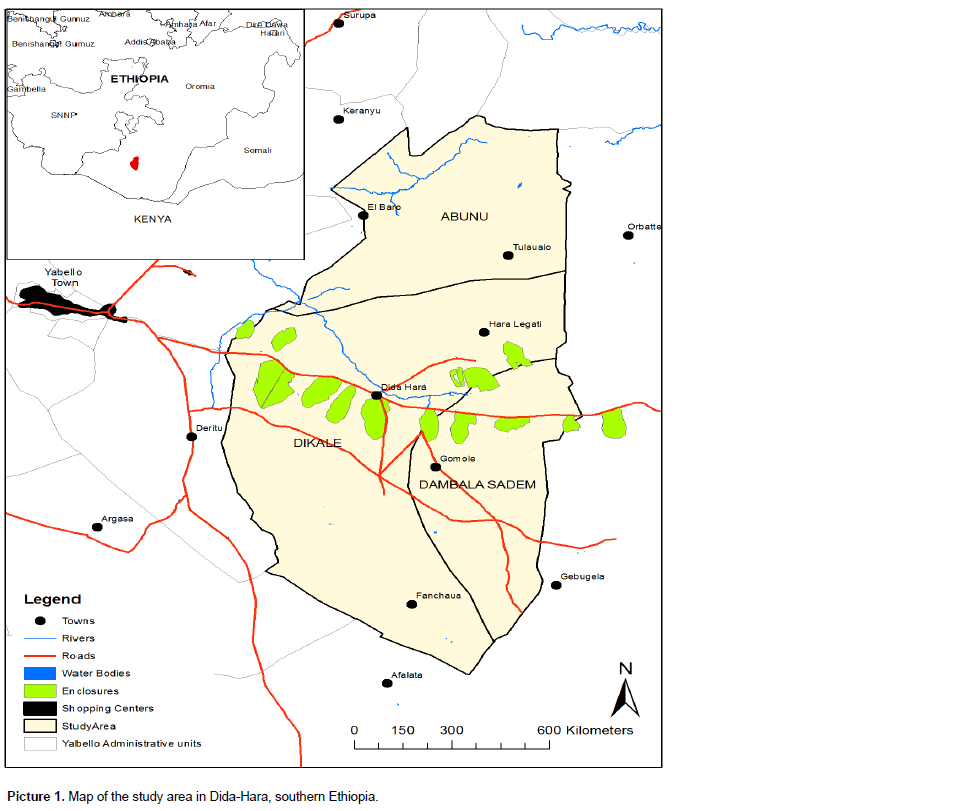
The study questions were examined by conducting vegetation sampling at different altitudes in relation to season and grazing pressure to understand their effects on rangeland vegetation dynamics. According to Peters and Havstad (2006), plot is a patchy spatial unit in association with plant communities. In vegetation samplings, plots are the smallest sampling units nested within landscape patches. The same authors stated that patchy plant communities are often associated with unique microclimate, variation in the physical environments and weather conditions interacting with vegetation community to influence variations among ecological units that also affect plant community success with consequences for rangeland vegetation patterns. Vegetation responses to rangeland ecological variables at broader spatial scales can vary from those influences at intermediate and fine scales (Peters and Havstad, 2006). The second scale of interest in this study was the altitudinal gradient, consisting of groups of patchy mosaics of rangeland units. The scale for the altitude and plot matrixes was assessed based on uplands and bottomlands. The third scale used for the evaluation of rangeland vegetation dynamics was land use types or grazing pressure (that is, enclosures vs. communal rangelands) replicated across two elevation ranges. The fourth independent variable was season of rainfall as a factor regulating plant productivity on the rangelands.
Sampling procedure
In each study location, two land-use types (that is, enclosures vs. communal grazing lands) were identified. Enclosures are areas protected from livestock grazing to conserve pasture for calves for dry season grazing, while communal grazing lands are continuously grazed areas throughout the year and lacks rest for plant species to recover. These land-use types were replicated twice in both upland and bottom land altitudes for sampling vegetation in enclosures vs. communal rangelands by altitude. Upland altitudes are areas > 1500 m above sea level (m.a.s.l.), while area < 1500 m.a.s.l was considered as bottomlands. A 500 m long transect was established at each site in each land-use system per altitude gradient. The 500 m long transect was divided into 10 units consisting of 10 plots at 50 m intervals between each plot. Vegetation variables were sampled in each plot along the transect within each land-use system per altitude using 1 m × 1 m plot to quantify herbaceous vegetation, whereas 10 m × 10 m plot was used for sampling woody plants. During two rainy seasons and one dry season (that is, in May and November, 2004, and February, 2005, respectively), biomass, basal cover, species richness, diversity and evenness of herbaceous plants, as well as density and richness of woody plants were measured. Plant species were identified and counted in each plot. Herbaceous biomass was harvested using hand cutting and samples were oven dried at 65°C for 24 h. Herbaceous biomass was determined based on the dry matter estimation of herbaceous samples. The proportion of grass basal cover was estimated visually based on the area (soil part) covered by a grass base compared with bare ground (Angassa and Oba, 2010). The density of woody species were determined by counting the total number of individuals in plots of 10 m × 10 m. Species richness were counted per plot per census (that is, the total count of all species per plot), while species diversity index, H´ = -(Σ (pilnpi)) (Shannon, 1948) was calculated using the relative abundance of each plant species (where pi represents the proportion of individual species in each plot).
Data analysis
SAS statistical software version 9.1 (SAS Institute, 2001) was used for statistical analysis. Analysis of variance (ANOVA) was used to evaluate vegetation attributes (that is, biomass, basal cover, density, species richness, diversity index and evenness of herbaceous plants) and woody vegetation variables (woody density and richness) in response to spatial scales in rangeland ecological units. Post-hoc comparisons of significant differences were done using the least square difference (LSD) method. The relationship between woody density and grass biomass were also determined using a polynomial regression. Statistical significance was reported at P< 0.05.
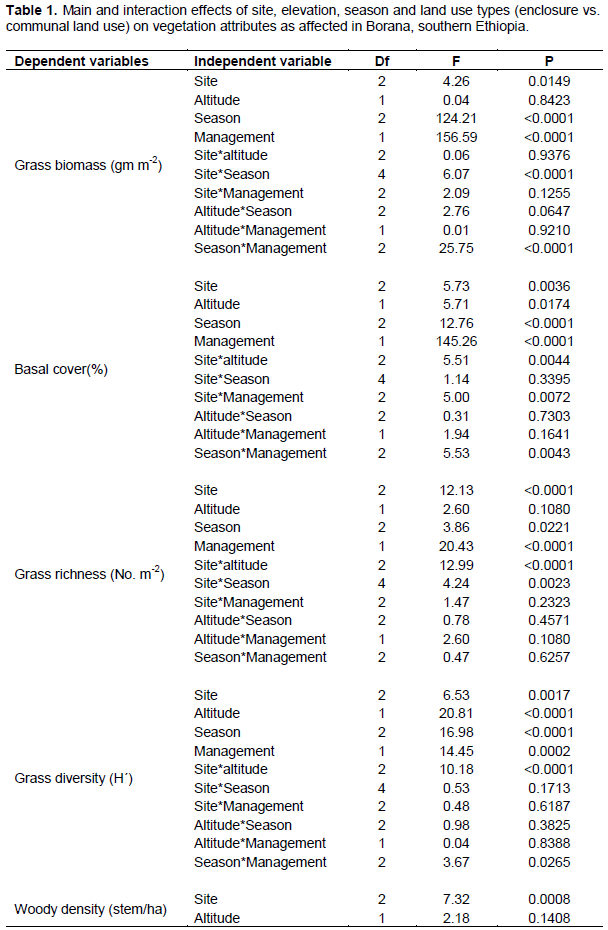
Site, elevation and land use types on vegetation attributes
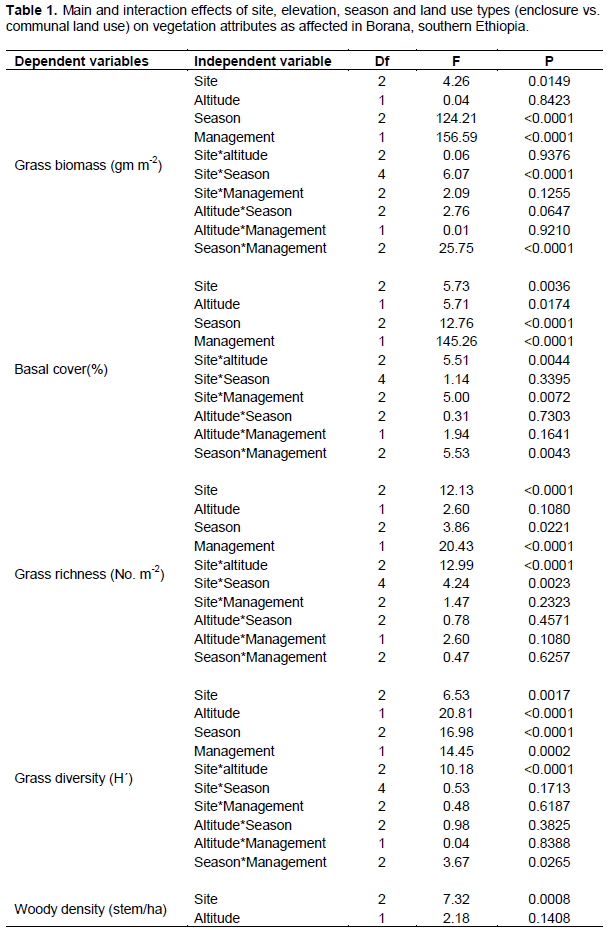
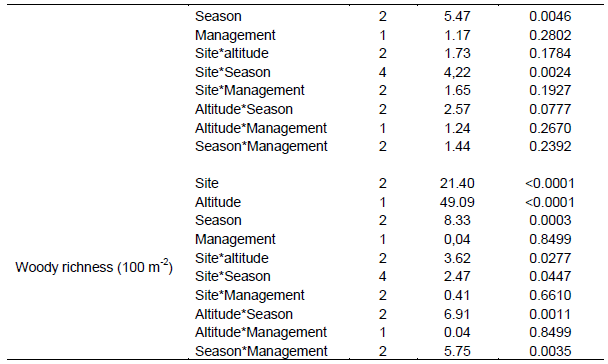
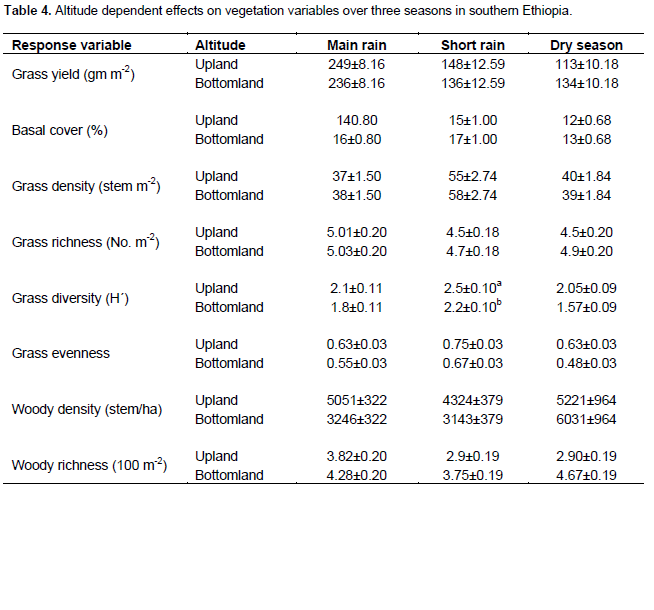
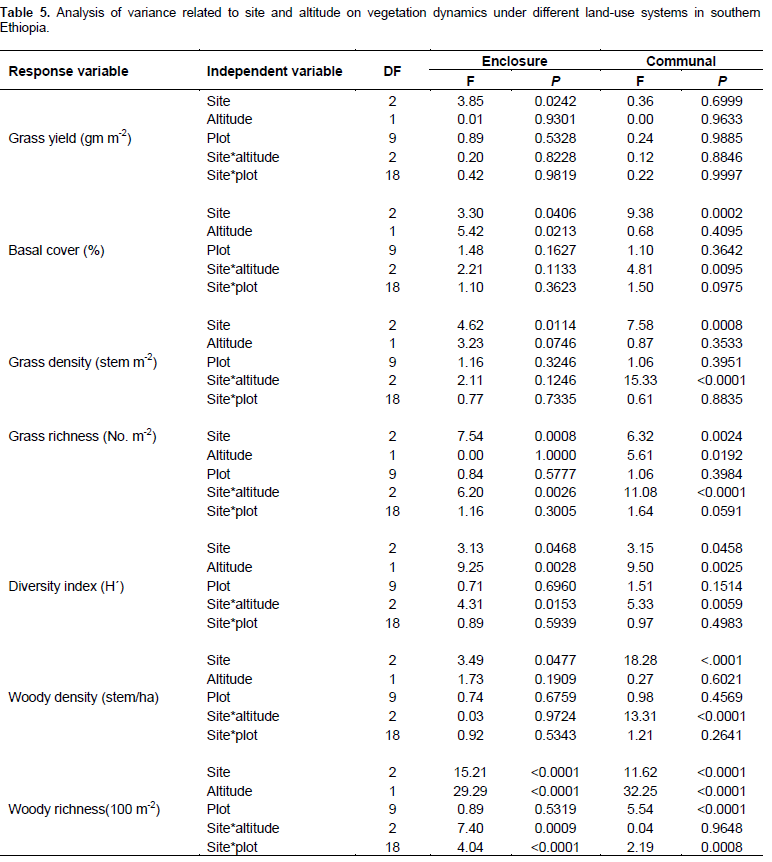
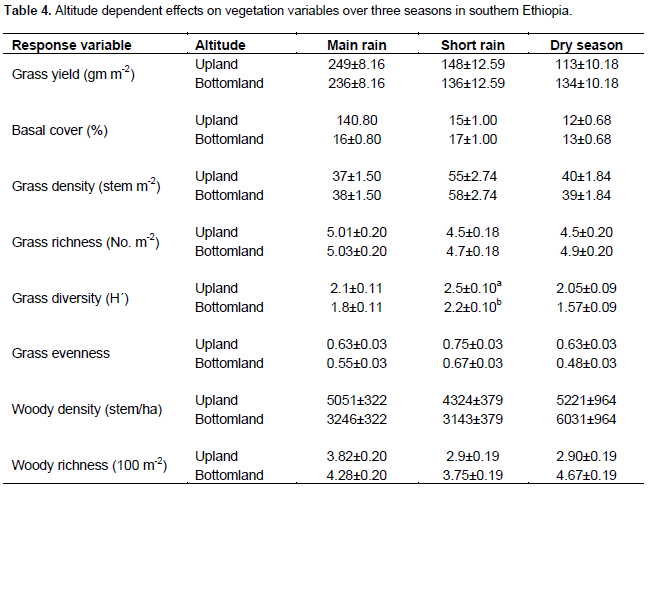
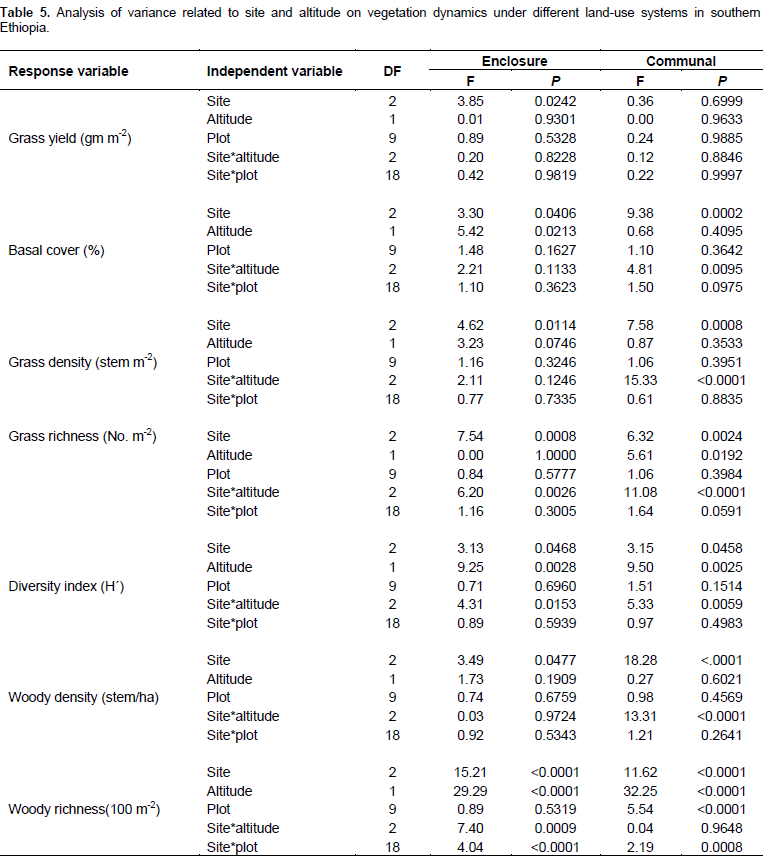
The present results showed that all vegetation variables were significantly related to site (P < 0.05) (Table 1). Season and land use types were significantly related to most herbaceous vegetation variables (P < 0.001). Interactions between site and season, as well as season and management (that is, land use types) on the yield of grasses were significant (P < 0.001) (Table 1). Grass basal cover, herbaceous diversity and woody richness were significantly related to the effects of altitudinal gradient (Table 1). There were significant interactions between site and altitude on grass basal cover, herbaceous species richness and diversity, and woody richness (Table 1). The interaction between season and altitude was significantly related to woody richness (Table 1).
Season
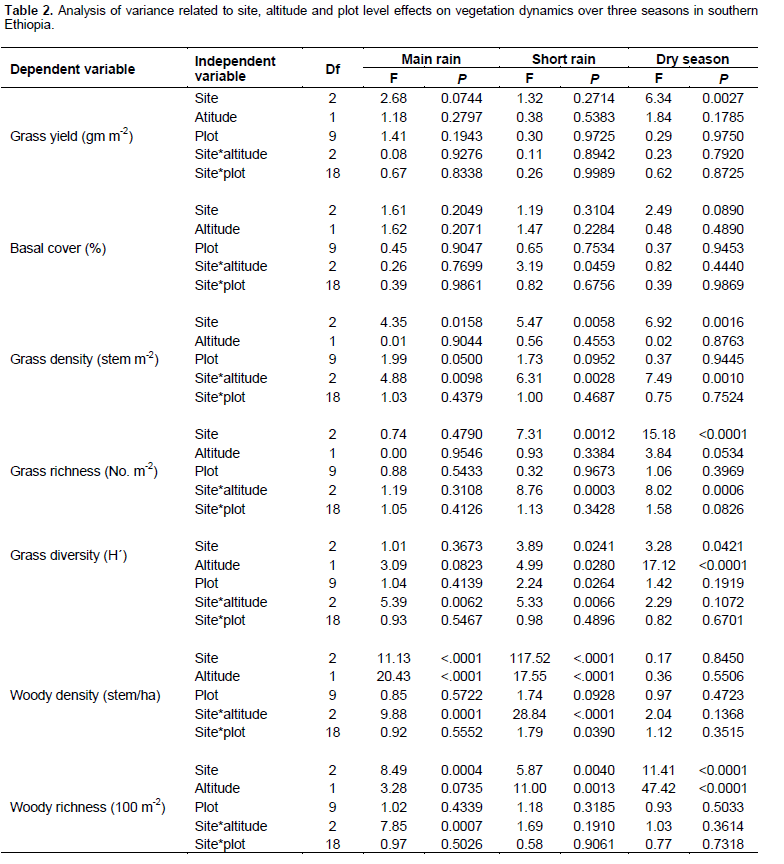
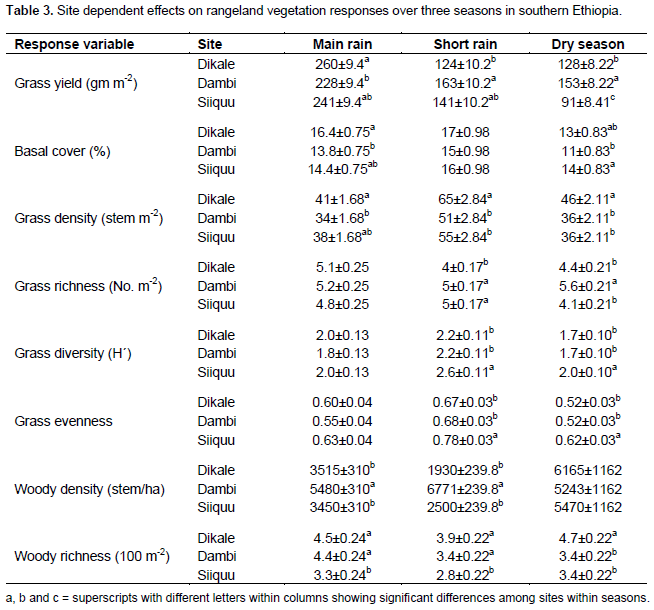
No significant variation in grass yield was recorded between the study sites and interactions between site and season (Table 2). However, site was significantly (P < 0.05) related to the yield of grasses during the dry season (Tables 2 and 3). The interaction between site and altitude also significantly (P < 0.05) affected the basal cover of grasses during the short rainy season (Table 2). Grass density was significantly (P < 0.05) differed between sites despite seasonal variation (Table 2). Likewise, grass density was significantly (P < 0.05) affected by the interaction between site and altitude during the different seasons (Table 2). Site had a significant effect on grass richness only during the short and dry seasons.
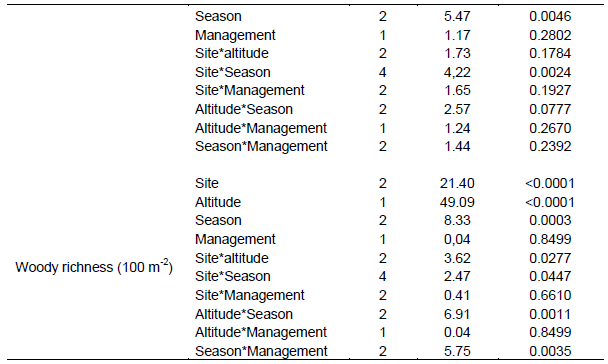
Grass diversity was significantly affected at all spatial scales and their interaction (exception being the interaction between site and plot) during the short rainy season (Table 2). Hence, our results showed that the interaction between site and landscape had a significant effect on grass diversity during the main rainy season (Table 2). Moreover, grass diversity was significantly affected by site and landscape, as well as the interaction between site and landscape during the dry season (Tables 2 and 4). Woody plant density was significantly affected by site, landscape and their interaction during the main and short rainy seasons (Tables 2 and 3). Similarly, the interaction between site and plot had a significant effect on the density of woody plants during the short rainy season (Table 2). Woody plant richness was significantly affected by site and the interaction between site and landscape during the main rainy season. Overall, site and landscape had a significant effect on woody plant richness during the short rainy and dry seasons.
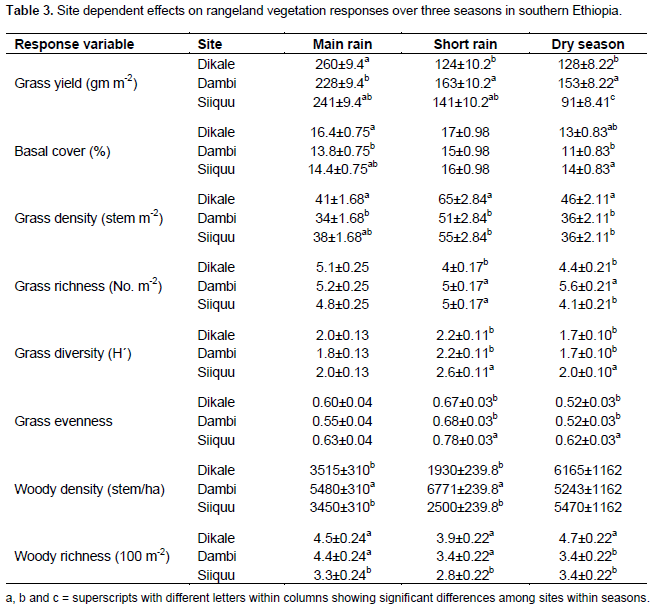
Land-use
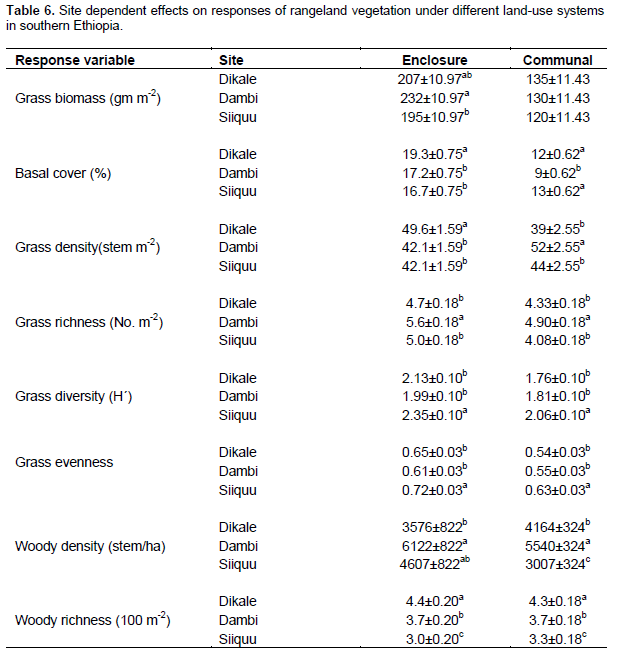
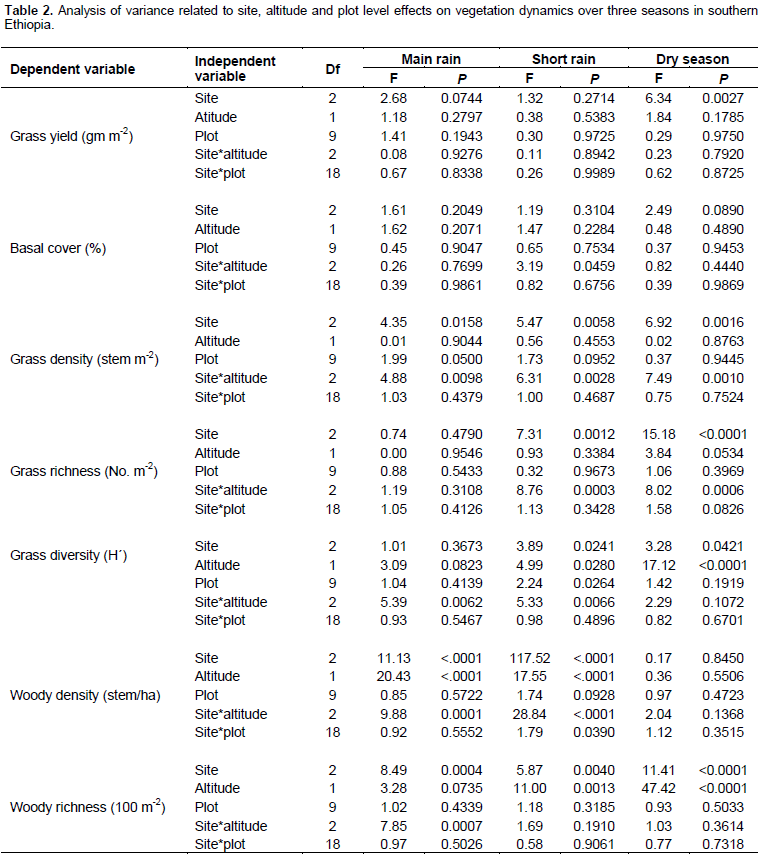
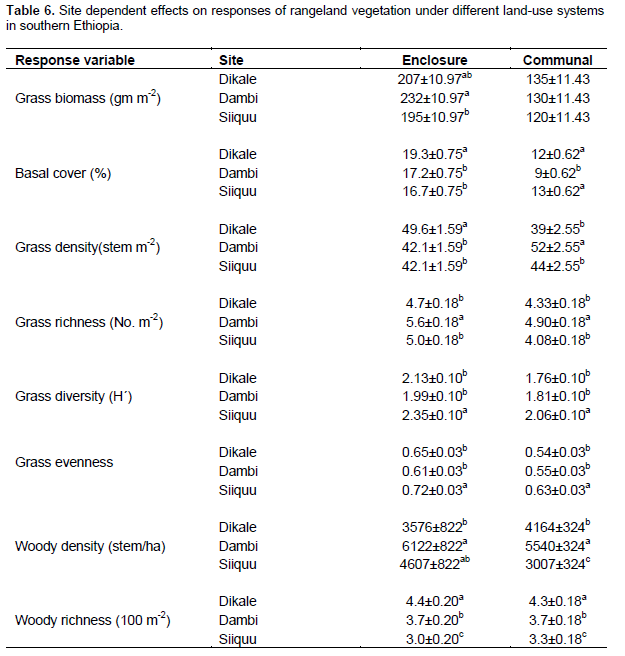
Grass biomass was significantly (P < 0.05) affected by site differences under the enclosure (Tables 5 and 6). However, the interaction effects were non-significant (P > 0.05) at altitude and plot levels when considering the yield of grasses under both land-use systems (Table 5). The basal cover of grasses showed significant differences both at site and altitude levels under enclosure management (Tables 5 and 7). The basal cover of grasses was significantly affected by site, and the interaction between site and altitude under communal land-use (Table 5). Site differences had a significant effect on the density of grasses under both land-use systems, while the interaction between site and landscape was only significant under the communal land-use (Table 5). Site differences significantly affected grass richness under both land-use systems (Tables 5 and 6). Nevertheless, the effect of altitude on grass richness was only significant under the communal land-use. The interaction between site and altitude had a significant effect on grass richness under both land-use systems.
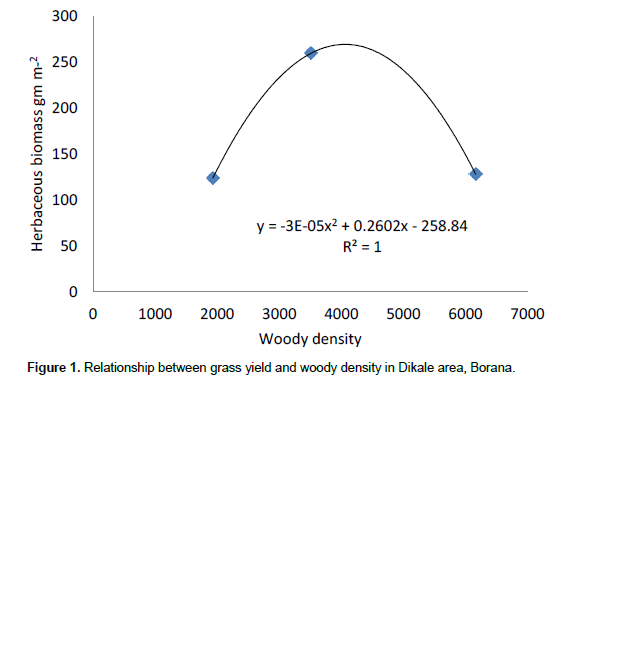
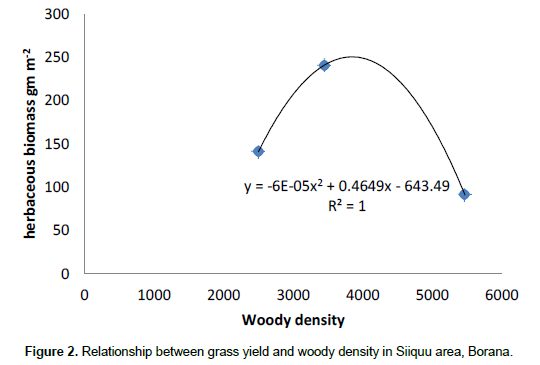
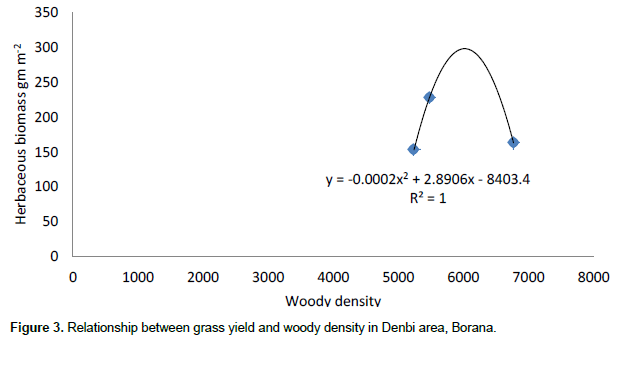
The results showed that grass diversity was significantly affected by site, altitudinal variation and their interaction under both land-use systems. Woody plant density significantly varied which was caused by site differences under both land-use systems (Tables 5 and 6). The interaction between site and landscape under both land-use systems was also significantly (P < 0.05) different. The current result showed that site differences had a highly significant effect on the density of woody vegetation attributes under the communal land-use system. Woody plant richness was significantly (P < 0.05) affected by site, altitude and the interaction between site and plot under both land-use systems. Overall, woody vegetation richness was significantly affected by the interaction between site and altitude under the enclosure management (Table 5). The relationship between woody density and grass yield is shown in Figures 1, 2 and 3. The present results showed that woody plant density was inversely related to grass cover in all study sites (Figures 1, 2 and 3).
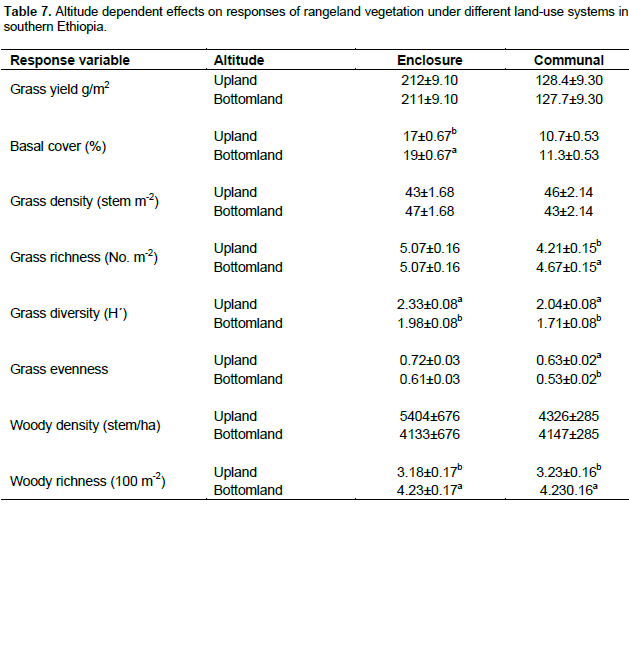
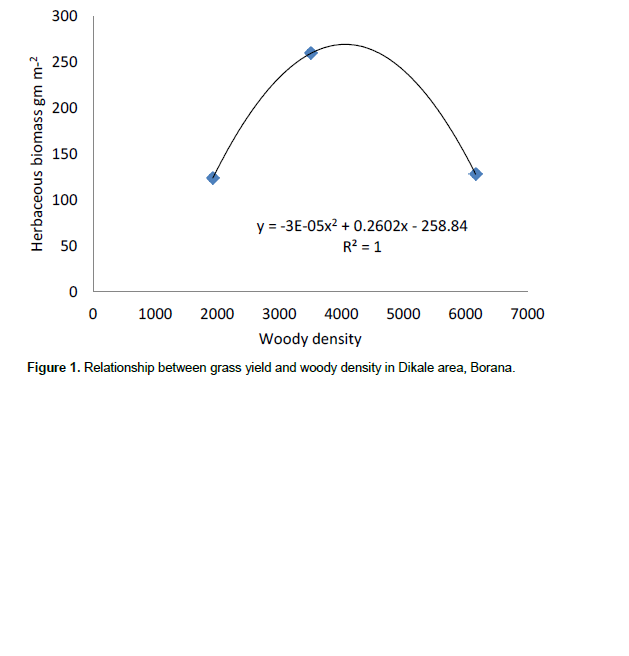
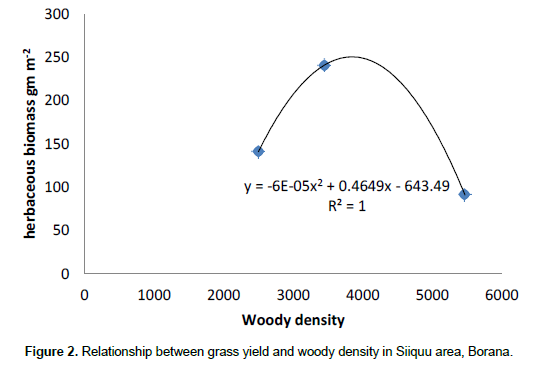
Effects of site, elevation and land use on vegetation attributes
It was predicted that the impacts of spatial scales are more decisive in shaping the distribution of vegetation than management intervention in arid rangelands, while the temporal scales could influence vegetation production, keeping land use type constant. Such impacts were observed in terms of woody vegetation composition, while herbaceous biomass was strictly controlled by grazing pressure and season. Oba et al. (2003) suggested that rangeland spatial variability would tend to increase differences in vegetation composition and production. The findings of the present results indicate that vegetation responses could be influenced by the spatial and temporal variability of arid rangelands. Oba et al. (2000) have also indicated that landscape heterogeneity and temporal scales would contribute to the spatial variability of rangeland vegetation. The current results also support this argument, as the observed vegetation variables greatly varied across spatial and temporal scales. Similarly, several authors, for example, Meentemeyer and Box, (1987), Turner et al. (1989), Oba et al. (2003), Peters and Havstad (2006), Snyder and Tartowski (2006) and Wu and Archer (2005) have argued that vegetation dynamics are more responsive to site differences and altitudinal gradient than land use types.
The present study found that yield of grasses during the dry season varied between sites. The findings of the present study show that the interaction between site and landscape has important effect on basal cover of grasses during the short rainy season. Generally, the current findings suggest that variation in rangeland sites has greatly influenced the density of grasses regardless of seasonal variability. Vegetation composition and productivity within a region basically reflect the existing climate in terms of season and annual rainfall variability (Fernandez-Gimenez and Allen-Diaz, 1999; Oba et al., 2000; Peters and Havstad, 2006). Ultimately, climate variability plays a central role in determining the dynamics of rangeland vegetation (Ellis and Swift, 1988; Fernandez-Gimenez and Allen-Diaz, 1999; Briske et al., 2003). However, regional climatic variability may not account for the spatial patterns of vegetation at local level as a result of the irregularities that could arise due to altitudinal variation (Fernandez-Gimenezand and Allen-Diaz, 1999; Oba et al., 2000; Briske et al., 2003; Wu and Archer, 2005; Peters and Havstad, 2006). Hence, landscape spatial scales might exert a strong influence on the distribution, growth and abundance of plant communities over a wider altitudinal range due to substantial dissimilarities, in elevation, soil texture and moisture conditions, and nutrient supply (Wu and Archer, 2005). For example, Peters and Havstad (2006) have indicated that the nutrient and soil moisture distribution between high and low resource areas could be among the major factors for the variation in vegetation composition across spatial scales. Similarly, the results of the present study indicate that grass diversity was highly influenced by differences in site, altitude, season of rainfall. This is in accordance with the existing information that suggests that the dynamics of rangeland vegetation can be non-linear (Peters and Havstad, 2006) and locally mediated by the type of topography of an area (for example, Wu and Archer, 2005). These scale-dependent effects can have important implications for the local users and decision makers on the sustainability of arid land management.
Previous studies (for example, Ryerson and Parmenter, 2001; Briske et al., 2003) have shown that the occurrence of vegetation type along altitudinal gradient is independent of the exclusion of herbivores from a specific land site. The current study shows that grass density was greatly affected by the interaction between site and elevation during the different seasons, suggesting that plant composition always varies across landscapes. In addition to distinct and interactive effects of spatial scales, environmental variables such as season and land-use type are among the most important factors in driving vegetation dynamics (Peters and Havstad, 2006; Briske et al., 2003). In some locations, grasses are diminished prior to drought, while in others grasses remain high regardless of the occurrence of drought (Peters and Havstad, 2006). Similarly, Coppock (1994) has reported that intensive grazing can lead to reduced grass cover and increased bush encroachment. On the other hand, exclusion of livestock grazing is found ineffective in limiting the spread of bush encroachment in semi-arid savannas (Peters and Havstad, 2006; Angassa and Oba, 2010).
According to Kerstin et al. (2005), increase in the density of woody encroachment suppresses grass production. From the results of the present study it seems that as the density of woody species increased grass biomass tends to reduce. In the present study, heavy grazing pressure is a common feature of the study sites.
It seems that the dynamics of rangeland vegetation that potentially accompany shifts in season or disturbance regimes is likely to be controlled by altitudinal gradient. Knowledge of vegetation dynamics may therefore need to explicitly account for the spatial and temporal variability of rangeland ecological sites in terms of season and land-use types to precisely predict the drivers of vegetation changes in savanna ecosystems.
The authors have not declared any conflict of interests.
The author thanks the financial support from a NUFU grant in collaboration with Hawassa University. The author also thanks Katelo Guyo for his assistance during fieldwork.
REFERENCES
|
Angassa A, Baars RMT (2000). Ecological condition of encroached and non-encroached rangelands in Borana, Ethiopia. Afr. J. Ecol. 38:321-328.
Crossref
|
|
|
|
Angassa A (2005). The ecological impact of bush encroachment on the yield of grasses in Borana rangeland ecosystem. Afr. J. Ecol. 43:14-20.
Crossref
|
|
|
|
|
Angassa A, Oba G (2007). Relating long-term rainfall variability to cattle population dynamics in communal rangelands and a government ranch in southern Ethiopia. Agric. Syst. 94:715-725.
Crossref
|
|
|
|
|
Angassa A, Oba G (2008). Herder perceptions on impact of range enclosures, crop farming, fire ban and bush encroachment on the rangelands of Borana, southern Ethiopia. Hum. Ecol. 36:201-215.
Crossref
|
|
|
|
|
Angassa A, Oba G (2010). Effects of grazing pressure, age of enclosures and seasonality on bush cover dynamics and vegetation composition in southern Ethiopia. J. Arid Environ. 74:111-120.
Crossref
|
|
|
|
|
Angassa A (2014). Effects of grazing intensity and bush encroachment on herbaceous species and rangeland condition in southern Ethiopia. Land Degrad. Dev. 25:438-451.
Crossref
|
|
|
|
|
Bestelmeyer BT, Trujillo DA, Tugel AJ, Havstad KM (2006). A multi-scale classiï¬cation of vegetation dynamics in arid lands: What is the right scale for models, monitoring, and restoration? J. Arid Environ. 65:296-318.
Crossref
|
|
|
|
|
Bestelmeyer BT, Judy PW, Havstad KM (2006). Soil-geomorphic heterogeneity governs patchy vegetation dynamics at an arid ecotone. Ecology 87:963-973.
Crossref
|
|
|
|
|
Briske DD, Fuhlendorf SD, Smeins FE (2003). Vegetation dynamics on rangelands: A critique of the current paradigms. J. Appl. Ecol. 40:601-614.
Crossref
|
|
|
|
|
Coppock DL (1994). The Borana plateau of southern Ethiopia: synthesis of pastoral research development and changes, 1980-90. ILCA (International Livestock Centre for Africa): Addis Ababa, Ethiopia.
|
|
|
|
|
Dime TT, Angassa A, Nemomissa S, Woldemariam T (2012). Savanna Land Use and its Effect on Woody Plant Species Diversity in Borana, Southern Ethiopia. Star J. Sci. Technol. Arts Res. 1(2):43-52.
Crossref
|
|
|
|
|
Ellis EJ, Swift MD (1988). Stability of African pastoral ecosystems: Alternate paradigms and implications for development. J. Range Manag. 41:450-459.
Crossref
|
|
|
|
|
Fernandez-Gimenez ME, Allen-Diaz B (1999). Testing a non-equilibrium model of rangeland vegetation dynamics in Mongolia. J. Appl. Ecol. 36:871-885.
Crossref
|
|
|
|
|
Gibbens RP, Mcneely RP, Havstad KM, Beck RF, Nolen B (2005). Vegetation change in the Jornada Basin from 1858 to 1998. J. Arid Environ. 61:651-658.
Crossref
|
|
|
|
|
Kerstin W, David W, David S (2005). Multi-scale patterns and bush encroachment in an arid savanna with a shallow soil layer. J. Veg. Sci. 16:311-320.
Crossref
|
|
|
|
|
Meentemeyer V, Box EO (1987). Scale effects in landscape studies. In: Landscape heterogeneity and disturbance. Edited by Turner MG, Springer-Verlag,New York. pp. 15-34.
Crossref
|
|
|
|
|
Oba G, Post E, Syvertsen PO, Stenseth NC (2000). Bush cover and range condition assessments in relation to landscape and grazing in southern Ethiopia. Landsc. Ecol. 15:535-547.
Crossref
|
|
|
|
|
Oba G, Weladji RB, Lusigi WJ, Stenseth NC (2003). Scale-dependent effects of grazing on rangeland degradation in northern Kenya: A test of equilibrium and non-equilibrium hypotheses. Land Degrad. Dev. 14:83-94.
Crossref
|
|
|
|
|
Peters DPC, Havstad KM (2006). Nonlinear dynamics in arid and semi-arid systems: Interactions among drivers and processes across scales. J. Arid Environ. 65:196-206.
Crossref
|
|
|
|
|
Riginos C, Grace JB (2008). Savanna tree density, herbivores, and the herbaceous community: bottom-up vs. top-down effects. Ecology 89:2228-2238.
Crossref
|
|
|
|
|
Ryerson DR, Parmenter RR (2001). Vegetation change following removal of keystone herbivores from desert grasslands in New Mexico. J. Veg. Sci. 12:167-180.
Crossref
|
|
|
|
|
SAS Institute (2001). Statistical Analysis System/GLM Software. Institute Inc.: Cary, NC.
|
|
|
|
|
Shannon CE (1948). A mathematical theory of communication. Bell Syst. Technol. J. 27:379-423.
Crossref
|
|
|
|
|
Sheuyange A, Oba G, Weladji RB (2005) Effects of anthropogenic fire history on savanna vegetation in northeastern Namibia. J. Environ. Manag. 75:189-198.
Crossref
|
|
|
|
|
Snyder KA, Tartowski SL (2006) Multi-scale temporal variation in water availability: Implications for vegetation dynamics in arid and semi-arid ecosystems. J. Arid Environ. 65:219-234.
Crossref
|
|
|
|
|
Teka H, Madakadze CI, Angassa A, Hassen A (2012). Effect of Seasonal Variation on the Nutritional Quality of Key Herbaceous in Semi-arid Areas of Borana, Ethiopia. Indian J. Anim. Nutr. 29(4):324-332.
|
|
|
|
|
Turner MG, Costanza R, Sklar FH (1989). Methods to evaluate the performance of spatial simulation models. Ecol. Model. 48:1-18.
Crossref
|
|
|
|
|
Wu XB, Archer SR (2005). Scale-dependent influence of topography-based hydrologic features on vegetation patterns in savanna landscapes. Landsc. Ecol. 20:733-742.
Crossref
|
|