Full Length Research Paper

ABSTRACT
Soil of the former lake Texcoco is nitrogen (N) depleted, so any attempt to vegetate the area will require the application of an N fertilizer. Urea is commonly used as fertilizer, but its application to soil might affect emissions of greenhouse gases (GHG), such as carbon dioxide (CO2), nitrous oxide (N2O) and methane (CH4), and the high pH and electrolytic conductivity (EC) in the Texcoco soil might inhibit the hydrolysis of urea. Four soils of the former lake bed with EC 3.3, 88.3, 96.9, and 121 dS m-1, were amended with urea while dynamics of mineral N and emissions of GHG were monitored. Urea increased emission of CO2 in all soils and emission of N2O in soil with EC≤88.3 dS m-1, but emission of CH4 was not affected. Hydrolysis of urea occurred in all soils although it was significantly lower in soil with EC≥88.3 dS m-1. Oxidation of NH4+ occurred in soil with EC≤96.9 dS m-1, but oxidation of NO2- only in soil with EC 3.3 dS m-1. It was found that oxidation of NH4+ and NO2-, and hydrolysis of urea was inhibited by the high EC in soil of the former lake bed, while emissions of CO2 and N2O, but not CH4 were affected by application of urea.
Key words: Carbon dioxide (CO2), nitrous oxide (N2O) and methane (CH4), former lake bed, mineral N in soil.
INTRODUCTION
The former lake of Texcoco located in the valley of Mexico City (Mexico) at an altitude of 2240 m above sea level with a mean annual temperature of 16°C and annual precipitation of 705 mm was drained from the 17th century onwards to avoid flooding in Mexico City (O’Hara and Metcalfe, 1997). The drainage of the lake left a soil with a high Ph and salinity and little vegetation (Dendooven et al., 2010). During the dry season, the wind erosion was high. However, during the rainy season, flooding occurred frequently. The groundwater, which is highly alkaline and saline, was just under the soil surface and after heavy rainfall the area flooded. The national water authority (Commission Nacional de Agua, CNA) installed drainage pipes so that the area could be vegetated to stop wind erosion and dust storms in Mexico City (Luna-Guido et al., 2000). However, previous research showed that the soil of the former lake Texcoco is N depleted (Conde et al., 2005). It can be speculated that the high pH and salinity inhibits N2 fixing microorganisms thereby limiting the amount of mineral N that enters the soil (Barua et al., 2011; Welsh et al., 2007).
Urea is cheap and often applied to fertilize crops. It could easily be applied to grass, shrubs and trees that might be used to vegetate the former lake bed. Hydrolysis of urea can occur in three ways, biotic (Burton and Prosser, 2001), abiotic in soil with a high pH (Ghandi and Paliwal, 1976) and abiotic through extracellular ureases (Conrad, 1996) generating two NH3 molecules. As such, concentration of NH4+ will increase in urea-amended soil (Burton and Prosser, 2001). However, if most of the process is biological then the high pH and salinity might inhibit the release of NH4+. Additionally, application of urea is known to increase emissions of nitrous oxide (N2O) and might affect emissions of carbon dioxide (CO2) and methane (CH4) (Wang et al., 2011). Hydrolysis of urea will generate ammonium ions that can be oxidized by nitrifiers, first to NO2- and subsequently to NO3-. However, high electrolytic conductivity (EC) is known to inhibit the activity of nitrifiers so emissions of N2O after application of urea might not increase (Zhu et al., 2011). Therefore, four soils of the former lake Texcoco with different pH and EC were amended with urea, with or without acetylene (C2H2), known to inhibit nitrification (Bateman and Baggs, 2005), while emissions of CO2, CH4 and N2O were monitored in an aerobic incubation. Acetylene was applied to half of the soil samples so that the importance of the nitrification process in the emissions of N2O could be determined. The objective of this study was to investigate the effect of high alkalinity and salinity on hydrolysis of urea, emissions of GHG and dynamics of mineral N (ammonium (NH4+), nitrite (NO2-) or nitrate (NO3-)).
MATERIALS AND METHODS
Soil sampling
Details of the soil of the former lake Texcoco can be found in Dendooven et al. (2010) and mineralogy in Gutiérrez-Castorena et al. (2005). Part of the former lake bed has been drained and irrigated with sewage effluent from a waste water plant to remove excess of salt (Luna-Guido et al., 2000). For instance, the concentration of the sodium ions (Na+) decreased from 21 g kg-1 dry soil to 3 g kg-1 dry soil after 8 years of flooding and the chloride ions (Cl-) from 21 g kg-1 dry soil to undetectable amounts (Luna-Guido et al., 2000). The Distichlis spicata, an indigenous grass with a high tolerance to salt and Na+ and tamarix (Tamarix species) have been introduced since the early 1970s to control erosion, and they now cover much of the area. More details on the vegetation and the effluents used to drain the plots can be found in Luna-Guido et al. (2000).
At four locations with different EC and pH due to different periods of drainage, five approximately 400 m2 plots were defined and sampled by augering the 0 to 15 cm layer 30-times with a stony soil auger with diameter 7 cm (Eijkelkamp, Nl) at random on 7 March 2011. The 30 soil samples taken from each site (n = 4) and plot (n = 5) were pooled, 5 mm sieved and characterized (Table 1). As such, 20 soil samples were obtained. Details of the sampled soils can be found in Table 1. The soil with an EC 3.3 dS m-1 was denoted soil A, with EC 88.3 dS m-1 soil B, with EC 96.9 dS m-1 soil C and with EC 121 dS m-1 soil D. This field based replication was maintained in the incubation study.
Treatments and experimental set-up
The experimental design was a completely randomized 2 ´ 4 factorial with five replications (maintained from the field site replications for each soil). The factors were four soil types and four soil amendments which were: 1) 200 mg N kg-1 applied as urea; 2) 200 mg N kg-1 applied as urea plus acetylene (C2H2) at 0.1%; 3) unamended soil; and 4) unamended soil plus C2H2 at 0.1%.
Sixteen sub-samples of 10 g dry soil from each of the four soils and five sampled plots were added to 120 ml flasks. Eight soil samples were adjusted to 40% water holding capacity (WHC) by adding distilled water and eight by adding an urea solution. The flasks were air-tight sealed with a Suba-seal. Four of the flasks amended with distilled water and four with the urea solution were injected with 0.1 ml acetylene (C2H2) to inhibit nitrification (Bateman and Baggs, 2005). Additionally, 15 flasks without soil were air-tight sealed and incubated in the same way to determine the concentration of CO2, N2O and CH4 in the atmosphere. The flasks were incubated in the dark at 25 ± 2°C. After 0, 1, 3 and 7 days, one flask was selected from each soil and treatment at random and the headspace was analyzed for CO2, CH4 and N2O. Additionally, three flasks without soil were selected at the same time and the headspace analyzed. The flasks were opened, the soil removed and extracted for mineral N (ammonium (NH4+), nitrate (NO3-) and nitrite (NO2-)) with 0.5 M K2SO4.
Chemical analyses
Details of the techniques used to measure WHC, pH, EC, total N and soil particle size distribution can be found in Ruíz-Valdiviezo et al. (2010). The extracted NH4+, NO3- and NO2- were measured colourimetrically with a San Plus System-SKALAR automatic analyzer (Breda, The Netherlands).
The headspace of the vials was analyzed for CO2 and N2O on an Agilent Technologies 4890D gas chromatograph fitted with an electron capture detector (ECD) and CH4 on an Agilent Technologies 4890D gas chromatograph fitted with a flame ionization detector (FID). Details of the columns used, gas flow, and oven, detector and injector temperatures can be found in Ruíz-Valdiviezo et al. (2010). Concentrations of CO2, N2O and CH4 were calculated by comparing peak areas against a standard curve prepared from known concentrations, 10 ppm N2O in N2, 5 ppm CH4 in N2 and 2500, 20000 and 40000 ppm CO2 in N2, every time samples were analysed.
Statistical analysis
The experimental design was a completely randomized 2 ´ 4 factorial with five replications. The factors were four soil types with different EC and four soil treatments, that is, unamended soil, urea-amended soil, C2H2 applied soil, and urea + C2H2. Emission of CH4, N2O and CO2 was regressed on elapsed time using a linear regression model, which was forced to pass through the origin, but allowed different slopes (production rates) for each treatment. Significant differences for the production of CH4, N2O and CO2 production between treatments, soil and their interactions were determined using PROC MIXED (SAS Institute Inc. 1989).
Concentrations of NH4+, NO2- and NO3- were subjected to an analysis of variance using PROC GLM (SAS Institute Inc. 1989) to test for significant differences between soils, treatments and their interactions with Tukey’s Studentized Range test. All data presented were the mean of five replicates, that is, n = 5.
RESULTS
Emissions of CO2, CH4 and N2O
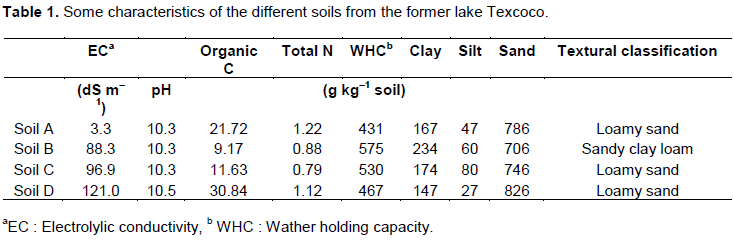
In the unamended soil, the emission of CO2 was largest in soil A and lowest in soil D (Figure 1a). Application of C2H2 to the unamended soil had no significant effect on the CO2 emission rates. The emission of CH4 was similar in all unamended soils and was not affected by the application of C2H2 (Figure 1b). In the unamended soil, the emission of N2O was significantly larger in soil A than in the other soils (P<0.05) (Figure 1c). Application of C2H2 had no significant effect on the N2O emission rate.
Application of urea increased the emission of CO2 in soils A, B and D significantly, but not in soil C (P<0.05) (Figure 1d). Application of C2H2 to the urea-amended soil decreased the emission of CO2 significantly in soils A and B, but not in soils C and D (P<0.05). The emission of CO2 was similar in the C2H2-amended soils applied with or without urea. Application of urea did not affect the emission of CH4 and was similar in the C2H2-amended soils applied with or without urea (Figure 1e). Application of urea increased the emission of N2O significantly in soil A, but not in the other soils (P<0.05) (Figure 1f). Application of C2H2 to the urea-amended soil decreased the emission of N2O in soils A and B, but not in soils C and D.
The emission of CO2 was significantly affected by soil and the interactions between urea ´ C2H2 and soil ´ urea ´ C2H2 (P<0.05) (Table 2). The emission of N2O was significantly affected by the different interactions between urea, soil and C2H2, and the emission of CH4 was affected significantly only by soil (P<0.05).
Dynamics of inorganic N
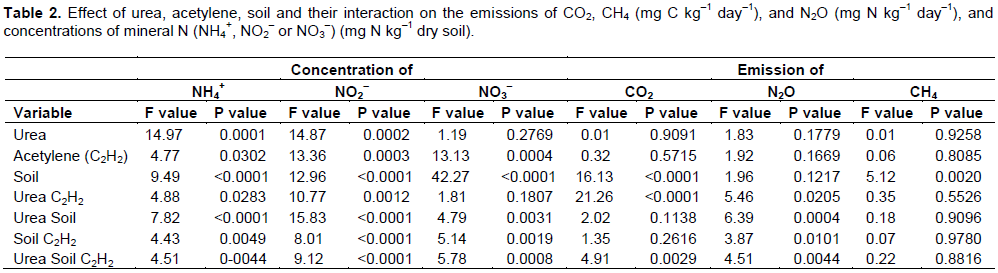
The concentration of NH4+ was similar in the unamended soils and soils applied with C2H2 (Figure 2a). Application of C2H2 to the unamended soil reduced the concentration of NO2- significantly in soils B and C, but not in soils A and D (P<0.05) (Figure 2b). The concentration of NO3- was similar in the unamended soils and soils with applied C2H2 (Figure 2c).
Application of urea increased the concentration of NH4+ significantly in all soils and the increase was most accentuated in soil A (P<0.05) (Figure 2d). Application of C2H2 to the urea-amended soil decreased the amount of NH4+ significantly in soil A, but not in the other soils (P<0.05). Application of urea increased the concentration of NO2- significantly in soil A, but not in the other soils (P<0.05) (Figure 2e). Application of C2H2 to the urea-amended soil decreased the amount of NO2- significantly in soil A, but not in the other soils (P<0.05). Application of urea increased the concentration of NO3- significantly in soil A, but not in the other soils (Figure 2f). Application of C2H2 to the urea-amended soil decreased the amount of NO3- in soil A, but not in the other soils.
The concentrations of NH4+, NO2- and NO3- were affected significantly by urea, C2H2, soil and their interactions, except for the effect of urea and its interaction with C2H2 on the concentration of NO3- (Table 2).
DISCUSSION
Emissions of CO2, CH4 and N2O
The emission of CO2 decreased with increased EC in the soils. It is well known that increased salinity reduces the soil microbial biomass and inhibits microbial activity (Setia et al., 2011a, b). However, it has to be remembered that other characteristics, such as soil organic matter content, clay content and pH, are also known to affect microbial activity and thus emissions of CO2 (Setia et al., 2011a, b).
The application of urea increased the CO2 emission rate significantly in soils A, B and D compared to the unamended soil. It is well known that application of an N fertilizer to an N depleted soil can increase emission of CO2 as microbial activity is stimulated (Wang et al., 2011). The high salt content in the Texcoco soils will inhibit N2 fixation, which will limit the N content of the soil (Barua et al., 2011; Welsh et al., 2007). Additionally, hydrolysis of urea will release CO2 (Snyder et al., 2009). The emission of CO2 will thus increase with 86 mg CO2 after the application of 200 mg N kg-1 soil if all urea was hydrolyzed.
The application of C2H2 had no effect on emission of CO2. Acetylene can be used by certain organisms, e.g. Rhodococcus opacus, Rhodococcus ruber and Gordona species, as C substrate thereby increasing emission of CO2 (Rosner et al., 1997). Soil characteristics are known to affect C2H2 degradation (Brzezinska et al., 2011), although the limited time that the soil microorganisms were exposed to C2H2 (7 days) might have reduced the possibility that they use C2H2 as C source, that is, they were not yet adapted.
Agricultural soils can be a source or a sink for CH4, but they are normally a sink and fluxes are normally small (Wang et al., 2011). Large amounts of CH4 are only emitted from paddy soils or wetlands (Wright et al., 2011). Production of CH4 occurs under anaerobic and oxidation under aerobic conditions. Although, the soils were incubated aerobically, emission of CH4 occurred in all soils. Anaerobic micro-sites exist even in a soil at 40% WHC that will stimulate production of CH4, and oxidation of CH4 did not match production. The high salt content might have inhibited methanotrophic activity.
Application of urea or C2H2 did not affect emissions of CH4. Application of NH4+, released after the hydrolysis of urea, is known to inhibit oxidation of CH4, but not in the Texcoco soils (Stiehl-Braun et al., 2011; Xu et al., 2011). Aronson and Helliker (2010) reported after a meta-analysis that not only the amount of N applied, but also the history of the soil affected the inhibitory effect. They reported that managed soil and soil with a longer duration of fertilizer application showed greater inhibition of CH4 uptake with added N. The Texcoco soil was not fertilized and is N depleted so it can be assumed that N fertilizer would not inhibit CH4 oxidation. Bronson and Mosier (1994) reported a strong inhibitory effect of C2H4 on oxidation of CH4 (76 to 100% inhibition) in two soils. No such inhibitory effect was found in the Texcoco soil, so it can be speculated that little or no CH4 oxidation occurred as stated before.
Application of urea increased emission of N2O in soil A compared to the unamended soil, but not in the other soils. It is well known that application of urea to soil increases emission of N2O (Wang et al., 2011). Emission of N2O from soil is mainly due to nitrification, that is, the oxidation of NH4+ to NO2- and NO2- to NO3- under aerobic conditions and denitrification, that is, the reduction of NO3- to NO2-, N2O and N2 under anaerobic conditions (Wrage et al., 2001). As the soil was incubated under aerobic conditions and the concentration of NH4+ sharply increased after application of urea and its subsequent hydrolysis, emission of N2O was most likely due to oxidation of NH4+. Application of C2H2 (as an inhibitor of the oxidation of NH4+) to soil A sharply reduced the emission of N2O confirming that oxidation of NH4+ was the main source of N2O emission. The emission of N2O in soil A amended with urea plus C2H2, however, was still higher than in the unamended control soil. As such, although the soil was incubated aerobically, it is likely that some anaerobic microsites were formed in soil stimulating denitrification and thus emission of N2O.
Emission of N2O also increased when urea was added to soil B, and C2H2 decreased it. As such, nitrification contributed to the emission of N2O in soil B. No increase in emission of N2O occurred in soils C and D amended with urea. As such, the high salt content inhibited the nitrification process in soils C and D. Application of urea might be used as N fertilizer for a pioneering vegetation to minimise N2O emission in the alkaline saline Texcoco soil. However, it would have to be injected into the soil as the high pH will favour NH3 volatilization.
The soils were incubated at a constant water content in this experiment. In the field, water content will fluctuate continuously thereby changing soil conditions constantly. These constantly changing conditions will put a further strain on the microbial population. The soil micro-organisms will have to adapt strategies to survive a dried out or flooded environment, to altering anaerobic and aerobic conditions, and salt concentrations that decrease in the rainy season but increase in the dry season when evaporation will concentrate the salt ions mostly in the upper soil layer.
Dynamics of mineral N
Concentration of NH4+ increased in all urea-amended soils. Hydrolysis of urea can occur in three ways, biotic (Burton and Prosser, 2001), abiotic in soil with a high pH (Ghandi and Paliwal, 1976) and abiotic through extracellular ureases (Conrad, 1996) generating two NH3 molecules. As such, concentration of NH4+ will increase in urea-amended soil (Burton and Prosser, 2001) as found in this study. The increase in the concentration of NH4+ was lower in soil with EC ≥88.3 dS m-1 than in soil with EC 3.3 dS m-1, so the high EC had an inhibitory effect on the hydrolysis of urea (Wilson et al., 1999).
Application of C2H2 reduced the concentration of NO2- in the unamended or urea-amended soils A, B and C, but not in soil D. Consequently, oxidation of NH4+ occurred in soil with EC ≤96.9 dS m-1. Ammonium oxidizing organisms have been found in extreme environments (Sorokin and Kuenen, 2005) so it would come as no surprise that oxidation of NH4+ occurred in soil with EC≤96.9 dS m-1. Oxidation of NO2- only occurred in the soil with the lowest EC. Although NO2- oxidizing bacteria have been isolated from alkaline environments (Sorokin et al., 1998), it might well be that the extreme high EC inhibited NO2- oxidation as the energetic gain from this process is low (Oren, 2011).
Under the experimental conditions, NO2- oxidation seems to be absent in soils with EC ≥88.3 dS m-1. However, NH4+ oxidation occurred even at 96.9 dS m-1 suggesting that NH4+ oxidation is less sensitive to salinity than NO2- oxidation. From a biological point of view, there are several possible explanations for this phenomenon. First, aerobic ammonium oxidation yields more energy for growth than nitrite oxidation (Bock and Wagner, 2006). Second, aerobic NH4+ oxidation is done not only by a restricted group of Bacteria, but also by Archaea belonging to the phylum Thaumarchaeota (Leininger et al., 2006). These Archaea have a different physiology than NH4+ oxidizing Bacteria. In some environments, these Archaea can even be the major NH4+ oxidizers (Prosser and Nicol, 2008). Furthermore, even when aerobic NH4+ and NO2- oxidizers form tight associations, that is, the NO2- produced by NH4+ oxidizers is consumed by NO2- oxidizers, there seems to be ecophysiological differences between both groups that might be in part the consequence of the evolution of their metabolic life styles. Nitrite oxidizers can be heterotrophic/mixotrophic or strict chemotrophics (Bock and Wagner, 2006), and even between them there are differences since Nitrospira species can be K-strategists with high substrate affinity and low growth rate, while Nitrobacter species might be r-strategists (Schramm et al., 1999). However, more studies need to be done as new nitrite oxidizers groups are emerging and their physiological must still be studied.
In this study, the microbial population was not investigated. It would be interesting to study the microbial population in each of the treatments and investigate which organisms were involved in each of the processes discussed, e.g. nitrifiers, methanogens and methanotrophs. A transcriptomics analysis would surely reveal genes that are relevant in these extreme environments, but absent in more normal soil conditions.
It was found that urea increased emission of CO2 in all soils and emission of N2O in soil with EC ≤88.3 dS m-1, but emission of CH4 was not affected. Hydrolysis of urea occurred in all soils although it was significantly slower in soil with EC ≥88.3 dS m-1. Oxidation of NH4+ occurred in soil with EC≤96.9 dS m-1, but oxidation of NO2- only in soil with EC 3.3 dS m-1.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The research was funded by Cinvestav. C.V.-E. received grant-aided support by ‘Consejo Nacional de Ciencia y Tecnología’ (CONACyT).
REFERENCES
|
Aronson EL, Helliker BR (2010). Methane flux in non-wetland soils in response to nitrogen addition: a meta-analysis. Ecology 91:3242-3251. |
|
|
Barua S, Tripathi S, Chakraborty A, Ghosh S, Chakrabarti K (2011). Studies on non-symbiotic diazotrophic bacterial population of coastal arable saline soils of India. Indian Journal of Microbiology 51:369-376. |
|
|
Bateman EJ, Baggs EM (2005). Contributions of nitrification and denitrification to N2O emission from soils at different water-filled pore space. Biology and Fertility of Soils 41:379-388. |
|
|
Bock E, Wagner M (2006). Oxidation of Inorganic Nitrogen Compounds as an Energy Source. In The Prokaryotes, 3rd ed (eds Dworkin M, Falkow S, Rosenberg E, Schleifer K-H & Stackebrandt E), Springer New York pp. 457-495. |
|
|
Bronson KF, Mosier AR (1994). Suppression of methane oxidation in aerobic soil by nitrogen fertilisers, nitrification inhibitors, and urease inhibitors. Biology and Fertility of Soils 17:263-268. |
|
|
Brzezinska M, Rafalski P, Wlodarczyk T, Szarlip P, Brzezinski K (2011). How much oxygen is needed for acetylene to be consumed in soil? Journal of Soils and Sediments 11:1142-1154. |
|
|
Burton SAQ, Prosser JL (2001). Autotrophic ammonia oxidation at low pH through urea hydrolysis. Applied and Environmental Microbiology 67:2952-2957. |
|
|
Conde E, Cardenas M, Ponce-Mendoza A, Luna-Guido ML, Cruz-Mondragón C, Dendooven L (2005). The impacts of inorganic nitrogen application on mineralization of 14C-labelled maize and glucose, and on priming effect in saline alkaline soil. Soil Biology and Biochemistry 37:681-691. |
|
|
Conrad R (1996). Soil microorganisms as controllers of atmospheric trace gases (H2, CO, CH4, OCS, N2O and NO. Microbiological Reviews 60:609-640. |
|
|
Dendooven L, Alcántara-Hernández RJ, Valenzuela-Encinas C, Luna-Guido M, Pérez F, Marsch R (2010). Dynamics of carbon and nitrogen in an 'extreme' alkaline saline soil: a review. Soil Biology and Biochemistry 42:865-877. |
|
|
Ghandi AP, Paliwal KW (1976). Mineralization and gaseous losses of nitrogen from urea and ammonium sulfate in salt-affected soil. Plant and Soil 45:247-255. |
|
|
Gutiérrez-Castorena MD, Stoops G, Solorio CAO, Avila GL (2005). Amorphous silica materials in soils and sediments of the Ex-lago de Texcoco, Mexico: An explanation for its subsidence. Catena 60:205-226. |
|
|
Leininger S, Urich T, Schloter M, Schwark L, Qi J, Nicol GW, Prosser JI, Schuster SC, Schleper C (2006). Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature 442:806-809. |
|
|
Luna-Guido ML, Beltrán-Hernández RI, Solis-Ceballos NA, Hernández-Chavez N, Mercado-García F, Olalde-Portugal V, Catt JA, Dendooven L (2000). Chemical and biological characteristics of alkaline saline soils from the former Lake Texcoco as affected by artificial drainage. Biology and Fertility of Soils 32:102-108. |
|
|
O'Hara SL, Metcalfe SE (1997). The Climate of Mexico since the Aztec Period. Quatern Int. 43:25-31. |
|
|
Oren A (2011). Thermodynamic limits to microbial life at high salt concentrations. Environmental Microbiology 13:1908-1923. |
|
|
Prosser JI, Nicol GW (2008). Relative contributions of archaea and bacteria to aerobic ammonia oxidation in the environment. Environmental Microbiology 10:2931-2941. |
|
|
Rosner BM, Rainey FA, Kroppenstedt RM, Schink B (1997). Acetylene degradation by new isolates of aerobic bacteria and comparison of acetylene hydratase enzymes FEMS Microbiology Letters 148:175-180. |
|
|
Ruíz-Valdiviezo VM, Luna-Guido M, Galzy A, Gutiérrez-Miceli FA, Dendooven L (2010). Greenhouse gas emissions and C and N mineralization in soils of Chiapas (México) amended with leaves of pi-ón (Jatropha curcas L.). Applied Soil Ecology 46:17-25. |
|
|
SAS Institute (1989). Statistic Guide for Personal Computers. Version 6.04, Edn SAS Institute, Cary. |
|
|
Schramm A, de Beer D, van den Heuvel JC, Ottengraf S, Amann R (1999). Microscale distribution of populations and activities of Nitrosospira and Nitrospira spp. along a macroscale gradient in a nitrifying bioreactor: quantification by in situ hybridization and the use of microsensors. Applied and Environmental Microbiology 65:3690-3696. |
|
|
Setia R, Marschner P, Baldock J, Chittleborough D, Smith P, Smith J (2011a). Salinity effects on carbon mineralization in soils of varying texture. Soil Biology and Biochemistry 43:1908-1916. |
|
|
Setia R, Marschner P, Baldock J, Chittleborough D, Verma V (2011b). Relationships between carbon dioxide emission and soil properties in salt-affected landscapes. Soil Biology and Biochemistry 43:667-674. |
|
|
Snyder CS, Bruulsema TW, Jensen TL, Fixen PE (2009). Review of greenhouse gas emissions from crop production systems and fertilizer management effects. Agriculture, Ecosystems and Environment 133:247-266. |
|
|
Sorokin DY, Kuenen JG (2005). Chemolithotrophic haloalkaliphiles from soda lakes. FEMS Microb. Ecol. 52:287-295. |
|
|
Sorokin DY, Muyzer G, Brinkhoff T, Kuenen JG, Jetten M (1998). Isolation and characterization of a novel facultatively alkaliphilic Nitrobacter species -Nb. alkalicus. Archives of Microbiology 170:345-352. |
|
|
Stiehl-Braun PA, Hartmann AA, Kandeler E, Buchmann N, Niklaus PA (2011). Interactive effects of drought and N fertilization on the spatial distribution of methane assimilation in grassland soils. Global Change Biology 17:2629-2639. |
|
|
Wang WJ, Dalal RC, Reeves SH, Butterbach-Bahl K, Kiese R (2011). Greenhouse gas fluxes from an Australian subtropical cropland under long-term contrasting management regimes. Global Change Biology 17:3089-3101. |
|
|
Welsh A, Burke DJ, Hahnn D (2007). Analysis of nitrogen-fixing members of the epsilon subclass of proteobacteria in salt marsh sediments. Applied and Environmental Microbiology 73:7747-7752. |
|
|
Wilson DJ, van der Wal R, Chang ER, Jensen A, Jefferies RL (1999). Urea hydrolysis and nitrification in arctic salt-marsh soils: Possible constraints on the growth of forage plants. Ecoscience 6:72-78. |
|
|
Wrage N, Velthof GL, Van Beusichem ML, Oenema O (2001). Role of nitrifier denitrification in the production of nitrous oxide. Soil Biology and Biochemistry 33:1723-1732. |
|
|
Wright EL, Black CR, Cheesman AW, Drage T, Large D, Turner BL, Sjogersten S (2011). Contribution of subsurface peat to CO2 and CH4 fluxes in a neotropical peatland. Global Change Biology 17:2867-288. |
|
|
Xu XK, Han L, Luo XB, Han SJ (2011). Synergistic effects of nitrogen amendments and ethylene on atmospheric methane uptake under a temperate old-growth forest. Advances in Atmospheric Sciences 28:843-854. |
|
|
Zhu TB, Zhang JB, Cai ZC (2011). The contribution of nitrogen transformation processes to total N2O emissions from soils used for intensive vegetable cultivation. Plant Soil 343:313-327. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


