ABSTRACT
Moringa oleifera is a fruit species of economic interest to West African smallholder growers. To generate beneï¬ts to poor rural communities, improved knowledge is needed on plantation management and selection of the most reliable seed sources. The aim of the study was to investigate variation in seed traits and germination rates among 12 provenances of M. oleifera from West Africa in the women gardening center named “Amicale des Forestières du Burkina Faso (AMIFOB)” located at Ouagadougou, Burkina Faso (12°7’32’’N, 01°40’24’’W). The authors conducted an analysis of variance, a principal component analysis on seed traits variables (length, thickness and weight) and germination rates at 5 and 12 days after seed sowing, and finally ascending hierarchical classification based on similarity indices. The results showed significant variations (P<0.05) among provenances in seed traits: Ouahigouya provenance in the Sahelian area of Burkina Faso had the largest and heaviest seeds. Germination rate was significantly different after 5 days for Ouahigouya, Ouagadougou, Koudougou, Dano and Tamale provenances (P<0.05). Five days after seed sowing, the Sahelian provenance (Ouahigouya) recorded the greatest and fastest germination rate of 63%. Correlation analyses revealed no significant links between germination rate in 5 and 12 days after sowing and seed sizes. Seed traits and germination rates did not show clear cut distinct groups between Sahelian, Sudanian and sub Equatorial provenances. This research output provides an evidence of the genetic variability among M. oleifera provenances and hence the potential for future tree improvement programme.
Key words: Seed variability, agroclimatic zone, Burkina Faso.
Moringa oleifera Lam. or ‘drumstick tree’ is a member of the Moringaceae family, which grows throughout the tropics and is native to the sub-Himalayan tracts of north-western India, Pakistan, Bangladesh and Afghanistan (Makkar and Becker, 1997). The tree has a height of 7 to 12 m up to the crown. The leaves are twice or thrice pinnate and spirally arranged. The flowers are bisexual, white or cream colored with yellow stamens, and the fruit is a three sided or nearly cylindrical capsule. The seed is round, dark chestnut-colored, and with 3 white wings that facilitate spreading by the wind in natural conditions. Moringa can be propagated either by using seeds or cuttings (Morton, 1991). Fruits, seeds, leaves and flowers are consumed by humans as nutritious vegetables. M. oleifera leaves are known to be an excellent source of vitamins, minerals and protein and are used to fight against malnutrition and its associated diseases including blindness (Asante et al., 2014). In West Africa, leaves are commonly used to make sauces. It is grown in all types of soil, from acid to alkaline (Galdima et al., 2013) and can tolerate up to 6 months of dry season reasonably well. The tree grows at different altitudes from 0 to 1800 m.a.s.l. and rainfall between 500 and 1500 mm per year.
M. oleifera is planted and smallholder growers grow subsistence and cash crops in their rainfed, complex and resource deprived fields often combining the cultivation of the crops with scattered multipurpose trees and realizing a wide range of benefits (Mudyiwa et al., 2013). It is one of such multipurpose tree of global interest and is grown in combination with agricultural and horticultural crops by smallholder growers and this give growers a wide range of benefits (Mudyiwa et al., 2013). It is a suitable tree for traditional agroforestry in the home because of its versatility (Nduwayezu et al., 2007).
In Burkina Faso, the seeds sources for planting are coming from various agro ecological zones. For example, more than 80% of women growers used non certified seeds as plantation materials to produce M. oleifera leaves because they do not access the high cost of certified one. The demand of plantation material is most likely growing in the future due to increasing demand of the potential market. Better plantation and nursery management would generate beneï¬ts to poor rural communities, since villages and hamlets hold their own local nursery. However, before plantations can be recommended on a large-scale, improved knowledge is needed on plantation management and selection of the most reliable seed sources. There are noticeable differences among M. oleifera agroecological ecotypes concerning growth performances (Edward et al., 2014) which have considerable impact on the economic return. Differences among geographic sources in tree species are often substantial and economic improvement can be made by an appropriate provenance selection (Wright et al., 1976).
Altitudinal variations in seed and seedling characters of tree species have been reported by various studies (Gera et al., 2000). However, for the utilization of observed variation in species, it is a prerequisite to know the extent of variation and also whether it is due to genetic or environmental factors. Hence, information on variation in the desirable parameters and their correlation is vital for any breeding programme (Fenner and Thompson, 2005).
Little is known about the role of M. oleifera seed size. Seed size has been regarded as an important plant property (Fenner, 1991). Considering the presence in various ranges of climate and soil conditions, and different agro ecological zones with different ecotypes may show contrasting performances (Aronson et al., 1993).
In the current study, the authors investigated the seed size among 12 provenances of M. oleifera from four agroecological zones of West Africa. They hypothesized that the seed size and germination rates correlated with geoclimatic data of seed collection sites.
Study site
The study was conducted at the women gardening center named “Amicale des Forestières du Burkina Faso (AMIFOB)” located at Ouagadougou, Burkina Faso (12°7’32’’N, 01°40’24’’W). The rainfall is uni-modal with a mean annual rainfall of the last 15 years data from the nearest meteorological station in Ouagadougou, of 800 mm.year-1, and the mean temperature of 28.5°C month-1 (Figure 1). Soils are sandy clay to clay-sandy Ferruginous leached with very low nutrient content according to French soil classification (Pallo et al., 2009). The common natural vegetation found at Ouagadougou is described as semi-deciduous open woodland. Main genera include, Eucalyptus, Azadirachta, Mangifera, Vitellaria, Lannea, Piliostigma, Acacia., Ziziphus, Tamarindus and Combretum.
Seed sources, experimental design and establishment
The experiment included twelve M. oleifera provenances: 1 provenance from Segou (Mali), 1 from Niangon-Lokoua, Abidjan (Ivory Coast), 1 from Tamale (Ghana) and 9 from Burkina Faso (Ouahigouya, Dano, Gaoua, Ouagadougou, Fada N’Gourma, Dédougou, Bobo-Dioulasso, Koudougou, Centre National de Semences Forestières-CNSF) (Figure 2).
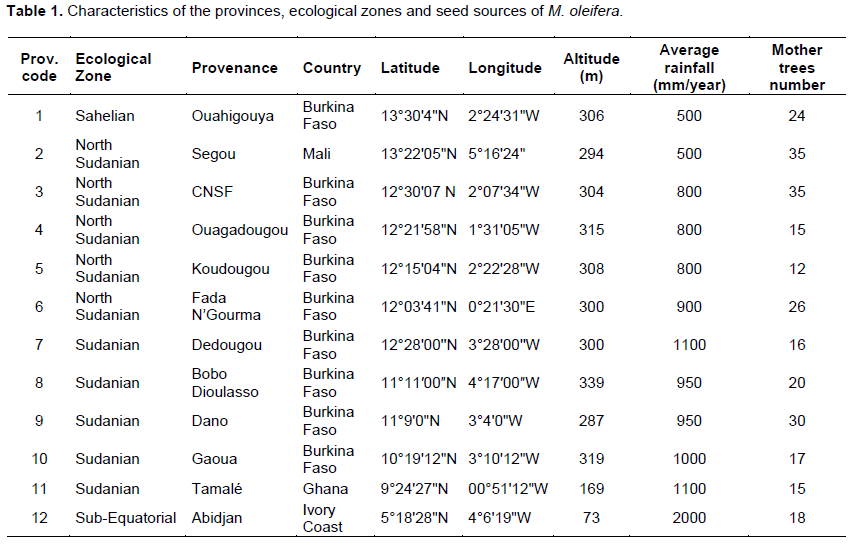
In this paper, provenances were referred to four climate areas according to their agro-ecological characteristics (Sahelian, sub Equatorial, Sudanian and north Sudanian) (Table 1). Seeds were collected in 2014 in plantation farmland from at least 12 mother trees per provenance.
Trial was planted in a randomized complete block design (RCBD) with three replications. Each plot represented a provenance planted at 5 x 6 rows in a contiguous arrangement of 20 x 20 cm (Figure 3). Plot measured 10 x 1 m and contained 30 trees and the distances between blocks were 2 m. Seed samples were pretreated with water for 24 h and sown in June 1st 2014 in a prepared and cleaned soil using hand hoes. Weeding was done twice during the rainy season. Watering was done once a day in the morning. The seedlings were grown without fertilizers and chemical control.

Data collection and analysis
To investigate the variability in seed parameters (length, thickness and weight), each provenance was represented by 240 randomly selected seeds, assessed in four replications of 60 seeds each. Each seed weight was determined by weighing three random samples of 60 seeds each. By the end of 2 weeks, the experiment had been measured twice at effective ages of 5 and 12 days since seed sowing, providing data for the assessment of germination rates of the twelve provenances. All statistical analysis were carried out using ANOVA performed with the MIXED procedure of JMP® Pro 11.1.1 (SAS Institute, Cary, NC, USA). Normality and homoscedasticity were graphically verified on residual plots of the linear models (Quinn and Keough, 2002). When effects were significant, the Tukey-Kramer test was used for multiple mean comparisons. Statistical significance was fixed at 0.05, and only significant results are presented in graphs. A factorial analysis was conducted to determine the relationship of seeds, germination and their provenances of origin.
Seed size traits
Analysis of variance showed highly significant differences among M. oleifera provenances within ecological zones for seed length, thickness and seed weight (P < 0.0001) (Figure 4). Estimates of variance components indicated that the inter provenances effects varied from 42 to 45% of the total variation for all seed characters (Table 2). The provenance heritability for all seed characters was relatively high (0.94).
Maximum mean values for seed length (15.8 mm) were found in Fada N’Gourma, for seed thickness (11 mm) in 5 provenances (Ouahigouya, Ségou, Koudougou, Gaoua and Abidjan) and for seed weight (1.8 g) in Ouahigouya
and Gaoua provenances (Table 3). Ranking of provenances based on mean seed length, thickness and weight indicated consistently lower values for Koudougou and Bobo Dioulasso (Table 3). Ouahigouya (Sahelian) and Gaoua (south Sudanian) were characterized by heaviest and largest seeds, while Koudougou (north Sudanian) and Bobo Dioulasso (south Sudanian) provenances were the shortest, tiniest and lightest seeds. The provenances were classified in descending order of magnitude by high, intermediate and low groups of seed characters as follows:Seed length: Fada N’Gourma > Ouahigouya, CNSF, Abidjan, Gaoua, Ségou and Tamale > Ouagadougou, Dano, Koudougou and Bobo Dioulasso Seed thickness: Ouahigouya, Gaoua, Ségou, Ouagadougou, Tamale > Fada N’Gourma, Dano, CNSF and Tamale > Koudougou and Bobo Dioulasso Seed weight: Ouahigouya, Gaoua > Ségou, Ouagadougou, Tamale, Abidjan, CNSF, Dano > Fada N’Gourma, Koudougou and Bobo Dioulasso.
Seed germination
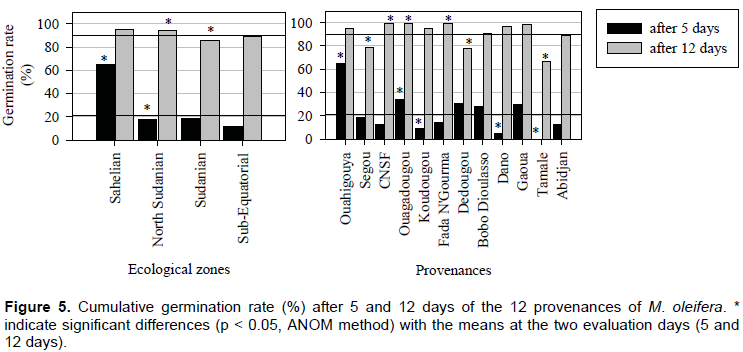
Seeds started germinating 5 days after sowing and after day 12 no seeds germinated. The overall means of germination rate of M. oleifera seeds under experimental conditions were 21 and 90% after 5 and 12 days after sowing, respectively (Figure 5). The cumulative germination rate was significantly different after 5 days for Ouahigouya, Ouagadougou, Koudougou, Dano and Tamale (P < 0.05) (Figure 5). Five days after seed sowing, Ouahigouya provenance recorded the greatest and fastest germination rate of 63%.
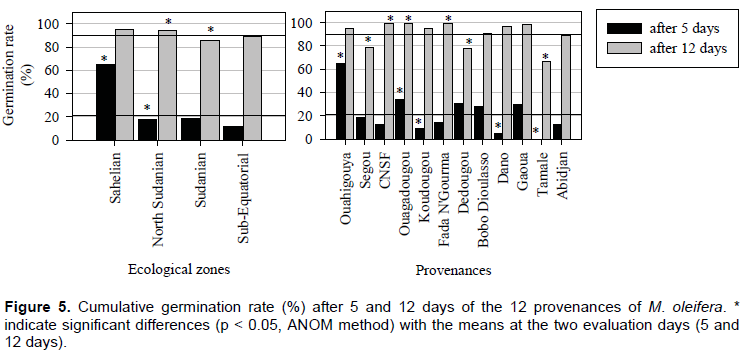
The response of seeds showed great variability depending on provenance and ecological zone (Figure 5). Significant effects were found between ecological zones (P < 0.05). After 5 days, germination rate of the provenance in the Sahelian zone (Ouahigouya) was 3 times higher (more than 60%). Within the ecological zone, no significant effects were observed at both dates 5 and 12 days after sowing with Abidjan (sub Equatorial), Gaoua and Bobo Dioulasso (south sudanian) provenances (Figure 3). The highest final germination percentages of 100% were observed with provenances from the north Sudanian zone of Burkina Faso (CNSF, Ouagadougou and Fada N’Gourma). The final lowest germination rates were observed in Tamale (68%), Dedougou (78%) and Segou (80%) provenances (Figure 5).
Correlations of seed characteristics with germination rates
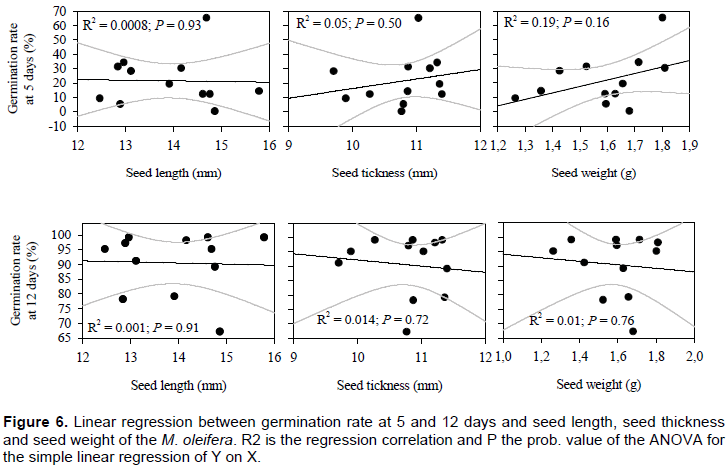
Correlation analyses revealed no significant links between germination rate in 5 and 12 days after sowing and seed length (r2 = 0.00; P = 0.93 and r2 = 0.00; P = 0.91 respectively in 5 and 12 days after seed sowing), seed thickness (r2 = 0.05; P = 0.50 and r2 = 0.01; P = 0.72, respectively in 5 and 12 days after seed sowing) and seed weight (r2 = 0.19; P = 0.16 and r2 = 0.01; P = 0.76, respectively in 5 and 12 days after seed sowing) (Figure 6).
Multivariate analysis of seed characters, germination and geoclimatic data of provenance origin
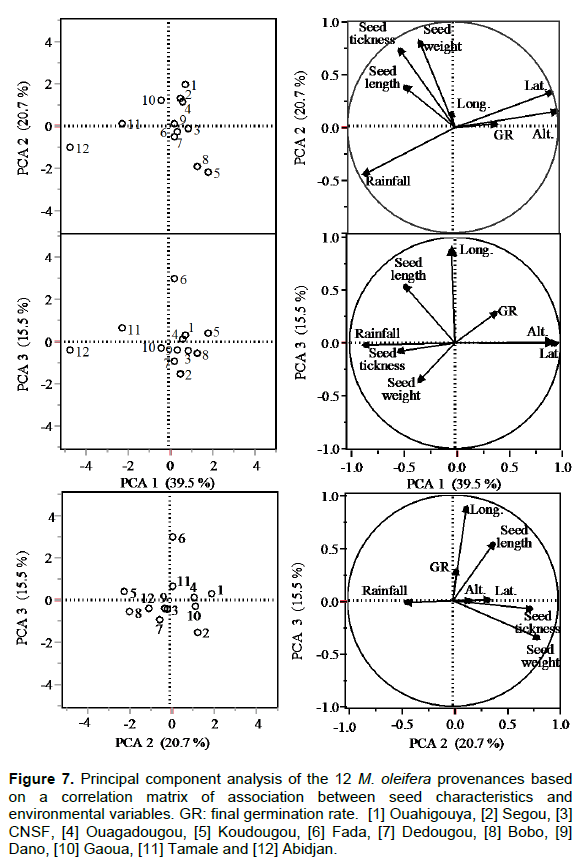
The principal component analysis (PCA) performed on the characteristics of seeds and twelve-day-old seedlings showed that the first three principal axes explained 77% of the observed variability (Axis 1: 40%, axis 2: 21% axis 3: 16%) (Table 3). The axis 1 variables were influenced by the average rain and latitude and altitude of the provenances origin and constituted the axis of geoclimatic characters of the provenance origin. The axis 2 explained the contribution to total variation of variables seed thickness and weight. This axis may be defined as the center of the seed characteristics. The axis 3 was mainly explained by the seed length, germination rate and longitude and was defined as the axis of the seed length (Figure 7).

By overlaying the correlation circles of variables and map of overall seed provenances with the main axes, it appears that provenances 11 (Tamale), and 12 (Abidjan) are highly correlated with average rainfall of the axis 1 and located at the negative side of the axis 1 (Figure 7). Axe 3, the provenance 12 and 11 and the variables seed thickness and weight were located on the positive side of the axis 3 while the provenance 6 and the variable seed length were on the opposite side .A hierarchical cluster analysis dendrogram showned no clear identified provenance groups (Figure 8).
This study was conducted in Ouagadougou on 12 provenances of M. oleifera from West African, and tested the hypothesis that seed size and germination rates were correlated with geoclimatic data of seed collection sites. The results revealed clear differences among the 12 M. oleifera provenances in terms of seed weight, length and thickness. Seeds collected from Ouahigouya in the sahel zone were heavy and large with faster germination rate in the natural environmental conditions. In contrast, seeds collected in provenances located in more humid zones (north and south sudanian, sub equatorial) were small, less heavy and thin.
Seed size, is regarded as key factor of reproductive strategy because of its important role in establishment of the juvenile phase of tree life cycle. For example, large seed size has been documented to prolong dormancy during unfavorable light conditions, development of large amounts of photosynthetic tissue, allowing quick seedling growth and dispersal modes (Mamo et al., 2006). According to Roach et al. (1987), in very wet areas where rainfalls vary between 1000 and 1500 mm · year-1, the seeds would be generally large. In this study, our results in contrary showed that the provenances seeds from wetter areas were smaller. The observed seed sizes differences are rather due to the effects of the genetics and would therefore not be environmentally based (Roach et al., 1987). Much of the total variation in seed traits are due to genetic influence as evidence from high heritability estimates (>0.90%). These genetic variations in seed traits have been reported for several tree species (Bonito et al., 2011; Khan et al., 2012; Ky-Dembele et al, 2014). Our findings in M. oleifera are in conformity with these research findings. Similar genetic variation in seed sides has been reported for several tree species (Diallo et al., 2010; Ky-Dembele et al., 2014).
The present study revealed the existence of significant differences of germination rates between provenances 5 days after seeds sowing. Ouahigouya provenance from the dry agroclimatic area recorded the greatest and fastest germination rate of 63%. This variation among provenances at early stage of germination indicated that greater and heavy seeds of Ouahigouya provenance results in faster seedlings growth. Rapid germination and subsequent seedling growth are, therefore, key phenotypes of vigorous seeds that are known to differ with genetic background (Bettey et al., 2000). Thus, a vigorous seed must possess key trait of rapid germination to establish seedlings across a wide range of environments (Khan et al., 2012).
In the present study, the superior performance of Ouahigouya in seed size and germination rate suggests that conservation and management of this provenance as seed source could improve M. oleifera in the north Sudanian and Sahelian areas of Burkina Faso. In most plant species, seeds vary in their degree of germinability between and within populations and between and within individuals (Nyoka et al., 2015). Some of this variation can be of genetic origin, but much of it is known to be environmental, that is caused by the local conditions under which the seed matured (Mamo et al., 2006).
The study revealed no significant correlations between seed characters and germination capacity. The finding suggests that seed weight, length and thickness have little importance in predicting the germinability of Moringa seeds. This was already demonstrated by Fenner and Thompson (2005) and Kazmi et al. (2012). Thus, seed sides are beneficial for the establishment of seedlings, but there appear to be no consistent correlation between seed and germination characteristics.
The hierarchical cluster analysis dendrogram revealed no clear difference between provenances. Thus, no groups of provenances were markedly discernable for seed characters, germination and geoclimatic data of provenance origin. This method of grouping of seed provenances indicated that distribution into different groups did not follow any pattern with regard to agroclimatic seed sources.
This study is the first in West African to characterize Moringa seed sizes and germination rates. The results indicate that variability exist among provenances in seed sizes of M. oleifera from different agroclimatic zones of West Africa. Based on observed seed sizes and germination rates, the most prominent provenance is Ouahigouya. Seeds collected from this provenance are heavy and large with faster germination rate; thus this provenance might be considered for an eventual M. oleifera improvement program to enhance productivity in plantations in north Sudanian and Sahelian areas of Burkina Faso.
The authors have not declared any conflict of interests.
REFERENCES
|
Aronson J, Floret C, Le Floc'h E, Ovalle C, Pontanier R (1993). Restoration and Rehabilitation of Degraded Ecosystems in Arid and Semi-Arid Lands. I. A View from the South. Restor. Ecol. 1(1):8-17.
Crossref
|
|
|
|
Asante WJ, Nasare IL, Damian TD, Kwame OB, Kwami BK (2014). Nutrient composition of Moringa oleifera leaves from two agro ecological zones in Ghana. Afr. J. Plant Sci. 8(1):65-71.
Crossref
|
|
|
|
|
Bettey M, Finch-Savage WE, King GJ, Lynn JR (2000). Quantitative genetic analysis of seed vigour and pre-emergence seedling growth traits in Brassica oleracea. New Phytol. 148(2):277-286.
Crossref
|
|
|
|
|
Bonito A, Varone L, Gratani L (2011). Relationship between acorn size and seedling morphological and physiological traits of Quercus ilex L. from different climates. Photosynthetica 49(1):75-86.
Crossref
|
|
|
|
|
Diallo BO, Joly MH, Mc Key D, Hossaert-Mckey M (2010). Variation des caractères biométriques des graines et des plantules de neuf provenances de Tamarindus indica L. (Caesalpinioideae). Fruits 65(03):153-167.
Crossref
|
|
|
|
|
Edward E, Chamshama SAO, Ngaga YN, Mndolwa MA (2014). Survival, growth and biomass production of Moringa oleifera provenances at Gairo inland plateau and Ruvu Coastal Region in Tanzania. Afr. J. Plant Sci. 8(1):54-64.
Crossref
|
|
|
|
|
Fenner M (1991). The effects of the parent environment on seed germinability. Seed Sci. Res. 1(02):75-84.
Crossref
|
|
|
|
|
Fenner M, Thompson K (2005). The Ecology of Seeds. Cambridge University Press.
Crossref
|
|
|
|
|
Galdima M, Bakoumé C, Ogéné NA, Boukong A (2013). Impact of Previous Vegetation Cover on Mycorrhizal Colonization and Performance of Moringa oleifera in Rainforest Regions of Cameroon. Agric. Sci. 1(2):32-42.
|
|
|
|
|
Gera MN, Gera B, Gupta BN (2000). Preliminary observations on genetic variability and character association in Dalbergia sissoo Roxb. Indian For. 134:608-615.
|
|
|
|
|
Kazmi RH, Khan N, Willems LAJ, Van Heusden AW, Ligterink W, Hilhorst HWM (2012). Complex genetics controls natural variation among seed quality phenotypes in a recombinant inbred population of an interspecific cross between Solanum lycopersicum × Solanum pimpinellifolium. Plant Cell Environ. 35(5):929-951.
Crossref
|
|
|
|
|
Khan N, Kazmi RH, Willems LAJ, van Heusden AW, Ligterink W, Hilhorst HWM (2012). Exploring the Natural Variation for Seedling Traits and Their Link with Seed Dimensions in Tomato. PLoS ONE 7(8):e43991.
Crossref
|
|
|
|
|
Ky-Dembele, C, Tigabu M, Bayala J, Odén PC (2014). Inter- and intra-provenances variations in seed size and seedling characteristics of Khaya senegalensis A. Juss in Burkina Faso. Agrofor. Syst. 88(2):311-320.
Crossref
|
|
|
|
|
Makkar HPS. Becker K (1997). Nutrients and antiquality factors in different morphological parts of the Moringa oleifera tree. The J. Agric. Sci. 128(03):311-322.
Crossref
|
|
|
|
|
Mamo N, Mihretu M, Fekadu M, Tigabu M, Teketay D (2006). Variation in seed and germination characteristics among Juniperus procera populations in Ethiopia. For. Ecol. Manage. 225(1-3):320-327.
Crossref
|
|
|
|
|
Morton J (1991). The horseradish tree, Moringa pterygosperma (Moringaceae) - A boon to Arid Lands? Econ. Bot. 45(3):318-333.
Crossref
|
|
|
|
|
Mudyiwa SM, Gadzirayi CT, Mupangwa JF, Gotosa J, Nyamugure T (2013). Constraints and opportunities for cultivation of Moringa oleifera in the Zimbabwean smallholder growers. Int. J. Agric. Res. Innov. Tech. 3(1):12-19.
Crossref
|
|
|
|
|
Nduwayezu JB, Sao AM, Ngaga YN, Khonga EB, Chabo RG (2007). Comparisons in seed kernel sizes and early growth performance of different Moringa oleifera provenances in southeast of Botswana. Discov. Innov. 19(1-2):52-58.
Crossref
|
|
|
|
|
Nyoka BI, Chanyenga T, Mng'omba SA, Akinnifesi FK, Sagona W (2015). Variation in growth and fruit yield of populations of Sclerocarya birrea (A. Rich.) Hochst. Agrofor. Syst. 89(3):397-407.
Crossref
|
|
|
|
|
Pallo FJP, Zombré SN, Nabsanna P, Sedogo MP (2009). Statut de la matière organique des sols de la zone nord soudanienne au Burkina Faso. Biotechnol. Agron. Société Environ. 13(1):139-142.
|
|
|
|
|
Quinn GP, Keough MJ (2002). Experimental design and data analysis for biologists. United States of America: Cambridge University Press, New York P 537.
Crossref
|
|
|
|
|
Roach DA, Wulff RD (1987). Maternal Effects in Plants. Annu. Rev. Ecol. System. 18(1):209-235.
Crossref
|
|
|
|
|
Wright JW, Kung FH, Read RA, Sremnorr RJ, Anders JW (1976). Nine year performance of Pinus flexilis and P. strobiformus progenies in Michigan and Nebraska. Silvae Genet. 20:141-144.
|
|