Full Length Research Paper

ABSTRACT
Parasitic weed Striga gesnerioides (Willd.) is one of the major constraints of cowpea production. Host-plant resistance seems to be efficient and economical in controlling the pest. The objectives of this study were to evaluate recombinant inbred lines developed between IT97K 499-35 (Striga resistant parent,) and Sanzi (susceptible parent), by Single Seed Descent (SSD), for Striga resistance in Northern Ghana. The study also evaluated the promising Striga gesnerioides resistant lines and susceptible checks for yield loss due to Striga infestation. The studies involved a field and pot screening under artificial inoculation. Twenty-seven (27) recombinant inbred lines (RILs) out of the 251 RILs screened were resistant to Striga gesnerioides. The percentage reduction in the grain yield and dry biomass were lower in the resistant RILs (0.55 to 3.08% and 1.11 to 7.7%, respectively) than the susceptible ones (28.45 to 58.88% and 47.29 to 61.71%, respectively). The negative effect of Striga infestation on cowpea grain yield and dry biomass can then be reduced when resistant genotypes are used.
Key words: Cowpea, Striga gesnerioides, recombinant inbred lines, yield loss.
INTRODUCTION
Cowpea Vigna unguiculata (L.) Walp.) is an important crop in the semi-arid tropics including parts of Asia, Africa, Southern Europe, Southern United States, Central and South America (Singh, 2005; Timko et al., 2007a). It is an affordable source of quality protein for rural and urban dwellers in Africa (Ajeigbe et al., 2012; Dube and Fanadzo, 2013). The dry grain protein concentration oscillates from 21 to 33% (Abudulai et al., 2016). It is well adapted to hectic environments where several crops fail to grow well (Bisikwa et al., 2014; Ddamulira et al., 2015). According to FAO, cowpea was cultivated on about 12.08 million hectare in Africa in 2016 with a total production of 5.83 million hectare in West Africa, predominantly in Nigeria, Niger, Burkina Faso, Mali and Senegal (FAOSTAT, 2018). Currently, cowpea yields are estimated around 300 to 500 kg ha-1 on farmer’s field in Sub Saharan Africa (SSA) while its yield potential isup to 3000 kg ha-1 in optimum growing conditions (Tanzubil et al., 2008).
Cowpea production is mostly affected by major constraints. Parasitic plants are a major constraint to today’s agriculture with most crop species being potential hosts (Westwood et al., 2010). Striga gesnerioides, is a key threat to cowpea production throughout West and Central Africa (Omoigui et al., 2017). It is one of the greatest devastating parasitic weed in most parts of the world. It is an obligate root parasitic flowering weed that belongs to the Orobanchaceae family (Parker, 2012). Out of about 30 Striga species which have been identified, Striga gesnerioides is the only Striga species that is virulent to dicots (Mohamed and Musselman, 2008). Striga gesnerioides is a major limitation to cowpea production in Africa (Timko et al., 2007b), causing considerable yield losses (Aggarwal and Ouédraogo, 1989). The extent of the damage in cowpea due to S. gesnerioides could be up to 70% depending on the extent of damage and level of infestation (Alonge et al., 2004). On susceptible cultivars, yield losses can reach up to 100% when S. gesnerioides population is over 10 emerged shoots per plant (Kamara et al., 2008). Omoigui et al. (2009) reported that yield losses caused by Striga in dry savannah of SSA are estimated in millions of tons annually and the prevalence of Striga infested soils is steadily increasing. Methods including improved cultural practices and the use of chemicals to control S. gesnerioides are available but most of them are ineffective whilst others are not affordable for small-scale farmers of Sub Saharan Africa (Singh et al., 1997; Timko et al., 2007b). The long viability of the seeds and the subterranean nature of the initial stages of parasitism make the control of the parasite by conventional approaches challenging (Berhane, 2016). In general, S. gesnerioides control is difficult to achieve due to the close association with its host (Lane et al., 1997). The identification of sources of S. gesnerioides resistance and their incorporation into breeding schemes would be a useful approach to combat the damage caused by the parasite in cowpea fields (Omoigui et al., 2017).
The objective of this study was to evaluate the field performance of 251 recombinant inbred lines (RILs) under S. gesnerioides infestation in Northern Ghana and to assess yield loss due to Striga infestation.
MATERIALS AND METHODS
The experiments were conducted from July 2015 to April 2016 at the Manga Station of Council for Scientific and Industrial Research-Savannah Agricultural Research Institute (CSIR-SARI). Manga near Bawku in the Upper East Region is geographically located within latitude 11.02° and longitude 0.27°, with an altitude of 224 m above sea level. The area is situated in the Sudan Savanna agro-ecological zone of Ghana. The mean rainfall of the area during the period of the experiment was approximately 44.33 mm. The average annual temperature was about 29.44°C, the highest being observed from March to April 2016. The relative humidity (RH) of the location fluctuated significantly, dropping in the dry season and rising during the rainy season with an average humidity of 55.4%. The study was conducted in two stages. The first stage was carried out in the field and the second stage in pots experiment.
Planting materials
Two hundred and fifty one recombinants inbred lines (RILs) at F8 generation (F8) (Table 1) derived from a cross between two cowpea lines, ‘Sanzi’ (susceptible to Striga) and ‘IT97K-499-35’ (resistant to Striga) (Omoigui et al., 2009), were used in the study.

Field experiment
The field study was carried out under rain fed conditions (between July and September) and under irrigation during the dry season. In a preliminary screening, each of the RILs and the parents (Sanzi and IT97K-499-35) were planted in a 2-meter single row plot without any replication on a field known to be a Striga hot spot. During this preliminary screening, data collected included days to 50% flowering, presence or absence of Striga plants, number of Striga per plants, total number of Striga per plot and Striga height. The presence or absence of Striga was recorded by visual observation on the different plots from thirty five (35) days after planting (DAP) up to maturity.
Pot experiment
Pot experiment was carried out to confirm the resistance or otherwise of the sixty-nine (69) the RILs that were identified as Striga resistant in the field experiment (Table 2). The pots were filled with top soil and then artificially infested with 5 g of Striga seeds. The top most, (1/3) portion of the soil per pot was mixed with the 5 g of Striga seeds. The infested pots were watered and allowed to condition before planting of the cowpea seeds. The pots were arranged in a randomized complete block designs with three replications. Thirty five days after planting (DAP), the pots were monitored on daily basis to check for Striga emergence. At maturity, the early pods were harvested on single plant basis to get some seeds from each plant. This was followed by gently washing the soil off the roots of the plants to confirm or otherwise that there were no Striga attachment to the roots of those that did not record Striga emergence.

Yield loss assessment due to Striga infestation
Twelve RIL’s were selected based on their good agronomic traits on the field (white seed coat, big size and early maturity). The 12 consisted of five Striga resistant lines, five Striga susceptible lines and the two parents (IT97K-499-35 and Sanzi) as checks (Table 3).

The experiment was designed as a split plot with four replications. The Striga treatments (infested and no infestation) were the main plots while the 12 lines were the sub plots. The soil used to fill the pots were steam sterilised at 100°C to get rid of all Striga seeds. A metallic barrel was used for the sterilization of the soil. A wire mesh was fitted at 1/3 of the length of the barrel from the bottom. This served as a separator between the soil and the water. The setup was placed on fire. Water was poured in the barrel to fill up to the level where the wire mesh is fitted, jute sack was then laid over the wire mesh before filling the remaining two thirds with soil. The soil was covered with jute sack. The steam generated from the boiling water was allowed to pass through the soil for an hour and half to heat up the soil up to 100°C. The fire was put off upon attaining the 100°C to allow the soil to cool down. The soil was then scooped and spread on a plastic sheet to allow it to further cool down under shade before filling the plastic pots.
Forty-eight pots were infested with 5 g with S. gesnerioides. The other forty-eight pots were not infested with Striga seeds. All the pots were watered to field capacity and allowed to drain for 24 h before planting. The pots were irrigated as when it is needed and kept weed-free through hand pulling. Monitored spray was done against insects. From thirty-five days after planting, Striga emergence was recorded on daily based on visual observation. The other agronomic data collected included first day of flowering, days to 50% flowering, plant height, number of peduncles per plant and days to maturity.
The post-harvest data collected included dry pod weight, grain weight, number of seeds per pod, hundred seed weight as well as fresh and dried biomass weight. The dried biomass was obtained after drying in an oven for 24 h to a constant weight.
Yield loss assessment due to Striga infestation was estimated using the formula:

YL: Yield losses.
Statistical analysis
All field data collected were subjected to analysis of variance (ANOVA) using the GenStat analytical software (version 12.1.0.3338). Varietal means were compared using least significant difference at 5% level of probability (LSD 5%).
RESULTS
Cowpea RILs reaction to natural Striga gesnerioides in the field screening
The result showed that sixty-six (66) RILs out of the 251 (26.29%) used for this trial were resistant. (Table 4). Striga plants emerged from the soil of the plots containing susceptible RILS.


Reaction of cowpea RILs to artificial Striga gesnerioides in pot experiments
The results of artificial inoculation showed that 27 RILs were found to be resistant (no Striga emergence or Striga attachment) whiles 39 were susceptible (having Striga emergence or Striga attachment at the roots level) (Table 5). The number of days to flowering and maturity varied from 35 to 55 and 60 to 86, respectively.

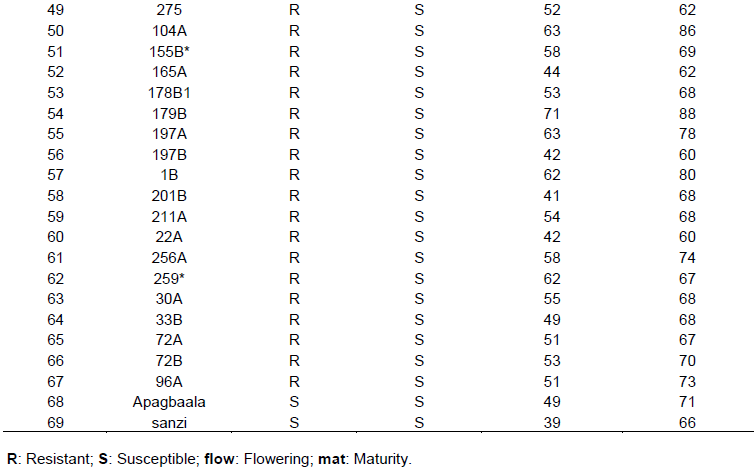
Evaluation of Striga promising lines in yield loss
Genotypes varied significantly in terms of days to 50% flowering and maturity under both infested and not infested (Table 6). Days to flowering and maturity varied from 41 to 55 and 63 to 73 days after planting. Under Striga infestation, the genotype 191 was the earliest to flower at 44 days after planting (Table 6). Under no Striga infestation, the genotypes 25 and 112A flowered earlier than the rest of the genotypes (41 and 43 days). 155A2 flowered 50 days after planting. The remaining genotypes were considered as medium maturity cultivars based on the days to flowering (43 to 49 days).
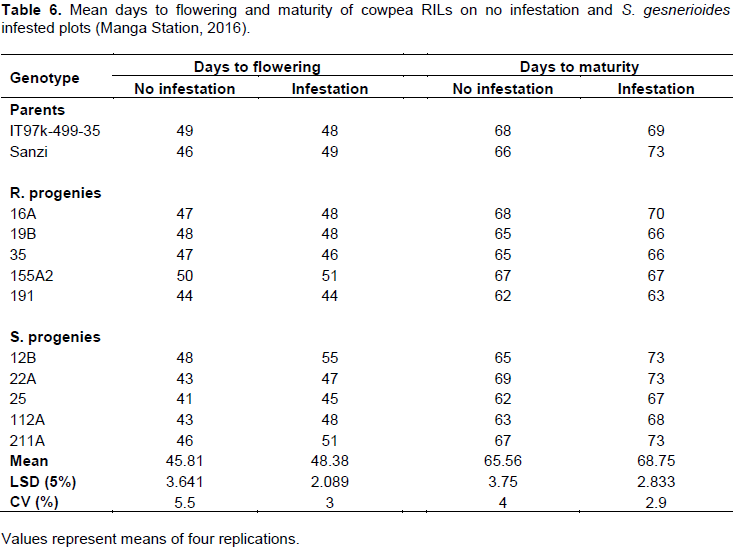
Under Striga infestation, all the resistant lines significantly (P<0.001) flowered and matured almost at the same time as in no Striga infested pots whiles the susceptible lines delayed in flowering and maturity (Table 6).
The resistant genotype 19B for instance flowered at 48 DAPS and matured at 65 and 66 days DAP in the non-infested and Striga infested pots, respectively.
The susceptible genotype 12B flowered at 48days after planting under no Striga infestation and 55 DAP under Striga infestation. The days to maturity were 65 and 73 days non-infested and Striga infested pots respectively (PË‚ 0.001).
Seed yield and dry biomass per hectare
The analysis of variance revealed significant differences between the progenies under Striga infestation and no Striga infestation (Table 9)
Among the RILs, 16A, under no infestation produced the highest grain yield (754.2 kg ha-1) followed by 25 (473.3 kg ha-1) and 19B (470.1 kg ha-1) (Table 7). The genotype, 155A2, a resistant cultivar recorded the lowest grain yield (320.1 kg ha-1). The cultivar 16A which recorded the highest yield under no infestation (754.2 kg ha-1) also recorded the significant yield under Striga infestation. (750 kg ha-1). Ironically, the susceptible cultivar 25, one of the highest grain producers under no Striga infestation (436.1 kg ha-1) also had one of the lowest grain yield under the infestation (338.6 kg ha-1). In general, the reduction in grain yield was higher in the susceptible progenies than the resistant ones.
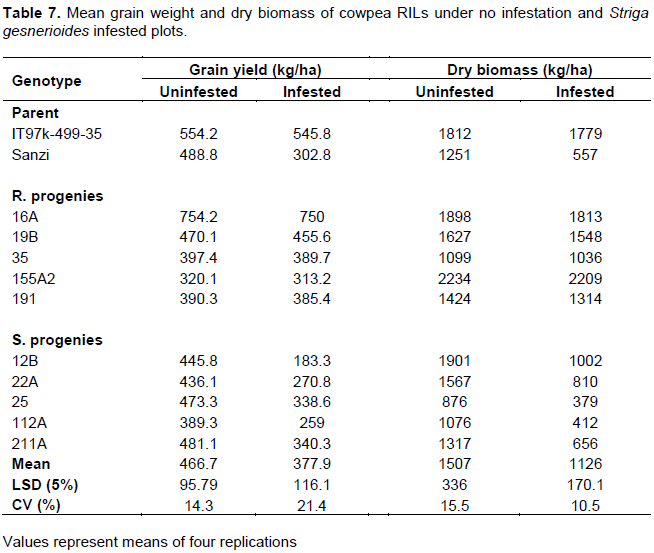
Dry biomass yield showed significant differences among the Striga infested (P Ë‚ 0.001) and non-infested conditions. The mean values of dry fodder yield were 1507 kg ha-1 under no Striga condition and 1126 kg ha-1 in the infested conditions. The progenies with the highest dry biomass under no infestation conditions were 155A2 and 12 B with 2234 and 1901 kg ha-1, respectively. The lowest fodder yield was recorded for the cultivar 25 with yield of 876 kg ha-1. The dry biomass yields for the other genotypes ranged from 1076 to 1812 kg/ha.
Under the Striga infestation condition, the genotype 155A2 still recorded the highest fodder likewise in the no infestation. The dry fodder yield of genotype 12B drastically dropped from 1901 kg ha-1 in the non-infested condition to 1002 kg ha-1 under the infested condition. The genotype 16A also recorded good production of fodder in both infested (1813 kg ha-1) and no infested condition (1898 kg ha-1).
Plant height and number of pods per plant
The number of pod per plant was significantly different in both infested (P Ë‚ 0.001) and non-infested environment. The analysis of variance indicated a significant difference for the plant height in both infested and non-infested environments.
The resistant plants were taller than the susceptible RILs under Striga infested condition (Table 8). The resistant parent IT97k-499-35 was the tallest plant (35.61 cm) followed by 16A and then 191 with plant heights of 31.63 and 31.53 cm, respectively. The susceptible cultivars 12B recorded the shortest plants height with 15.74 cm, in the no Striga infested pots. Striga susceptible genotypes were shorter compared to the resistant RILs.

Among the progenies, the highest mean number of pods per plant (10 pods) was recorded in the susceptible genotype, 22A, under no Striga infestation, but produced 7 pods under Striga infested condition. However, for the resistant RIL 19B, the mean number of pods was not affected when grown on Striga infested soils.
Grain yield and dry biomass loss due to Striga gesnerioides
In general the grain and biomass yield loss were higher in the susceptible lines compared to the resistant RILs. For the resistant RILs the dry grain yield losses ranged from 4.5 kg ha-1 (0.55%) to 14.5 kg ha -1 (3.08%) (Table 9). In the susceptible genotypes, grain yield losses oscillated from 134.7 kg ha-1 (28.45%) to 262.5 kg ha-1 (58.88%).

The highest grain yield loss (58.88%) was recorded for the susceptible RIL 12B followed by the susceptible line 22A which registered 37.9% grain yield loss. Grain yield losses for the resistant progenies were found to be between 0.55% for the cultivar 16 A to 3.08% for the genotype 19B. The resistant line 16A also showed a lower yield loss (0.55%) than the resistant parent (1.51%).
Dry biomass losses for the susceptible progenies ranged from 889 kg ha-1 (47.29%) to 664 kg ha-1 (61.71%) for the cultivars 12B and 112A, respectively.
Similarly for the dry grain yield, the resistant RILs did not show any significant biomass losses. With regard to the biomass losses, the cultivar 155A2 performed better in both Striga infested (2209 kg ha-1) and non-infested (2234 kg ha-1) then to the resistant parent IT97K-499-35, and also recorded the least biomass loss (1.1%) (Table 9).
DISCUSSION
Field screening for cowpea genotypes resistant to Striga gesnerioides
The field study recorded high emergence of Striga gesnerioides per plot (243 shoots) of the susceptible lines and this is similar to observations in other studies (Carsky et al., 2003; Kamara et al., 2008). The high Striga emergence observed on the Striga Susceptible lines was an indication that the site was really a hot spot for S. gesnerioides and the field had been infested with high concentration of Striga seeds over the years. However, a rigorous screening of the 66 genotypes in artificially infested soils in pot experiments revealed that only 10.75% were truly resistant to Striga. A susceptible genotype could be heavily infested underground without any Striga emergence as a results of several factors. According to Kim et al. (2002), one of the major limitations of screening under natural infestation is the variability in Striga seeds dissemination and cultivars escaping infestation. Striga sp. seeds need warm stratification for a certain time at a right temperature (approximately 30°C) before the seeds start responding to germination stimulants (Matusova et al., 2004). The high interference such as soil and climatic factors observed in the field makes the field screening less accurate (Baptiste et al., 2013).
Pot screenings
Field screening under artificial infestation is not always practical due to the fact that it can cause Striga seeds spreading to novel regions and it is moreover not consistent because breeders do not have any control of the parasite density and distribution (Haussmann et al., 2000). Pot screening has been operative as an alternative technique to confirm uniform infestation of Striga seeds.
After the pot experiment, the number of resistant lines was reduced from sixty-six (66) RILs to twenty seven (27) RILs after the pots experiment. This is essentially due to the high level of infestation (five grams of Striga seed per pot), the uniformity and a better control of the environment. A previous study done by Baptiste et al. (2013), confirmed the reliability of the pot screening compared to field screening.
The increased number of susceptible recombinant inbred lines found among the 66 could also be implied that these genotypes though showed no emerged seedlings of Striga had Striga attached to their roots. According to Ba (1983), some cowpea genotypes stimulate the Striga to germinate and penetrate their root tissues, but the Striga fails to grow more.
After both field and pot screening for Striga resistance, and taking into consideration farmers preferred traits, the genotypes 16A, 19B, 35, 155A2 and 191 were identified as promising Striga resistant lines.
Striga infestation delayed the flowering and maturity of susceptible cowpea genotypes. The susceptible genotypes also experienced huge reduction in grain yield and dry biomass in the Striga infested environment compared to when they were grown under no Striga environment. The study also confirmed that Striga infestation induces stunted growth hence the significant reduction of plant height at 50% flowering recorded for the susceptible genotypes. It also had an effect on the production of number of pods per plant. These data corroborated with previous studies (Press, 1995; Alonge, 1999; Gworgwor et al., 1991), which produced similar results. The stunted growth of genotypes, 12B, 22A, 25, 112A and 211A, could be attributed to the competition between the host and Striga for resources. The reduced vegetative growth of the susceptible varieties resulted in reduced leaf area, photosynthetic capacity and therefore affected flowering, podding and seed production (Alonge, 1999). According to Press (1995), the lower biomass accumulation by the susceptible genotypes could be the result of competition among the host and the weed for solutes, as well as carbon, water and minor rate of photosynthesis in the leaves of Striga infested plant. The reduced photosynthesis might have resulted in lower number of pods per plant and translocation of photosynthates to the sink.
Graves et al. (1992), showed that the low chlorophyll content which characterizes susceptible genotypes may account for the reduced development of the susceptible cowpea genotypes causing a decrease in both grain and biomass yield. The low biomass yield could also be attributed to the reduced shoot growth of the susceptible genotypes. The same phenomenon has also been reported for cereals infected with Striga hermonthica and for some cowpea genotypes infected with S. gesnerioides (Graves et al., 1992).
The resistant cultivars showed a relatively good growth compared to the susceptible lines in the infested pots. The relative good growth and the reduced export of assimilate to the weed would have ensured sufficient biomass accumulation and seed development as suggested by Gworgwor et al. (1991) on S. gesnerioides. The superior growth of some genotypes like 16A, 19B, 35, 155A2 and 191 indicated the positive relationship between crop vigour and crop performance even in Striga infested pots.
Grain and biomass loss due to S. gesnerioides
This current study has shown that all the resistant cowpea cultivars (16A, 19B, 35, 155A2 and 191) exhibited lower grain and dry biomass loss compared to the susceptible ones (12B, 22A, 25, 112A and 211A) indicating that these cultivars could play an essential role in controlling Striga in the endemic areas.
The susceptible genotypes recorded an average yield loss of 37.66% for dry grain yield which is quite consistent with the yield loss of 31±4% with a range of 26 to 65% observed by Aggarwal and Ouedraogo (1989). According to these authors, the loss could be attributed exclusively to the genotype effect as a consequence of Striga direct parasitism of susceptible cowpea lines (Muleba et al., 1996). S. gesnerioides diverts the host nutrient into themselves via the haustorium which establishes contact with the host tissues (xylem and phloem) (Okonkwo and Nwoke 1978; Okwonkwo, 1966). Consequently, this competition among host and parasite for water, and essential metabolites could be the explanation for the yield loss (Stewart and Press, 1990). Setty and Nanjapp (1985) and Kuijt (1969), reported that the osmotic pressure of the parasite is higher in both leaf and root than its host making the Striga more competitive. The use of high yielding Striga resistant varieties coupled with good agronomic practices can therefore help to reduce the yield losses in soil infested with Striga in the traditional farming systems.
CONCLUSION
The study revealed different reactions of cowpea RILs to S. gesnerioides during the field and the pot experiments. Out of the 251 RILs used, 27 RILS were found resistant similar to the resistant parent (IT97K-499-35), whiles 224 RILs were susceptible.
Yield loss assessment showed that the Striga resistant genotypes suffered less yield loss compared to the susceptible ones and therefore resistant genotypes can be one of the best means to minimize yield loss. These genotypes that expressed complete resistance are potential lines that will serve as resistant genotypes. The latest discovery of new sources of resistance to Striga provides an excellent way to supply farmers with new genotypes to replace their susceptible varieties.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abudulai M, Seini SS, Haruna M, Mohammed AM, Asante SK (2016). Farmer participatory pest management evaluations and variety selection in diagnostic farmer field Fora in cowpea in Ghana. African Journal of Agricultural Research 11:1765-1771. |
|
|
Aggarwal VD, Ouedraogo JT (1989). Estimation of cowpea yield loss from Striga infestation. Tropical Agriculture 66:91-92. |
|
|
Ajeigbe HA, Saidou AK, Singh BB, Hide O, Satoshi T (2012). "Potentials for cowpea (Vigna unguiculata) for dry season grain and fodder production in the Sudan and Sahel zones of West Africa," in Innovative Research Along the Cowpea Value Chain, eds O. Boukar, O. Coulibaly, C. A. Fatokun, K. Lopez, and M. Tamo [Ibadan: International Institute of Tropical Agriculture (IITA)] pp. 189-202. |
|
|
Alonge SO (1999). Effects of Alectra vogelii and Striga gesnerioides infestations on the growth, yield and grain chemical composition of cowpea (Vigna unguiculata (L.) Walp) varieties in the Nigerian savannah. Ph.D. Thesis, Department of Biological Sciences, Ahmadu Bello University, Zaria, 325 p, submitted for publication. |
|
|
Alonge SO, Lagoke STO, Ajakaiye CO (2004). Cowpea reactions to Striga gesnerioides I: Effect on growth. Crop Protection 24: 565-573. |
|
|
Ba AT (1983). Striga resistance screening of some cultivars of pearl millet, sorghum, maize and cowpea. In: K.V RAMAIAH., VASUDEVA RAO and M.J. (eds.). Proceedings of the Second International Workshop on Striga, P 47. Pantancheru, India. |
|
|
Baptiste TJS, Jéremy, Ouédraogo T, Tongoona RMP, Sibiya J, Makanda I, Drabo I (2013). Identification of new sources of resistance to Striga gesnerioides in cowpea germplasm Plant Breeding 132:330-336. |
|
|
Berhane S (2016). Review on Striga Weed Management International Journal of Life Sciences Scientific Research 2(2):110-120. |
|
|
Bisikwa J, Kawooya R, Ssebuliba JM, Ddungu SP, Biruma M, Okello DK (2014). Effects of plant density on the performance of local and elite cowpea varieties in Eastern Uganda. African Journal of Agricultural Science and Technology 1:28-41. |
|
|
Carsky RJ, Akakpo C, Singh BB, Detongnon J (2003). Cowpea yield gain from resistance to Striga gesnerioides in Southern Benin. Experimental Agriculture 39(3):327-333. |
|
|
Ddamulira G, Santos C A F, Obuo P, Alanyo M, Lwanga CK (2015). Grain yield and protein content of Brazilian cowpea genotypes under diverse Ugandan environments. American Journal of Plant Sciences 6:2074-2084. |
|
|
Dube E, Fanadzo M (2013). Maximizing yield benefits from dual-purpose cowpea. Food Sec. 5:769-779 |
|
|
Food and Agriculture Organization of the United Nations Statistics Division (FAOSTAT) (2018). |
|
|
Graves JD, Press MC, Smith S, Stewart GR (1992). The carbon economy of the association between cowpea and the parasitic angiosperm Striga gesnerioides. Plant, Cell and Environment 15:283-288. |
|
|
Gworgwor NA, Bdiya BS, Weber HC (1991). Screening cowpea varieties for resistance to Striga gesnerioides in the semi-arid zone of Nigeria. In: K. Weymann., L.J. Musselman. (Eds.), Progress in Orobanche Research, Obermarchtal, Germany, 19–22 August, 1989, pp. 322-328. |
|
|
Haussmann BIG, Hess DE, Welz HG, Geiger HH (2000). Improved methodologies for breeding Striga-resistant sorghums. Field Crops Research 66:195-221. |
|
|
Kamara AY, Chikoye D, Ekeleme F, Omoigui LO, Dugje Y (2008). Field performance of improved cowpea varieties under conditions of natural infestation by parasitic weed Striga gesnerioides. International Journal of Pest Management 54(3):189-195. |
|
|
Kim SK, Adetimirin VO, Thé C, Dossou R (2002). Yield losses in maize due to Striga hermonthica in West and Central Africa, International Journal of Pest Management 48(3):211-217. |
|
|
Kuijt J (1969). The Biology of Parasitic Flowering Plants. University of California Press, Berkley 246 p. |
|
|
Lane JA, Child DV, Reiss GC, Entcheva V, Bailey JA (1997). The gene for gene relationship in plant-parasite interactions. Proceedings of conference on crop resistance to parasitic weeds. I.R, Crute., E. B,Holub., and J.J, Burton. (ed.). CAB International, Wallingford, Oxon, United Kingdom plants 5:81-97. |
|
|
Matusova R, van, Mourik T, Bouwmeester HJ (2004). Changes in the sensitivity of parasitic weed seeds to germination stimulants. Seed Science Research 14:335-344. |
|
|
Mohamed KI, Musselman LJ (2008). Taxonomy of agronomically important Striga and Orobanche species. In: Progress on Farmer Training in Parasitic Weed Management (R,Labrada., ed.), Rome: FAO. pp. 7-14. |
|
|
Muleba N, Ouédraogo JT, Drabo I (1996). Yield stability in relation to Striga resistance in cowpea production in West and Central Africa. African Crop Science Journal 4:29-40. |
|
|
Okonkwo SNC (1966). Studies on Striga senegalensis II. Translocation of C-14 labelled photosynthate. Urea C-14 and sulphur-35 between host and parasite. American Journal of Botany 53:142-148. |
|
|
Okonkwo SNC, Nwoke FIO (1978). Initiation, development and structure of the primary haustorium in Striga gesnesrioides (Scrophulariaceae). Annals of Botany 42:455-463. |
|
|
Omoigui LO, Kamara AY, Alunyo GI (2017). Identification of new sources of Striga gesnerioides in cowpea Vigna unguiculata accessions. Genetic Resources and Crop Evolution 64(5):901–911. |
|
|
Omoigui LO, Kamara AY, Massawe FS, Ishiayaku MF, Boukar O, Alabi SO, Ekeleme F (2009). Evaluation of cowpea genotypes for their reactions to Striga gesnerioides I the dry Savannah of Northeast Nigeria. African crop Science conference Proceeding 8:273-278. |
|
|
Parker C (2012). Parasitic weeds: A world challenge. Weed Science 60:269-276. |
|
|
Press MC (1995). How do the parasitic weeds Striga and Orobanche influence host carbon relation? Aspects of applied Biology 42:63-70. |
|
|
Setty TKP, Nanjappa HV (1985). Studies on analysis of root exudates of Sorghum and osmotic potential of Sorghum and Striga. Abstracts of papers. Annual Conference of India Society of Weed Science 47 p. |
|
|
Singh BB (2005). Cowpea [Vigna unguiculata (L.) Walp. In: R.J, Singh, P.P, Jauhar (eds) Genetic Resources, Chromosome Engineering and Crop Improvement. CRC Press, Boca Raton, FL, USA 1:117-162. |
|
|
Singh BB, Chambliss OL, Sharma B (1997). Recent advances in cowpea. In: B.B. Singh, D.R. Mohan Raj, K.E. Dashiel and L.E.N. Jackai (eds.). Advances in cowpea research. Co-publication of International Institute of Tropical Agriculture (IITA) and Japan International Research Center for Agricultural Sciences (JIRCAS), Ibadan, Nigeria pp. 30-49. |
|
|
Stewart GR, Press MC (1990). The physiology and biochemistry of parasitic angiosperms. Annual Review of Plant Physiology and Plant Molecular Biology 41:127-151. |
|
|
Tanzubil PB, Zakariah M, Alem A (2008). Integrating Host Plant Resistance and chemical control in the management of cowpea pests. Australian Journal of Crop Science 2(3):115-120. |
|
|
Timko MP, Ehlers JD, Roberts PA (2007a). Cowpea. In: C, Kole (ed) Genome Mapping and Molecular Breeding in Plants, Volume 3, Pulses, Sugar and Tuber Crops, Springer Verlag, Berlin Heidelberg pp. 49-67. |
|
|
Timko MP, Gowda BS, Ouedraogo JT, Ousmane B (2007b). Molecular markers for analysis of resistance to Striga gesnerioides in cowpea. In: G, Ejeta and J, Gressel (eds) Integrating New Technologies for Striga control, towards ending the which-hunt. World Scientific Publishing Co. Pte Ltd, Singapore pp. 115-128. |
|
|
Westwood JH, Yoder JI, Timko MP, DePamphilis CW (2010). The evolution of parasitism in plants. Trends in Plant Science 15:227-235. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


