A study was carried out to study the combining ability effects of diallel cross hybrids for grain yield, agronomic traits and reaction to grey leaf spot (GLS). Forty five experimental diallel cross hybrids made from ten quality protein maize (QPM) inbred lines with varying level of resistance to GLS were evaluated along with three checks at Bako and Jima Research Centers during 2014/2015 cropping season. Analysis of variance showed significant variation (P≤0.05) among genotypes and between environments. This depicted the existence of genetic variation among genotypes in all studied traits. Mean squares characterized by general and specific combining ability effects were significant for the most traits and this suggested that both additive and non-additive gene actions have the contribution in the expression of the traits. However, the ratio of General combining ability (GCA) to specific combining ability (SCA) sum of squares were greater than unity, this revealed that there was preponderance of additive gene action in the expression of all the traits under study. Among inbred lines studied P6 and P10 had a desirable GCA effects for grain yield whereas P1, P4 and P10 were the best general combiners for GLS resistance. Furthermore, P10 was identified as good general combiner for grain yield as well as GLS is therefore recommended to be used in breeding programs with a purpose of developing both high yielder as well as GLS disease resistant genotypes. Crosses, P2×P8 and P4×P6 showed the most desirable performances and SCA effects for grain yield. Grain yield showed positive correlation between ear per plant, plant and ear height, and ear position whereas negative correlation were observed with GLS, days to anthesis, days to silking, plant aspect and ear aspect traits. The information which is generated in this study could be helpful to develop high yielding maize varieties with good agronomic traits as well as GLS resistance.
Maize [Zea mays (L.) 2n=2x=20] is one of the major cereal crops in terms of production and productivity in Ethiopia. It grows on 2.1 million hectares of land with total production of 7.2 million tonnes annually with the national average productivity of 3.42 tonnes per hectare (CSA, 2015). Maize is considered as a widely adaptable crop that is grown over a range of agro-climatic zones and suitable for diverse environments and can grow up to 2,600 masl from moisture deficit semi-arid lowlands, mid-altitude and highlands to moisture surplus areas in the humid lowlands, mid-altitudes and highlands of the country (Legesse et al., 2012).
In the developing countries particularly in Africa, South America and Asia, people consume maize as an important staple food and derive their protein and calories requirements from it (Prasanna et al., 2001). However, naturally maize has protein deficiency of the two essential amino acids of lysine and tryptophan. The opaque-2 gene which is responsible for the enhancement of lysine and tryptophan can be incorporated into any population, inbred lines or any other germplasm of maize through modified backcrossing recurrent selection program (Vasal et al., 1980). The modified opaque-2 maize which had hard endosperm and vitreous kernels is known as quality protein maize (QPM). Hence, nutritional superiority over normal maize is well established. Therefore, development of quality QPM maize would greatly reduce malnutrition problems of subsistence and resource poor farmers and low-income people that depend on maize as their staple food (Leta et al., 2003). Despite some efforts made so far to improve quality of maize (Twumasi et al., 2012), enhancing yield potential with having good agronomic trait and disease resistant QPM maize varieties are still the major problems that needs attention in Ethiopia.
Disease is the most important production challenge and creates a series of problems from year to year in the most part of maize growing agro ecologies of the country. Around 65 maize diseases have been recorded in Ethiopia (Tewabech et al., 2012). Among these diseases, gray leaf spot (GLS) caused by Cercospora zeae-maydis is the most important threat to maize production in the country (Dagne et al., 2001) and can create yield reduction of up to 37% in susceptible varieties under artificial disease pressure (Dagne et al., 2004). The most recommended control measure of the disease in maize is the use of relatively resistant or tolerant cultivars (Daniel et al., 2008). Hence, use of host plant resistance (HPR) is considered as the best option to alleviate GLS disease pressure. Identification of inbred lines which are high yielding as well as GLS resistant is a pre option to solve the problem.
Combining ability could help to provide information in the selection of inbred lines and desirable cross combination (Falconer and Mackay, 1996). The general combining ability (GCA) helps to evaluate the contribution of an inbred line to the hybrid performance and population improvement, whereas specific combining ability (SCA) is utilized to identify cross combination with superior performance. GCA indicates the additive gene action whereas SCA indicates non-additive gene actions mainly a function of dominance effects (Sprague and Tatum, 1942). So far, combining ability studies of QPM inbred lines conducted by the Gudeta (2007) and Demissew (2014) indicated both GCA and SCA effects to be significant and important for grain yield and most other studied traits. However, Melkamu et al. (2013) reported the preponderance of SCA or non-additive gene action for grain yield and most other studied traits. In Ethiopia studying combining abilities effects for newly developed QPM inbred lines to grain yield, agronomic traits and reaction to disease have been limited. This study was, therefore, undertaken to estimate magnitude of combining ability effects and the relationship of grain yield, agronomic and GLS traits.
Description of the study areas
The experiment was conducted during the 2014/2015 main season at Bako National Maize Research Center (BNMRC) and Jima Agricultural Research Center (JARC). Bako National Maize Research Center is located in East Wellega Zone of the Oromia National Regional State, Western Ethiopia at an altitude of 1650 m above sea level (masl). It lies between 9°06' North latitude and 37°09' East longitude in the sub-humid agro-ecology of the country. During the experimental season, the rainy season covered the period from May to October and maximum rainfall is received in the months of July and August and the average annual rainfall was 944.4 mm. The mean, minimum and mean maximum temperature was 12.3 and 29.8°C, respectively. The soil is classified under the Nitosol order. Naturally, this area is exposed to high maize foliar disease pressure and mostly used as a hot spot area to screen new maize genotypes for the most foliar diseases.
Jima Agricultural Research Center is located in Oromia Regional State South West part of Jima Zone at an altitude of 1750 m above sea level (masl). It lies between 7°46' North latitude and 36°00' East longitude in the sub-humid agro-ecology of the country. During the experimental season, the average annual rainfall is 1536 mm. The mean minimum and mean maximum temperature is 11.2°C and 25.9°C respectively. It is the most recommended area to evaluate new maize genotypes for GLS since it is a hot spot area for the most foliar diseases.
Experimental materials
The inbred lines used in this study were obtained from Bako National Maize Research Center, Maize Protection Research Section. Based on per se performance evaluation of GLS and other agronomic traits, ten inbred lines were selected (Table 1). The lines were categorized as resistant, moderately resistant, susceptible and highly susceptible. Selected lines were crossed in a 10×10 half-diallel mating design at Bako National Maize Research Center during 2014 cropping season and three commercial check hybrids (BHQPY545, BH546 and BH547) were used in the trials. All these checks are widely adapted in mid-altitude agro-ecologies. BHQPY545 is a popular quality protein maize hybrid whereas BH546 and BH547 are recently released high yielder non quality protein maize hybrids.
Experimental procedure
The 45 F1 progenies derived from the diallel cross of 10 selected inbred lines along with three commercial hybrid checks (BHQPY545, BH546 and BH547) were planted in 6 × 8 alpha-lattice designs with two replications at Bako and Jima research centers. There were two rows per plot with row length of 5.1 m spaced 0.75 m apart and plant-to-plant distance of 0.3 m. At planting, two seeds were planted per hill, and then thinned to one plant per hill after two weeks of emergence to attain a population density of 44,444 plants per hectare. Nitrogen (N2) and diammonium phosphate (P2O5) fertilizers were applied at the recommended rates of 92 kg per hectare (kg/ha) and 69 kg/ha, respectively. P2O5 was applied at planting and N2 was applied in two splits: At planting and knee height. All other agronomic management practices such as weeding and hoeing were done based on research recommendations for the areas.
Data collection
Yield and agronomic performance
Data collected include: number of plants per plot was recorded at emergence and harvesting time, days to anthesis and silking (days): Taken during 50% of plants in a plot shed pollen and 50% of the plants in a plot showing 2 to 3 cm long silk emergence, anthesis-silking interval (days): It was calculated by subtracting from days of silking to days of anthesis, plant height (cm) was measured in centimeter from ten randomly selected plants as a distance from the ground level to the first tassel branch and for ear height, from the ground level to the upper most ear bearing node. Ear position (ratio): It was calculated as the ratio of ear height to plant height. At harvest time; number of ears per plant was calculated from the total numbers of ears harvested in a particular plot divided by the total number of plants harvested and grain yield (tonnes per hectare) was recorded the weight of the ears per plot and this was adjusted to 12.5% moisture content to estimate grain yield per hectare. Plant aspect was recorded by observing overall phenotypic appearance of the plant in a plot by using 1 to 5 scoring scale; where 1 = excellent and 5 = poor. Ear aspect was recorded by observing overall phenotypic appearance of the ears in a plot at harvesting time by using 1 to 5 scoring scale; where 1 = excellent and 5 = poor.
Reaction to gray leaf spot
Disease severity (scale): Disease severity was rated using 1 to 5 scoring scales based on Pusa Campus (2012) procedure. 1 = very slight to slight infection, one or two few scattered lesions on lower leaves, 2 = light infection, moderate number of lesions on lower leaves only, 3 = moderate infection, abundant lesions are on lower leaves, few on middle leaves, 4 = heavy infection, lesions are abundant on lower and middle leaves, extending to upper leaves. 5 = very heavy infection, lesions abundant on almost all leaves plants prematurely dry or killed by the disease.
Statistical analysis
The collected data were analyzed using the lattice design procedure of SAS statistical procedure (SAS, 2004). For the analysis of variance, test genotypes were considered as fixed whereas replication and incomplete blocks within replication were considered as random. Mean separation was done using least significant difference (LSD). Analysis of combining abilities were made using the application of diallel-SAS procedure (Zhang et al., 2005) computer program following Griffing’s (1956) Method IV and Model I (fixed model). Tests of significance of the combining ability effects and their differences were made using t-test. The relationships between yield, agronomic traits and GLS were determined using the Proc Corr procedure in SAS (SAS, 2004). The correlation was determined using across location mean data. The phenotypic relationships among the traits were determined using the correlation coefficient, r and the significance of correlation coefficients was determined using t-test at alpha level of 5 and 1% probability level.
Analysis of variance
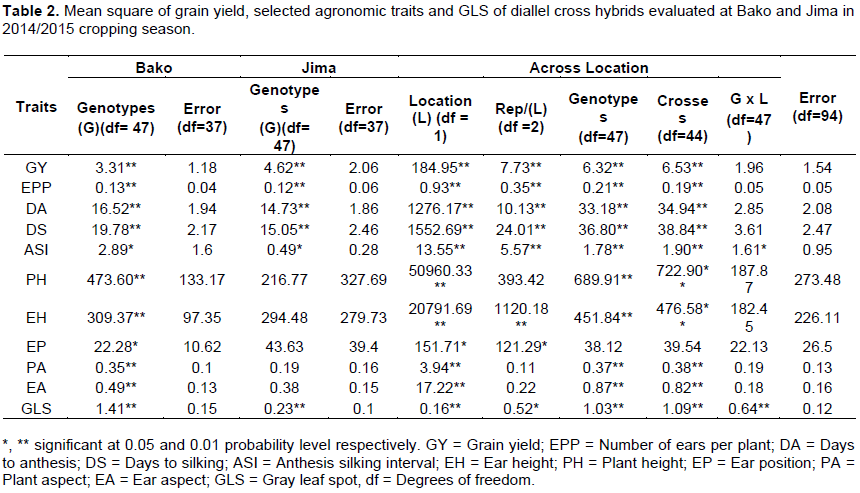
Analysis variance of genotypes showed significant mean square for all traits at Bako and except plant height, ear height, ear position, plant aspect and ear aspect other traits also revealed significant mean square at Jima location, (Tables 2). Combined analysis of mean square was significant at P≤0.05 and highly significance at P≤0.01 among locations, genotypes and crosses in all studied traits except ear position, this indicating that the presence of genetic variability among crosses. On the other hand, mean squares due to crosses × locations interaction for all studied traits, except ASI and GLS revealed non-significant variations (Table 2). Significant mean square for grain yield, agronomic traits and GLS were reported by Dagne et al. (2008), Legesse et al. (2009), Mousa et al. (2014), Hossein et al. (2014) and Mwai et al. (2015).
Mean performance of crosses
Mean performance of 45 experimental crosses and three commercial checks for grain yield, selected agronomic traits and GLS are presented in Table 3. The mean of grain yield (GY) for all crosses under this experiment ranged from 4.28 tonnes per hectare (P2 × P6) to 10.87 tonnes per hectare (P4×P6) with a mean value of 7.86 tonnes per hectare. Among the crosses, P4 × P6, P4 × P10, P5 × P6, P5 ×P10, P6 ×P10 and P9 ×P10 crosses showed significantly high yielder than the best check hybrid BH546 and these crosses also revealed greater than 10% yield advantages than the quality protein commercial maize hybrid BHQPY545 (Table 3). Number of ears per plant ranged from 0.94 for P3 × P8 to 1.76 for P9 × P10, with overall mean of crosses 1.34. The highest mean values for ears per plant were observed for crosses P9 × P10 and P3 × P6 as compared to the highest check BHQPY545.
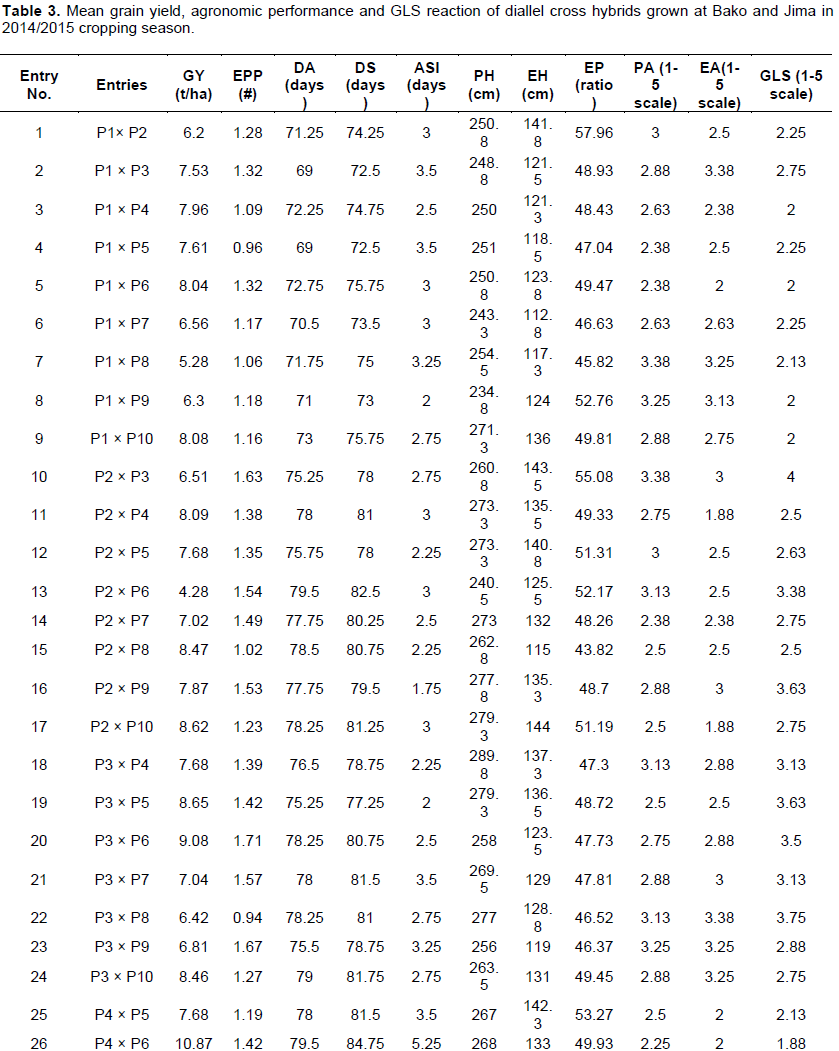
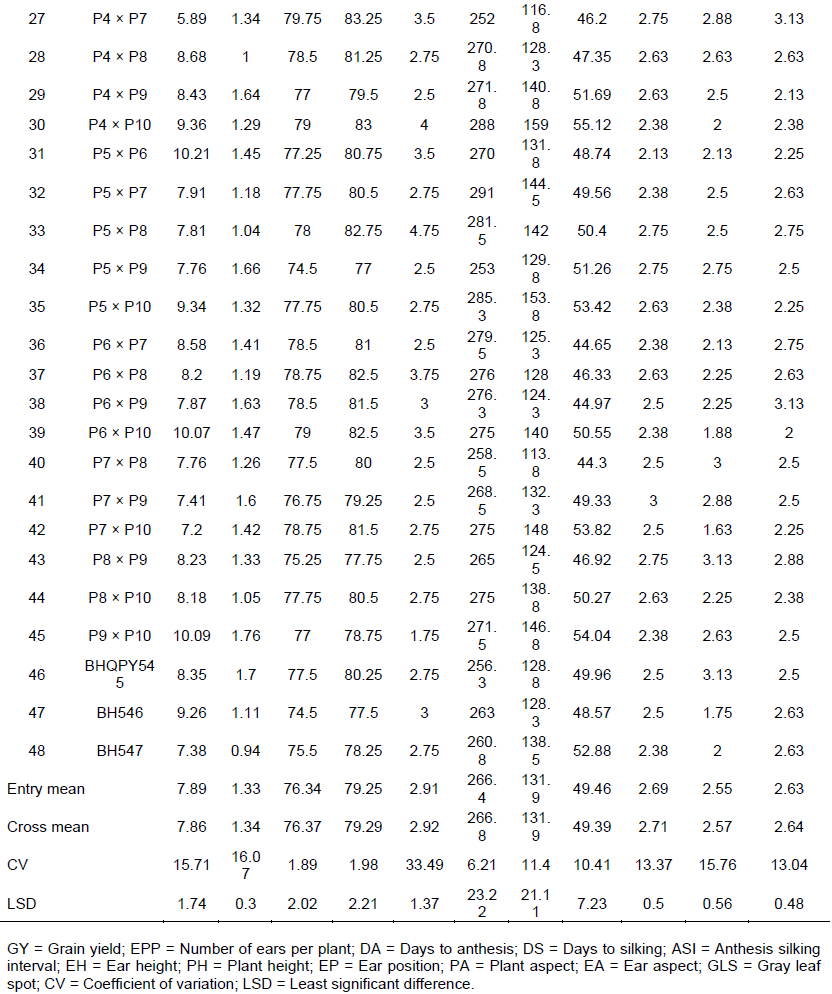
The number of days to anthesis (DA) ranged from 69.0 days (P1× P3 and P1 x P5) to 79.75 days (P4 × P7) with overall mean of 76.37 days and days to silking (DS) also ranged from 72.5 days (P1 × P3) to 84.75 days (P4 × P6). Crosses, P1 × P3, P1 × P5, P1 × P7, P1 × P2, P1 × P8, P1 × P4, P1 × P6, P1 × P10 and P5 × P9 revealed significantly short days to anthesis and silking as compared to hybrid check BH546, including these crosses 21 crosses exhibited earlier as compared to quality protein hybrid check BHQPY545. Crosses which showed longer number of days to anthesis and silking could be considered as late maturing types. Conversely, crosses which had shorter days to flowering could be regarded as early maturing types. Crosses which exhibited early anthesis and silking are desirable type of crosses especially in moisture stress environments since early type crosses can escape terminal moisture stresses during the growth stages (Banziger et al., 2004).
Anthesis-silking interval (ASI) ranged from 1.75 days (P2 × P9) to 5.25 days (P4 × P6). Eight crosses showed greater days to ASI than standard checks of BHQPY545 and BH547. In general, crosses which exhibited short anthesis-silking interval, indicates that the cross had short gaps between anthesis and silking days and it is a desired character for good seed setting since these crosses may showed anthesis-silking interva below 3 days could be found within an acceptable range.
Naturally, maize plant is proto-androus in which anthesis normally begins 1 to 3 days before the emergence of silks and 3 to 4 days after the silks emergence are ready to be pollinated (Poehlman, 1977). If, the gap between days of anthesis and silking is large, the viability of pollen would be minimized and abnormal fertilization might be takes place or fertilization may not be happened. In general, shorter ASI improves the pollen-silk synchronization, a major trait that is affected under moisture stress areas. The need for a shorter ASI to achieve high grain yield has been observed by Anderson et al. (2004), where the potential number of florets that could become grains was limited by the receptivity of the silks. In this regard, ASI has been widely used in indirect selection of higher grain yield under drought conditions (Banziger et al., 2004).
The mean value of plant and ear height for crosses varied from 234.75 cm for P1 × P9 to 291 cm for P5 x P7 and112.75 cm for P1 × P7 to 159 cm for P4 x P10 respectively. Twelve crosses showed significant plant height over check BHQPY545 and 18 crosses were showed shorter ear height than check hybrid BHQPY545. Among all the tested crosses, sixteen and seventeen crosses were showed shorter plant and ear height respectively over than high yielder check BH546. On the other hand, almost all high yielder crosses had tall plant and ear height as compared to standard checks (BHQPY545, BH546 and BH547). Ear position ranged from 43.82% for P2×P8 to 57.96% for P1×P2. In this study majority of crosses showed ear placement near to the mid part of the plant, indicating desirable character for lodging tolerance. In general, crosses which have shorter pant and ear height are desirable for lodging tolerance and to apply necessary management practices, whereas taller crosses are important to harvest high biomass yield that could be used as animal feed and source of fuel for poor farmers (Girma et al., 2015).
Plant and ear aspects are a visual evaluation of plants and ears before harvesting and at harvesting time respectively, by observing all over performance. The mean values of plant aspect ranged from 2.13 for P5×P6 to 3.38 for P2×P3 with overall mean of 2.71. Ear aspect ranged from 1.63 for P7×P10 to 3.38 for P1×P8 and P3×P8 crosses. Crosses such as P7 ×P10, P6 ×P10, P2×P10, P2×P10, P2×P4, P4×P6, P1×P6, P4×P10 and P4×P5 showed lower plant and ear aspect scores, indicating that these crosses had desirable characters such as clean, uniform and disease free.
Gray leaf spot (GLS) disease severity scores in combined analysis were showed highly significant variation. Mean value of GLS Severity was ranged from 1.88 (P4×P6) to 4.00 (P2 ×P3) with the overall mean value 2.64. Twenty four crosses were recorded higher disease severity as compared to hybrid check BHQPY545. Crosses such as P3 × P4, P3 × P7, P4 × P7, P6 × P9, P2 × P6, P3 × P6, P3 × P8 and P2 × P3 scored greater than 3 severity values, these crosses were vulnerable to GLS while P4 × P6, P1 × P4, P1 × P6, P1 × P9, P1 × P10 and P6 × P10 scored less than 2 disease severity and could be used in the development of resistant genotypes against grey leaf spot. All these resistant crosses were the results of P1, P4 and/or P10 as one of the parent and might be contribute good source of favorable genes. Generally, crosses which showed low severity values to GLS might be used as a source of resistant genes to develop GLS resistant QPM varieties, on the other hand crosses which had high severity score values considered as susceptible varieties.
Combining ability analysis
Mean squares due to general combining ability (GCA), specific combining ability (SCA), GCA × L (Location), SCA × L and error are presented in Table 4. The combined analysis of GCA showed a significant difference (p≤0.01) in all traits while mean squares of SCA were also significant for grain yield, days to silking, anthesis-silking interval, and plant and ear height and gray leaf spot. Therefore, significant GCA and SCA indicating both additive and non-additive gene action contribute the expression of the traits. However, mean squares due to SCA was non-significant for ear per plant, days of anthesis and ear position this showed that only additive gene actions contributed to the inheritance of these traits. The mean square of GCA × Location (GCA × L) interaction was significant only gray leaf spot, days of anthesis, plant aspect and ear aspect whereas the SCA × L interaction was non-significant for all traits (Table 4). Similar finding were reported by different investigators. Shushay et al. (2013) reported significant mean squares due to GCA of lines and SCA for grain yield and plant height, ear height and days to anthesis traits. In line by tester analysis, Melkamu et al. (2013) reported lines showed significant both GCA and SCA for all traits except for days to 50% tasseling, ear length, ear diameter, grain yield per plant and number of kernels per row. Mousa (2014) reported highly significant GCA and SCA mean squares for all traits studied. In a combined analysis of variance, Hossein et al. (2014) reported significant mean squares due to GCA and SCA for days to silking, plant height, ear height, 1000-kernel weight, ear length, number of rows per ear, number of leaf and kernel yield maize. In quality protein maize diallel analysis, Demissew (2014) reported GCA mean squares were significantly different (P < 0.01) for grain yield and all agronomic traits except EPP while mean squares due to SCA were significantly different for all traits excluding ASI and EPP. In addition to this significant GCA and SCA effects for GLS resistance were identified, resistance was conditioned by both additive and non-additive gene actions. Similar results were reported by Menkir and Ayodele (2005), Dagne et al. (2008), Leggesse et al. (2009), Nzuve et al. (2013) and Mwai et al. (2015).
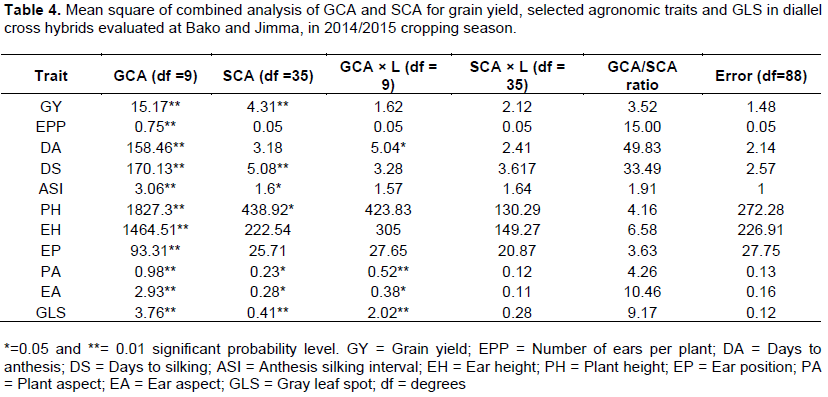
The ratio of GCA to SCA mean square was greater than unity for all studied traits. This suggests that additive gene action could be more important than non-additive gene action. The importance of additive gene effects in different traits has been reported in several investigators in different times (Dagne et al., 2008; Legesse et al., 2009; Nzuve et al., 2013; Hussien et al., 2014; Mwai et al., 2015).
Estimates of GCA and SCA effects
Significant differences in GCA effects were detected among lines for various traits (Table 5). The highest GCA effect for grain yield was observed from parent P10 (1.08 tonnes per hectare), followed by parent P6 (0.819 tonnes per hectare), whereas the lowest GCA value was observed form parent P1 (-0.9 tonnes per hectare), followed by parent P2 (-0.759 tonnes per hectare) and Parent P7 (-0.67 tonnes per hectare). Inbred lines, P6 and P10 were good combiners for grain yield since they showed positively significant GCA effects. These inbred lines can be desirable parents for synthesis of new synthetic varieties, since they may contribute favorable alleles in the improvements of the yield. On the other hand parents P3, P6 and P8 revealed positively significant GCA effects for ear per plant as well as parent P1 was good general combiner for days of anthesis, days to silking, plant height and ear height. These lines suggesting that they might be contribute favorable genes to develop early and short type of new varieties formation. Similarly, Parent P10 had also expressed highest significant GCA effects for plant height and ear height in the positive direction. Parent P7 and P8 were good general combiners in ideal ear placement formation. Parental lines such as P1, P3 and P8 showed positive significant GCA effects to the undesirable direction for plant and ear aspect. On the other hand, P6 and P10 revealed negative GCA effect which is the desirable direction which showed good plant and ear characters.
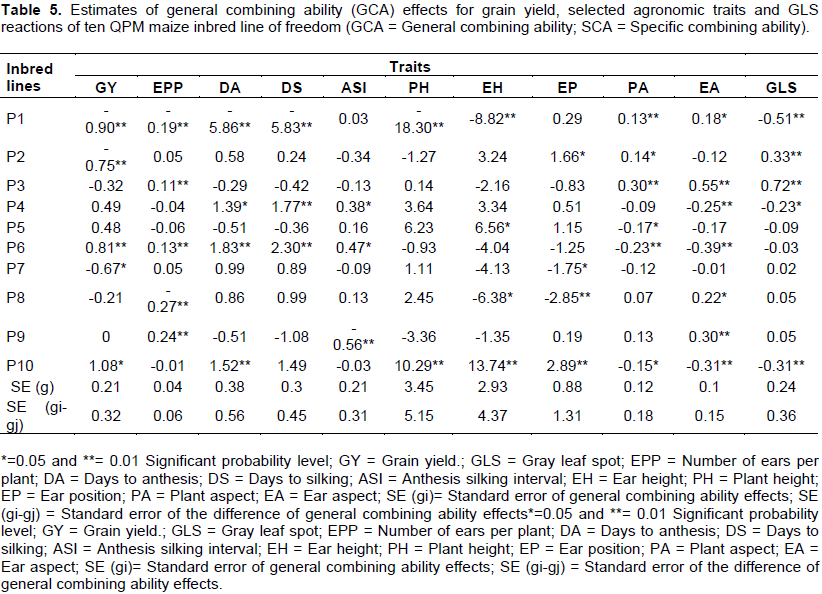
A line with negative GCA for gray leaf spot would be expected to be a useful source of resistance to the disease. Parents with significant negative GCA may contribute resistance in their crosses. Parents such as P1, P4 and P10 showed negatively significant GCA effects, this indicating that they could be good sources GLS resistant genes. It was recommended that these parents should be used in breeding programs with a purpose of incorporating GLS resistance genes with other desired traits. Especially, parental line P10 was a poor combiner for days of anthesis, plant height and ear height, but best for grain yield and GLS.
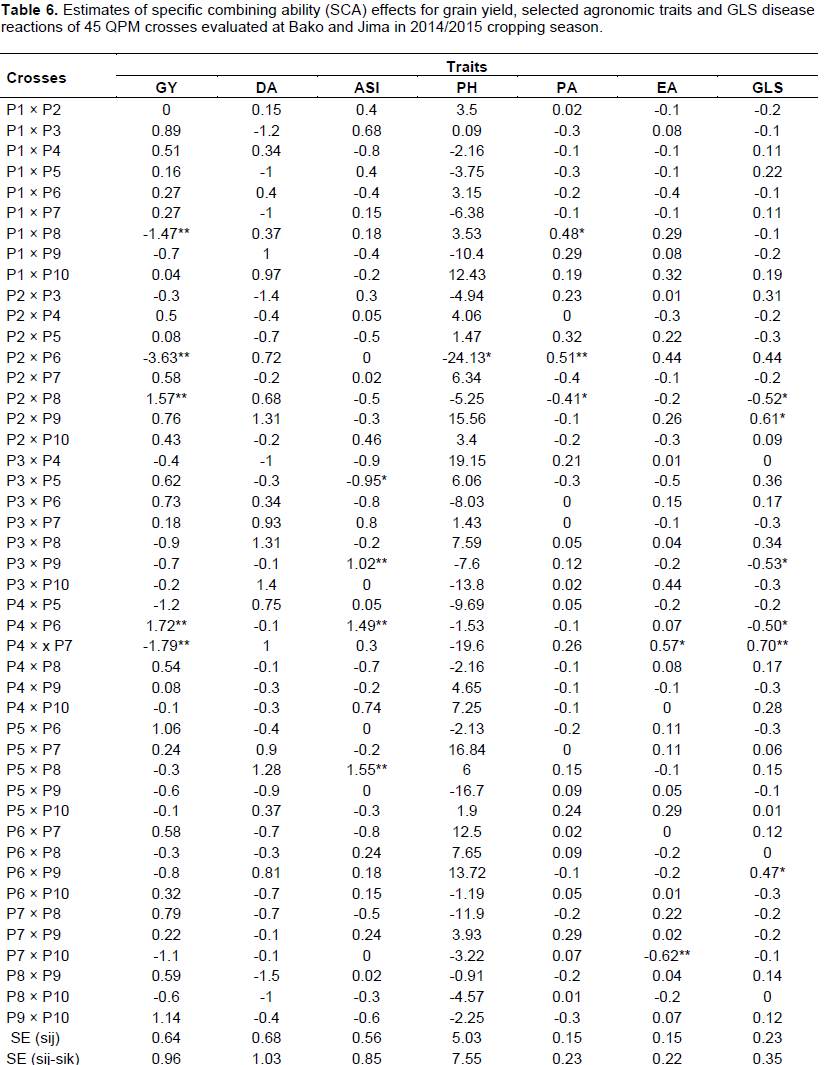
Significant SCA effects indicated that the crosses performed better or poorer than what would be expected based on GCA effects of the respective parents (Table 6). Crosses, P2 × P8 and P4 × P6 for grain yield, P3 × P5 for days of anthesis-silking interval, P2 × P6 for plant height, P2 × P8 for plant aspect and P7 × P10 for ear aspect depicted good specific combining ability effects, indicating the crosses showed top performance in each trait. In the case of GLS, P2 × P8, P3 × P9 and P4 × P6 crosses were showed significant and negative SCA effects, indicating the importance of non-additive gene action to be in the desirable direction for the improvement of the crosses against GLS reaction. Generally, crosses which revealed good SCA effects for the most studied traits could be help to choose good hybrids.
Correlation analysis
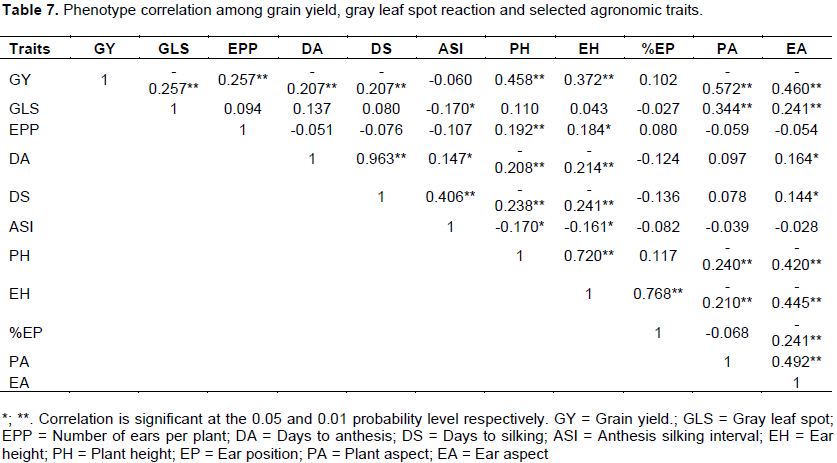
The result of Pearson correlation analysis indicated highly significant negative and positive relationship between grain yield and other traits observed (Table 7). Grain yield showed positive correlation between EPP, PH, EH and EP whereas negative correlation observed with GLS, DA, DS, PA and EA. However, non- significant correlation was observed with anthesis silking interval (Table 7). These results agreed with the work of Dagne et al. (2008) who reported grain yield had high positively correlated with plant height while contrast correlation between grain yield and GLS disease parameters. In addition to this the results were agreed with Daniel et al. (2008) reported negative correlation grain yield with disease severity. Non-significant-correlation was shown between gray leaf spot disease with days of anthesis, days of silking, plant height and ear height indicating the disease had less impact in reduction of days of flowering and height.
The combined analysis of variance showed that there was a significant variation among crosses for all studied traits. Crosses, such as P4 × P6, P5 × P6, P9 × P10, P6 × P10, P4 × P10 and P5 × P10 were high yielder hybrids as compare to the best hybrid checks BH546 and BHQPY546. Significant mean squares due to GCA for all traits and SCA for the most of the studied traits were detected, this indicating additive and non-additive gene effects contribute to the expression of the characters. Parental lines P6 and P10 were having good general combining ability for grain yield. On the other hand inbred lines P4 and P10 were good general combiner for GLS disease resistant hybrids development. Even though, there were no inbred lines that are good combiner for all traits studied, inbred line P10 is the best combiner for grain yield as well as GLS resistance. Hence, this inbred lines has a potential to be utilized for producing synthetic maize varieties which are high yielder and GLS resistant or can be used for other breeding purposes. Among the crosses P2 × P8 and P4 × P6 were found to be good specific combiners for grain yield and GLS. Hence, these parental combinations could be used for the development of high yielder as well as GLS resistant hybrids. Grain
yield showed positive correlation between EPP, PH, EH and EP traits whereas negative correlation were observed with GLS, DA, DS, PA and EA. Non-significant-correlation was showed between gray leaf spot disease with days of anthesis, days of silking, plant height and ear height indicating the disease do not have impact in days of flowering and height. Generally, this study identified inbred lines and crosses with desirable GCA and SCA effects for the studied traits. This had the possibility of developing desirable cross combinations and synthetic varieties through crossing and or recombination of inbred lines with desirable traits of interest.