Full Length Research Paper

ABSTRACT
Optimization of land use can be attained by incorporating technologies to crop production, such as the use of diazotrophic bacteria, fertilizers, and pesticides. Seed inoculation with Azospirillum is an alternative that favors the incorporation of green agriculture in regions of conventional farming, such as the Brazilian savannah (Cerrado). However, limited information is available about this bacterium’s contribution to agriculture when other technologies are also incorporated. This study evaluated the performance of maize hybrids inoculated, or not, with Azospirillum brasilense, with or without fungicide applications, and subjected to different nitrogen rates under Cerrado field conditions. Each factor analyzed contributes to the increased maize grain yield. The use of inoculants containing plant growth promoting bacteria is a good option to ensure high yield of maize. Still, nitrogen should not be replaced, neither totally nor partially, by seed inoculation with Azospirillum. Fungicide applications should be done, as required, during maize cycle. Moreover, specific maize breeding programs should consider the affinity between Azospirillum strains and maize hybrids, mainly for regions with nitrogen deficient soils, like Cerrado. Thus, by incorporating additional technologies, maize crop farmers can optimize land use and, consequently, reduce the expansion into new agricultural areas.
Key words: Foliar protection, nitrogen use, plant growth promoting bacteria, sustainability, Zea mays L.
INTRODUCTION
MATERIALS AND METHODS
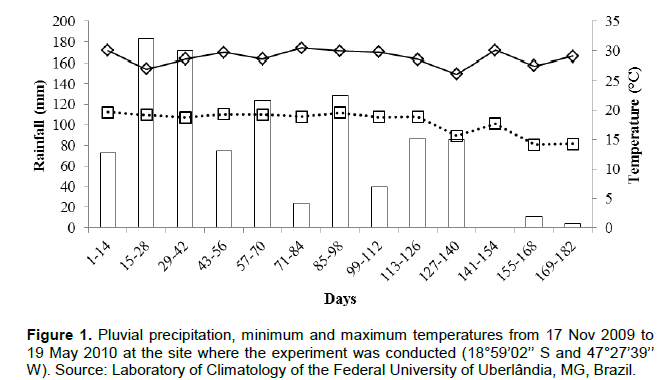
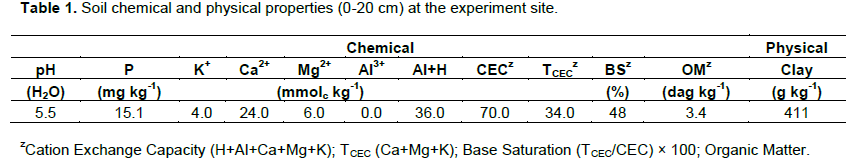
RESULTS
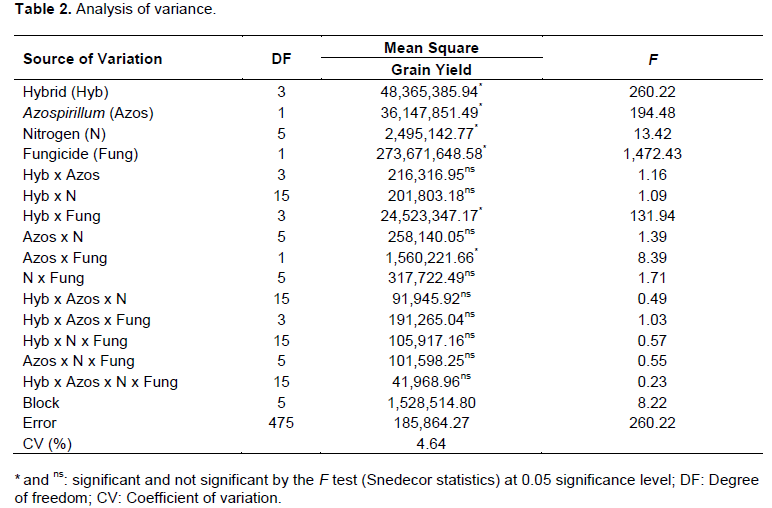
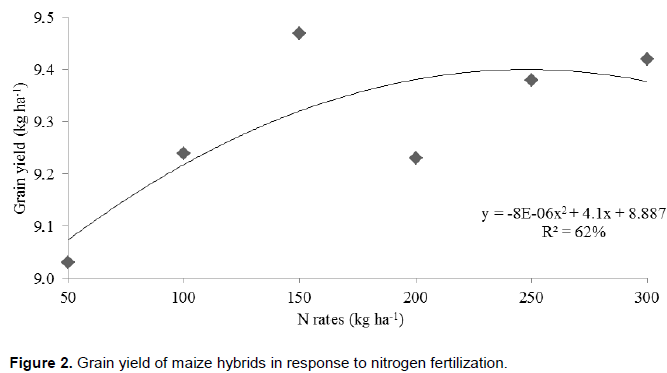
.png)
DISCUSSION
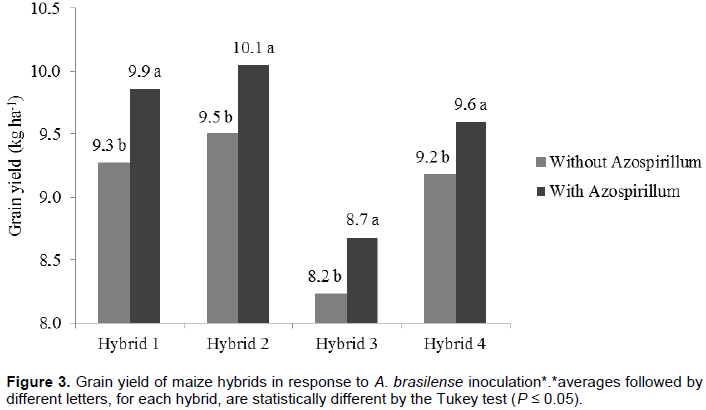
Therefore, inoculation enabled yield increases of maize crops growing under Cerrado conditions, which resulted from the affinity between Azospirillum and hybrids recommended for the region. Thus, inoculation allowed optimization of land use and even small and medium farmers (in low investment production systems) can obtain greater yields with this technology.
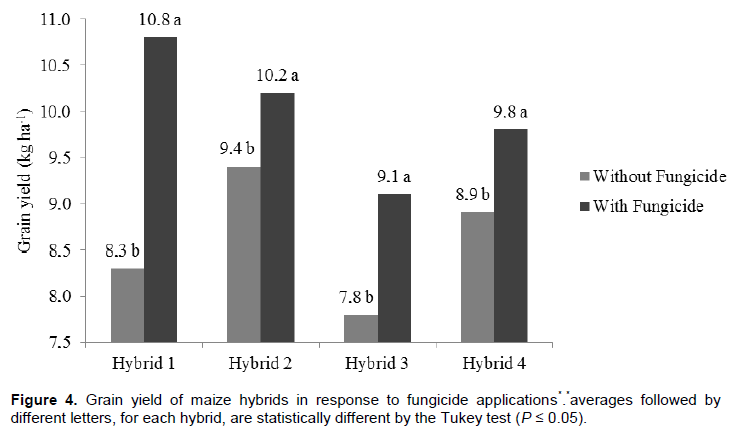
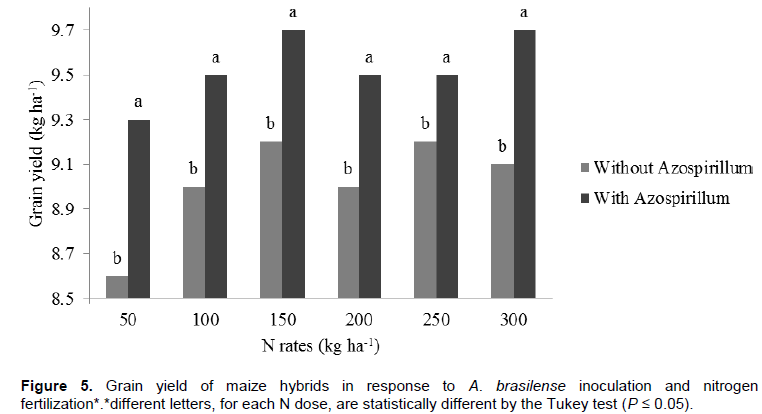
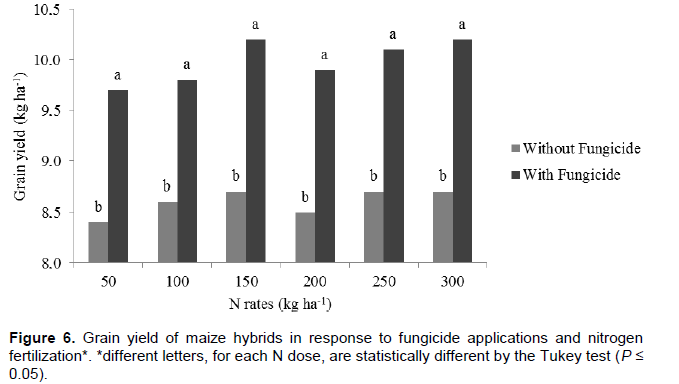
Increased production of maize can be attributed to the phytostimulatory effects of inoculation with Azospirillum, due not only to biological nitrogen fixation in the rhizosphere, but also to plant’s greater efficiency in water and nutrient uptake due to greater growth of root system provided by the production of plant growth promoting substances by the bacteria (Döbereiner, 1992; Reis et al., 2000; Cassán et al., 2008). Better nitrogen fertilizer use was observed when maize hybrids were inoculated with Azospirillum. From this result it can be inferred that Azospirillum inoculation enhances nitrogen use, although it does not replace it. Thus, even if part of the maize nitrogen demand is supplied by association with diazotrophic bacteria, reduction of nitrogen fertilizer rates is not recommended. This result, however, contrasts with the one reported by Hungria (2011), who found substantial reduction of nitrogen fertilization in maize plants inoculated with Azospirillum. Applying a nitrogen rate equivalent to half of that recommended for maize in Brazil (100 kg N ha-1), the researcher obtained grain yield of 7.8 t ha-1 (Hungria, 2011). However, it must be stated that such production was achieved only with the strain Ab-V5 (+ 54 kg N ha-1) in a single crop season. Besides the previously mentioned aspects, foliar protection affected maize yield as well. The hybrids obtained greater yield potential, reaching up to 10.2 t ha-1, after fungicide applications. This is certainly related to treated plants health. In treatments without fungicide application, hybrid photosynthetic activity may have been compromised, resulting in lower production. This ratifies the idea that investment in plant nutrition is jeopardized if correct phytosanitary management is not adopted. This statement is consistent with studies about effects of fungicide use to increase plant yield (Köhle et al., 2003).
CONCLUSIONS
CONFLICT OF INTERESTS
REFERENCES
|
Araújo GM, Haridasan M (1988). A comparison of the nutritional status of two forest communities on mesotrophic and dystrophic soils in central Brazil. Commun. Soil Sci. Plant Anal. 19:1075-1089. |
|
|
Bashan LE, Holguin G, Glick BR, Bashan Y (2007). Bacterias promotoras del crecimiento en plantas para propósitos agrícolas y ambientales. In: Microbiología agrícola: hongos, bacterias, micro y macrofauna, control biológico, planta-microorganismo (Eds.: R. Ferrera-Cerrato and A. Alarcon). Editorial Trillas, Mexico City, Mexico pp. 170-224. |
|
|
Bashan Y, Holguin G, Bashan LE (2004). Azospirillum-plant relationships: physiological, molecular, agricultural, and environmental advances (1997-2003). Can. J. Microbiol. 50(8):521-577. |
|
|
Beck C, Oerke EC, Dehne HW (2002). Impact of strobilurins on physiology and yield formation of wheat. Mededelingen (Rijksuniversiteit te Gent. Fakulteit van de Landbouwkundige en Toegepaste Biologische Wetenschappen) 67(2):181-187. |
|
|
Bortolini CG, Silva PRF, Argenta G, Forsthofer EL (2001). Rendimento de grãos de milho cultivado após aveia-preta em resposta à adubação nitrogenada e regime hídrico. Pesqui. Agropecu. Bras. 36(9):1101-1106. |
|
|
Brannstrom C, Jepson W, Filippi AM, Redo D, Xu Z, Ganesh S (2008). Land change in the Brazilian Savannah (Cerrado), 1986-2002: comparative analysis and implications for land-use policy. Land Use Pol. 25(4):579-595. |
|
|
Buff K, Mano DMS, Langenbach T (1992). Effect of endosulfan on Azospirillum lipoferum growth, morphology, nitrogenase activity and protein binding. Appl. Environ. Microbiol. 58:3173-3176. |
|
|
Bustamante MMC, Medina E, Asner GP, Nardoto GB, Garcia-Montiel DC (2006). Nitrogen cycling in tropical and temperate savannas. Biogeochemistry 79:209-237. |
|
|
Cassán FD, Perrig D, Sgroy V, Masciarelli O, Penna C, Luna V (2009). Azospirillum brasilense Az39 and Bradyrhizobium japonicum E109, inoculated singly or in combination, promote seed germination and early seedling growth in corn (Zea mays L.) and soybean (Glycine max L.). Eur. J. Soil Biol. 45(1):28-35. |
|
|
Cassán FD, Sgroy V, Perrig D, Masciarelli O, Luna V (2008). Producción de fitohormonas por Azospirillum sp. Aspectos fisiológicos y tecnológicos de la promoción del crecimiento vegetal. In: Azospirillum sp.: cell physiology, plant interactions and agronomic research in Argentina (Eds.:Cassán FD, Salamone IG). Asociación Argentina de Microbiologia, Buenos Aires, Argentina pp. 59-84. |
|
|
Cavaglieri L, Orlando J, Etcheverry M (2009). Rhizosphere microbial community structure at different maize plant growth stages and root locations. Microbiol. Res. 164:391-399. |
|
|
CEPF (Critical Ecosystem Partnership Fund) (2015). Cerrado. View Downloaded: Mar 01, 2015. |
|
|
CFSEMG (Comissão de Fertilidade do Solo do Estado de Minas Gerais) (1999). Recomendações para o uso de corretivos e fertilizantes em Minas Gerais, 5ª aproximação. Comissão de Fertilidade do Solo do Estado de Minas Gerais: Viçosa, Brazil. 359 p. |
|
|
Chamberlain K, Patel S, Bromilow RH (1998). Uptake by roots and translocation to shoots of two morpholine fungicides in barley. Pestic. Sci. 54(1):1-7. |
|
|
Compant S, Clément C, Sessitsch A (2010). Plant growth-promoting bacteria in the rhizo- and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 42:669-678. |
|
|
CONAB (Companhia Nacional de Abastecimento) (2015a). Brazilian Crop Assessment: grains: Fourth Assessment, January/2015. CONAB, Brasília, DF, Brazil. View Downloaded: Feb 08, 2015. |
|
|
CONAB (Companhia Nacional de Abastecimento) (2015b). Séries históricas: estimativa de produtividade safra 2014/2015 Milho. CONAB, Brasília, DF, Brazil. View Downloaded: Feb 08, 2015. |
|
|
Díaz-Zorita M, Fernández-Canigia MV (2009). Field performance of a liquid formulation of Azospirillum brasilense on dryland wheat productivity. Eur. J. Soil Biol. 45(1):3-11. |
|
|
Diedhiou PM, Oerke EC, Dehne HW (2004). Effects of the strobilurin fungicides azoxystrobin and kresoxim-methyl on arbuscular mycorrhiza. J. Plant Dis. Prot. 111(6):545-556. |
|
|
Döbereiner J (1992). Fixação de nitrogênio em associação com gramíneas. In: Microbiologia do solo (Ed.: Cardoso EJBN). Sociedade Brasileira de Ciências do Solo, Campinas pp. 173-180. |
|
|
Fagan EB, Dourado Neto D, Vivian R, Franco RB, Yeda MP, Massignam LF, Oliveira RF, Martins KV (2010). Efeito da aplicação de piraclostrobina na taxa fotossintética, respiração, atividade da enzima nitrato redutase e produtividade de grãos de soja. Bragantia 69(4):771-777. |
|
|
Ferreira AS, Pires RR, Rabelo PG, Oliveira RC, Luz JMQ, Brito CH (2013). Implications of Azospirillum brasilense inoculation and nutrient addition on maize in soils of the Brazilian Cerrado under greenhouse and field conditions. Appl. Soil Ecol. 72:103-108. |
|
|
Forlani G, Mantelli M, Branzoni M, Nielsen E, Favilli F (1995). Differential sensitivity of plant-associated bacteria to sulfonylurea and imidazolinone herbicides. Plant Soil 176(2):243-253. |
|
|
Gallardo ALCF, Bond A (2011). Capturing the implications of land use change in Brazil through environmental assessment: time for a strategic approach? Environ. Impact Assess. Rev. 31(3):261-270. |
|
|
Grossmann K, Kwiatkowski J, Caspar G (1999). Regulation of phytohormone levels, leaf senescence and transpiration by the strobilurin kresoxim-methyl in wheat (Triticum aestivum L.). J. Plant Physiol. 154:805-808. |
|
|
Grossmann K, Retzlaff G (1997). Bioregulatory effects of the fungicidal strobilurin kresoxim-methyl in wheat (Triticum aestivum L.). Pestic. Sci. 50(1):11-20. |
|
|
Haridasan M (2008). Nutritional adaptations of native plants of the cerrado biome in acid soils. Braz. J. Plant Physiol. 20(3):183-195. |
|
|
Haridasan M (1994). Solos do Distrito Federal. In: Cerrado: Caracterização, ocupação e perspectivas - O caso do Distrito Federal (Ed.: M. Novaes-Pinto). Editora Universidade de Brasília/SEMATEC, Brasília pp. 322-334. |
|
|
Hartmann A, Bashan Y (2009). Ecology and application of Azospirillum and other plant growth-promoting bacteria (PGPB) – Special Issue. Eur. J. Soil Biol. 45(1):1-2. |
|
|
Hungria M, Campo RJ, Souza EM, Pedrosa FO (2010). Inoculation with selected strains of Azospirillum brasilense and A. lipoferum improves yields of maize and wheat in Brazil. Plant Soil 331:413-425. |
|
|
Hungria M (2011). Inoculação com Azospirillum brasilense: inovação em rendimento a baixo custo. Embrapa-Soja: Londrina, Brazil. 36 p. |
|
|
Jena PK, Adhya TK, Rao VR (1990). Nitrogen fixing bacterial populations as influenced by butachlor and thiobencarb in rice soils. Zentbl. Mikrobiol. 145(6):457-460. |
|
|
Kaiser WM, Brendle-Behnisch E (1995). Acid-base-modulation of nitrate reductase in leaf tissues. Planta 196(1):1-6. |
|
|
Klink CA, Machado RB (2005). Conservation of the Brazilian Cerrado. Conserv. Biol. 19(3):707-713. |
|
|
Köhle H, Gold RE, Ammermann E (1994). Biokinetic properties of BAS 490 F and some related compounds. Biochem. Soc. Trans. 22(1):65S. |
|
|
Köhle H, Grossmann K, Jabs T, Gerhard M, Kaiser W, Glaab J, Conrath U, Seehaus K, Herms S (2003). Physiological effects of the strobilurin fungicide F 500 on plants. AgroConcept, pp. 61-74. |
|
|
Langenbach T, Mano DMS, De-Souza W, Hagler AN (1991). Influence of insecticides on growth, nitrogenase activity and morphology of Azospirillum lipoferum. Ciênc. Cult. 43:207-209. |
|
|
Lopes AJ, Cox FR (1977). A survey of the fertility status of surface soils under cerrado vegetation of Brazil. Soil Sci. Am. J. 41:752-757. |
|
|
Mehnaz S, Kowalik T, Reynolds B, Lazarovits G (2010). Growth promoting effects of corn (Zea mays) bacterial isolates under greenhouse and field conditions. Soil Biol. Biochem. 42(10):1848-1856. |
|
|
Myresiotis CK, Vryzas Z, Papadopoulouâ€Mourkidou E (2014). Effect of specific plantâ€growthâ€promoting rhizobacteria (PGPR) on growth and uptake of neonicotinoid insecticide thiamethoxam in corn (Zea mays L.) seedlings. Pest manag. Sci. 71(9):1258-1266. |
|
|
Ohland RAA, Souza LCF, Hernani LC, Marchetti ME, Gonçalves MC (2005). Culturas de cobertura do solo e adubação nitrogenada no milho em plantio direto. Ciênc. Agrotec. 29(3):538-544. |
|
|
Okon Y, Labandera-González CA (1994). Agronomic applications of Azospirillum: an evaluation of 20 years worldwide field inoculation. Soil Biol. Biochem. 26:1591-1601. |
|
|
Omar MNA, Berge O, Hassanein EE, Shalan SN (1992). In vitro and in situ effects of herbicide thiobencarb on rice-Azospirillum association. Symbiosis 13:55-63. |
|
|
Pereyra MA, Ballesteros FM, Creus CM, Sueldo RJ, Barassi CA (2009). Seedlings growth promotion by Azospirillum brasilense under normal and drought conditions remains unaltered in Tebuconazole-treated wheat seeds. Eur. J. Soil Biol. 45(1):20-27. |
|
|
Rava CA (2002). Eficiência de fungicidas no controle da antracnose e mancha angular do feijoeiro comum. Summa Phytopathol. 28(1):65-69. |
|
|
Reis VM, Baldani JI, Baldani VLD, Döbereiner J (2000). Biological dinitrogen fixation in gramineae and palm trees. CRC Cric. Rev. Plant Sci. 19:227-247. |
|
|
Revellin C, Giraud JJ, Silva N, Wadoux P, Catroux G (2001). Effect of some granular insecticides currently used for the treatment of maize crops (Zea mays) on the survival of inoculated Azospirillum lipoferum. Pest Manage. Sci. 57(11):1075-1080. |
|
|
Ritchie SW, Hanway JJ, Benson GO (1992). How a Corn Plant Develops. Iowa State University Extension Department: Ames, Iowa, Story County, USA. 26 p. |
|
|
Rivarola V, Fabra A, Mori G, Balegno H (1992). In vitro protein synthesis is affected by the herbicide 2,4-dichlorophenoxyacetic acid in Azospirillum brasilense. Toxicology 73:71-79. |
|
|
Ruske RE, Gooding MJ, Jones SA (2003). The effects of triazole and strobilurin fungicide programmes on nitrogen uptake, partitioning, remobilization and grain N accumulation in winter wheat cultivars. J. Agric. Sci. 140(4):395-407. |
|
|
Sá SA, Palmer C, Di Falco S (2013). Dynamics of indirect land-use change: empirical evidence from Brazil. J. Environ. Econ. Manag. 65(3):377-393. |
|
|
Salamone IEG, Döbereiner J (1996). Maize genotype effects on the response to Azospirillum inoculation. Biol. Fertil. Soils 21(3):193-196. |
|
|
Salamone IEG, Döbereiner J, Urquiaga S, Boddey RM (1996). Biological nitrogen fixation in Azospirillum strain-maize genotype association as evaluated by 15N isotope dilution technique. Biol. Fertil. Soils 23(3):249-256. |
|
|
Salmeron V, Martinez-Toledo MV, Gonzalez-Lopez J (1991). Effects of alachlor and metolachlor on the biological activity of Azospirillum brasilense grown in chemically defined and dialyzed-soil media. Environ. Toxicol. Chem. 10(4):493-499. |
|
|
Sánchez CE, Rodelas B, Martinez-Toledo MV, Salmeron V, Gonzalez-Lopez J (1994). Diflubenzuron and the biological activity of Azospirillum brasilense. Toxicol. Environ. Chem. 42(3-4):241-247. |
|
|
Scarano FR, Silva JMC, Guimarães AL, Raik D, Bolt F (2012). Brazil on the spot: Rio+20, sustainability and a role for science. Braz. J. Bot. 35(2):233-239. |
|
|
Sicbaldi F, Sacchi GA, Trevisan M, Del Re AAM (1997). Root uptake and xylem translocation of pesticides from different chemical classes. Pestic. Sci. 50(2):111-119. |
|
|
Silva EC, Buzetti S, Guimarães GL, Lazarini E, Sá ME (2005). Doses e épocas de aplicação de nitrogênio na cultura do milho em plantio direto sobre Latossolo Vermelho. Rev. Bras. Ciênc. Solo 29:353-362. |
|
|
Silva JF, Fari-as MR, Felfili JM, Klink CA (2006). Spatial heterogeneity, land use and conservation in the Cerrado region of Brazil. J. Biogeogr. 33(3):536-548. |
|
|
Souza JA (2006). Manejo da fertilidade de solo para a cultura do milho. Inf. Agropecuário 27(233):26-37. |
|
|
Trivedi M, Pinto LFG, Hall A, da Motta RS, Filho LM, Strassburg B, Mitchell A, Ortiz R, Costa D, Oakes N, Ometto J (2012). Think PINC: Securing Brazil's Food, Water and Energy with Proactive Investment in Natural Capital. Global Canopy Programme: Oxford, Oxfordshire, England, UK. 21 pp. |
|
|
Valipour M (2012). Critical areas of Iran for agriculture water management according to the annual rainfall. Eur. J. Sci. Res. 84(4):600-608. |
|
|
Valipour M (2013) Need to update of irrigation and water resources information according to the progresses of agricultural knowledge. Agrotechnology S10:e001. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


