Full Length Research Paper

ABSTRACT
This study aimed to phenotypically characterize indigenous Tswana sheep under its natural environment and develop prediction equations for body weight using linear body measurements in Kgatleng, Kweneng, Southern and South-East districts of Botswana. Multistage purposive sampling was used for selection of districts. Data on qualitative characters and quantitative measurements were made on 665 sheep stratified by dentition into four age categories (0PPI, 1PPI, 2PPI and ≥3PPI). Both qualitative and quantitative data were analyzed using Statistical Analysis System. Most Tswana sheep were characterized by plain coat color pattern with white dominating and plain white colors, short-fat tails with a straight tip, horizontal ears, no horns and wattles. District, sex and age had a significant influence on body weight and most linear body measurements. The highest correlation between body weight and heart girth for both sexes indicate that heart girth explained more variation than other measurements thus is the best variable for predicting body weight in both sexes. The best predicted body weight model for males is y=-64.15+1.28x and y=-53.47+1.14x for females where x and y are heart girth and body weight, respectively. This study reveals existence of diversity of sheep genetic resources across districts, indicating their potential response to selection.
Key words: Body weight, characterization, morphometric traits, Tswana sheep.
INTRODUCTION
Indigenous sheep are widely distributed in the tropics and subtropics due to their unique adaptive features that enable them to fit in a wide variety of environments (Berihulay et al., 2019). In Botswana, the indigenous sheep constitute about 195 000 of the 300 000 national flock (Botswana Statistics, 2016) and is adapted to different geographical regions of the country. It contributes significantly to the livelihoods of resource poor farmers by providing meat and milk as a source of nutrition to the household, and income which is used for a wide range of economic activities. Indigenous Tswana Sheep retain certain adaptive features such as drought, heat and disease tolerance (Nsoso et al., 2004b). Very little has been done towards improvement and characterization of Tswana Sheep. The major threat facing indigenous Tswana sheep genetic resources is uncontrolled breeding with exotic breeds or breed replacement with exotic breeds carried out in an endeavor to improve the breed to meet current market demands for more mutton.
It is therefore very important to develop strategies for sustainable utilization of Tswana sheep genetic resources. The prerequisite to developing these strategies is the characterization of the genetic resource under its natural environment (Msanga et al., 2012; Monau et al., 2018). Phenotypic characterization of local genetic resources is essential for conservation, breed inventory and monitoring, policy formulation and design of breeding programmes (Baker and Gray, 2004). Phenotypic characterization of indigenous Tswana sheep was undertaken more than a decade ago and may not reflect the current situation due to changes in production systems and within population’s changes resulting from evolutionary forces (Sölkner et al., 1998). Morphological and productive aspects in a population evolve over time as a result of natural and artificial selection, mutation, migration, and random genetic drift (Song et al., 2006). Differences in environment and differences in climatic factors such as rainfall and temperatures also influence adaptive features of local populations which might result in phenotypic differences in the general population (Schierenbeck, 2017). There is therefore need to carry out routine inventories and monitoring of the indigenous Tswana sheep genetic resource. The aim of this was therefore to phenotypically characterize indigenous Tswana sheep under its native environment and to develop a prediction equation for body weight by using linear body measurements in the Kgatleng, Kweneng, Southern and South-East districts of Botswana.
MATERIALS AND METHODS
Description of the study area
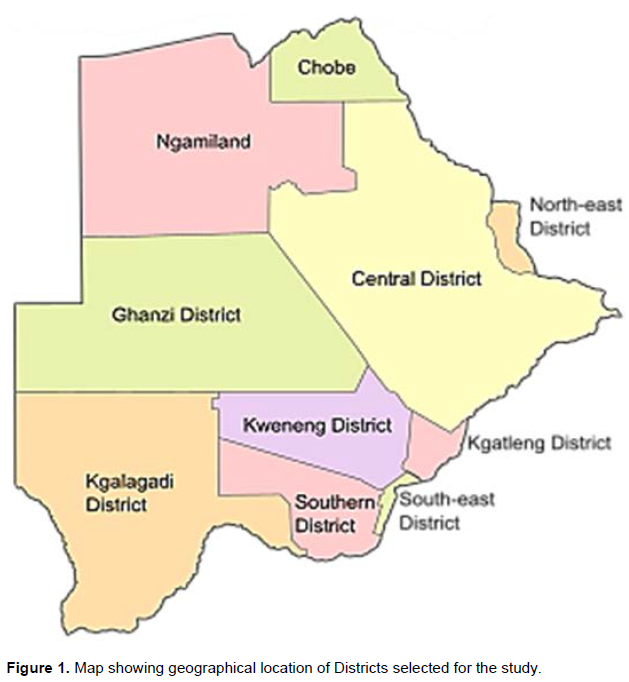
The study was conducted in Kgatleng (24°15'S 26°30’E with total land area of 7,960 km2) Kweneng (24°00'S 25°00'E with total land area of 35,890 km2), South East (25°00'S 25°45'E with total land area of 1,780 km2) and Southern (25°00'S 25°00'E with total land area of 28, 470 km2) districts of Botswana from November 2020 to January 2021 (Figure 1). The climate in the four districts is mainly semi-arid with high temperatures occurring from October to April and low temperatures occurring from around May to August. Rainfall is low, unreliable highly variable from one year to the next and not evenly distributed. The vegetation type in all districts is savannah with tall grasses, bushes and trees (Makhabu et al., 2002; Nsoso et al., 2004b).
Sampling method
A multi-stage purposive sampling technique was employed for selection of districts for the study. In the first stage, discussions were held with district agricultural officers of the Department of veterinary services to know the distribution of indigenous Tswana sheep population in each study district. The four districts (Kgatleng, Kweneng, South-East and Southern) were purposely selected for morphological characterisation based on the distribution of indigenous Tswana sheep. Random sampling was used to select villages within districts and farms/households within villages. Six villages per districts were randomly selected and four to five farms per village were also randomly selected for the study. One to six unrelated animals were sampled per household.
Data collection procedures
A total of six hundred and sixty five sheep were used for phenotypic characterisation. Morphological features were recorded for every animal sampled following breed morphological characteristics descriptor guidance list of FAO (2012). Visual observations of qualitative traits such as coat colour, presence of wattles and presence of horns were recorded. Quantitative traits [heart girth (HG), body length (BL), wither height (WH), rump width (RW), ear length (EL), tail length (TL), tail circumference (TC), head length (HL), head width (HW), shoulder width (SW), cannon bone length (CBL), cannon bone circumference (CBC), neck length (NL), rump length (RL), rump height (RH) and scrotal circumference (SC) (in males)] were measured using a flexible tailor’s measuring tape calibrated in centimetres (cm). All measurements were taken early in the morning to avoid the effect of feeding and watering on the animal’s size. The animals were restrained in an upright unforced plane position during data collection. All measurements were taken by the same personnel in all the districts for consistency.
Each experimental animal was identified by its sex, age and sampling site (district). Sex was characterized as females, rams and castrates. The age of each animal was estimated based on farmers’ information and dentition following the procedure described for African sheep by Wilson and Durkin (1984). Sheep were classified into four age groups: no pair of permanent incisors (0PPI), one pair of permanent incisors (1PPI), two pairs of permanent incisors (2PPI), and three and above pairs of permanent incisors (≥3PPI) to represent the ages of 6-11, 12–24, 25–36 and above 36 months, respectively.
The step-wise regression procedures of SAS Statistical Analysis System (SAS, release 9.1 2003) were used to develop prediction equations used for body weight (BW) in Tswana sheep. The prediction of body weight for Tswana sheep was based on the regression equations y= -64.15+1.28x for males and y= -53.47+1.14x for females where, x and y are heart girth and body weight, respectively.
Statistical analysis
Qualitative data from individual observation were analysed following the frequency procedures of Statistical Analysis System (SAS release 9.1 2003). The General Linear Model (GLM) procedures of Statistical Analysis System (SAS, release 9.1 2003) were used to estimate least squares means and standard errors of quantitative linear body measurements. Sex, district and age group of the sheep were fitted as fixed effects, while body weight and linear body measurements (except scrotal circumference) were fitted as dependent variables. Scrotal circumference was analysed by fitting age and district as fixed factors for intact males. Least square means and their corresponding standard errors were calculated for fixed effects of sex, age, district and the age by sex interaction for each body trait. Model used for the least square mean analysis of body weight and other linear body measurements in females and males except scrotal circumference was:
Yijk =µ + Ai + Dj + Sk+ (AxD)ij + (AxS)ik+ (DxS)jk + eijk
Where: Yijk = Observed body weight or linear measurements
µ = Overall mean
Ai = the fixed effect of ith age groups (i = 0PPI, 1PPI, 2PPI and ≥ 3PPI)
Dj = the fixed effect of jth district (j= Kgatleng, Kweneng, South-East and Southern districts)
Sk = the fixed effect of the kth sex (k= male, female)
(AxD)ij= the effect of the interaction of i of age group with j of district
(AxS)ik= the effect of the interaction of i of age group with k of sex
(DxS)jk= the effect of the interaction of j of district with k of sex
eijk= random residual error
Model used for the least square mean analysis in males for scrotal circumference was:
Yijk =µ + Ai + Dj + (AxD)ij + eij
Where: Yijk = Scrotal circumference
µ = Overall mean
Ai = the fixed effect of ith age groups (i = 0PPI, 1PPI, 2PPI and ≥ 3PPI)
Dj = the fixed effect of jth district (j= Kgatleng, Kweneng, South-East and Southern districts)
(AxD)ij= the fixed effect of the interaction of ith age group with jth of district
eijk= random residual error
Correlations and regression
Pearson’s correlation coefficients of indigenous Tswana sheep were estimated between body weight and other linear body measurements (LBMs) within each sex using the procedure correlation (PROC CORR) of Statistical Analysis System (SAS release 9.1 2003) to describe the strength and direction of relationships between the response variable (live body weight) and explanatory variables (LBMs). Body weight and other LBMs (BL, HG, WH, RH, SW, EL, RL, CBC, CBL, NL, RW, HW, HL, TL, TC and SC) were included for males whereas SC was excluded when calculating correlations coefficients for female sheep. Based on the correlations of body weight with other LBMs, a stepwise regression procedure (PROC REG) of Statistical Analysis System (SAS release 9.1 2003) was then used to regress body weight for each sex in order to determine the best-fit regression equation for the prediction of body weight using LBMs. The best-fit models were selected based on the coefficient of determination (R2) and the simplicity of measurements of the LBMs under field conditions. The following models were used for the analysis of multiple linear regressions.
For males:
Υj =β0 + β1X1 + β2X2 + β3X3 + β4X4 + β5X5 + β6X6 + β7X7+ β8X8 + β9X9+ β10 X10 + β11X11 + β12X12 + β13X13 + β14X14 + β15X15 + β16X16 + ej
Where:
Yj =the response variable (body weight)
β0 = the intercept
X1, X2, X3, X4, X5, X6, X7, X8, X9, X10, X11, X12, X13, X14, X15 and X16 are the explanatory variables BL, HG, WH, RH, SW, EL, RL, CBC, CBL, NL, RW, HW, HL, TL, TC and SC, respectively.
β1, β2…β16 are the regression coefficients of the variables X1, X2…X16.
ej = the residual random error.
For female:
Υj =β0 + β1X1 + β2X2 + β3X3 + β4X4 + β5X5 + β6X6 + β7X7+ β8X8 + β9X9+ β10 X10 + β11X11 + β12X12 + β13X13 + β14X14 + β15X15 + ej
Where:
Yj =the response variable (body weight)
β0 = the intercept
X1, X2, X3, X4, X5, X6, X7, X8, X9, X10, X11, X12, X13, X14 and X15 are the explanatory variables BL, HG, WH, RH, SW, EL, RL, CBC, CBL, NL, RW, HW, HL, TL, and TC, respectively.
β1, β2…β16 are the regression coefficients of the variables X1, X2…X15.
ej = the residual random error.
RESULTS AND DISCUSSION
Table 1 shows some qualitative characters of both male and female indigenous Tswana sheep raised in Kgatleng, Kweneng, South-East and Southern districts of Botswana. There was variation on coloration patterns amongst the sheep populations with predominantly plain coat colour patterns (81.25%) across districts, followed by patchy and spots of different colours (Figure 2). The results are consistent with the findings of Asaminew et al. (2016), Edea et al. (2010) and Tibbo and Ginbar (2004) for Bonga, Horro and Woliata sheep types of Ethiopia. The higher proportion of animals with white dominated coat colour and plain white coat colour could be a reflection of natural selection for animals manifesting white colour to withstand the hot environment of Botswana. This observation is different in other part of the African continent such as Ethiopia where Hailemariam et al. (2018) reported the dominance of black coat colour for Gamogofa sheep which helps in absorption of solar radiation to maintain an optimum body temperature in the cold Gamogofa zone.
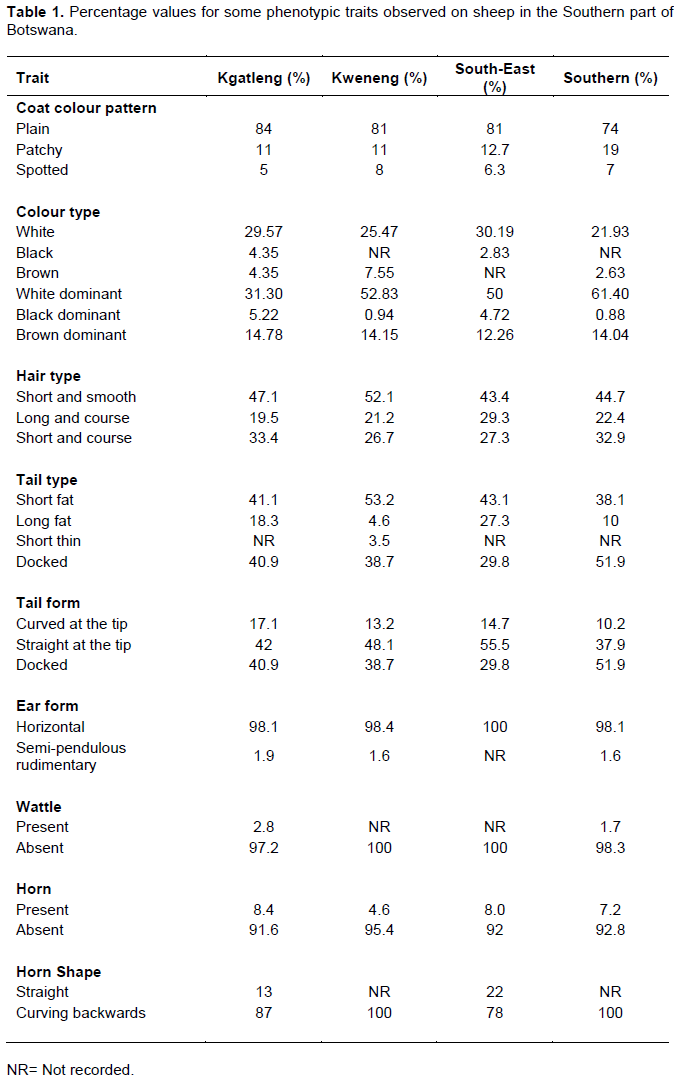

Most of sheep across the districts had a characteristic short fat tail with a straight tip pointing downwards (Figure 2). The fat tail is an adaptive attribute that serves as an energy reserve to enable indigenous Tswana sheep to adapt and survive feed fluctuation periods throughout the year (Ermias et al., 2002). This characteristic has also been reported in some South African sheep breeds including the South African Namaqua Afrikaner, the Zulu and the Pedi sheep (Soma et al., 2012). To the contrary, Getachew et al. (2010) reported short fat tails curved upwards in Menz sheep of Ethiopia. Contrary to the current study, Giwaz et al. (2007) and Edea et al. (2010) who reported a long fat tail characteristic in Adilo and Bonga sheep of Ethiopia. Differences between tail types is associated with genetic variations of sheep types (Ermias et al., 2002). The predominant ear form or orientation observed in indigenous Tswana sheep (overall 98.65%) was horizontal ear orientation. Almost all sheep across districts (97.2% for Kgatleng, 100% for Kweneng, 100% for South-East and 98.3% for Southern district) had no wattles. The findings are similar to Gamogofa sheep of Ethiopia (Hailemariam et al., 2018) and a lower proportion of Tswana sheep with wattles in Kgatleng and South-East district is consistent with Melesse et al. (2013) and Tibbo and Ginbar (2004) for Bonga sheep of Ethiopia. A comparatively high proportion of indigenous Tswana sheep across districts did not have horns and a high proportion of sheep with horns had their horns curving backwards as opposed to straight (Table 1).
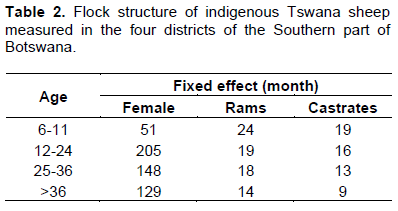
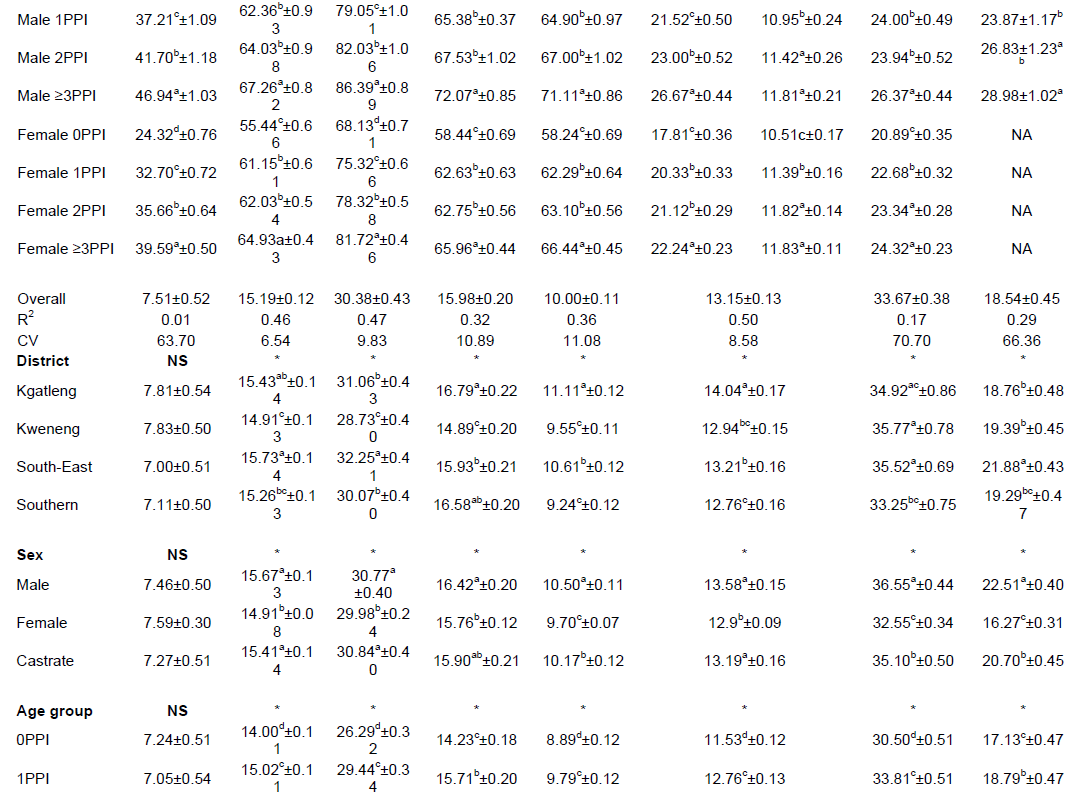
Table 2 shows the flock structure of indigenous Tswana sheep in Kgatleng, Kweneng, South-East and Southern districts of Botswana. Generally, the proportion of female sheep increased with age, hence females out-numbered males (rams and castrates) across all age groups except for the 6-11 month age group (Table 2). This is because selection of breeding rams is mostly done after 12 months of age after the rams had reached sexual maturity. Rams that have not been selected for breeding are mostly castrated or culled while almost all females are retained for breeding purposes. A few males are selected for breeding purposes based on favourable desired traits while culls are sold for meat production or to other farmers who further do some selection to suit their farming needs (Nsoso et al., 2004b). Again, the traditional meat market also requires animals of an older age group (>12 months) to attract a favourable price (Nsoso and Madimabe, 1999). Similar findings have been previously reported by Katongole et al. (1996) on goats and Nsoso et al. (2004b) on goats and sheep.
Body weight and linear measurements
Age effect
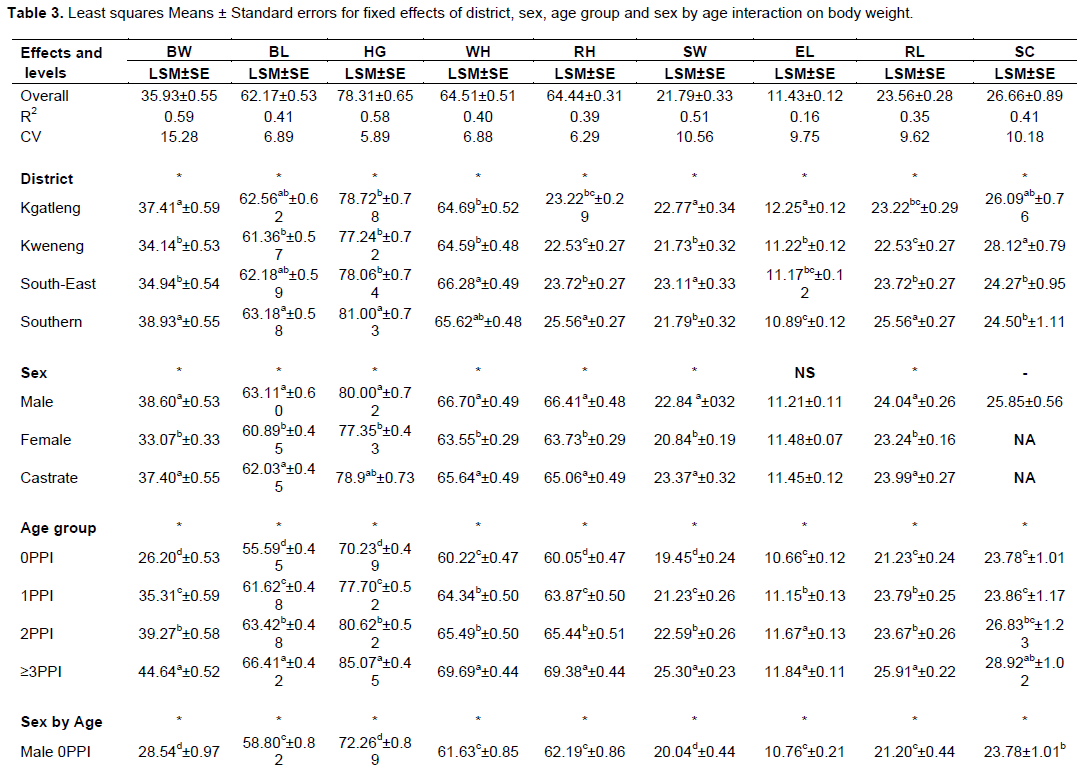
In this study, body weight and most quantitative linear body measurements were significantly (P<0.05) affected by the age group except cannon bone circumference (Table 3). All body measurements increased as the animal advanced in age from the 0PPI age group to the ≥3PPI age group. This is expected and has been echoed by several scholars who observed that age had a significant effect on body weight and other linear body measurements and that the size and shape of sheep increased as the animal advanced with age, until optimum growth or maturity (Fasae et al., 2006; Yoseph, 2007; Tesfaye, 2008; Asaminew et al., 2016).
Sex effect
The sex of the animal had a significant effect on body weight and most linear body measurements except ear length, neck length and cannon bone circumference across the four districts (Table 3). Similar findings have been reported by Kunene et al. (2007) in Zulu sheep of South Africa and Shibabaw (2012) in Hararghe highland sheep of Ethiopia. To the contrary, Asefa et al. (2017) reported a non-significant effect of sex on body and some linear body measurements in Bale Zone sheep of Ethiopia.
District effect
The least square means and standard errors for the effect of district on live body weight and other linear body measurements of indigenous Tswana sheep are presented in Table 3. The district effect was significant (P<0.05) for body weight and most linear body measurements except cannon bone circumference. The significant district effect on body weight and linear body measurements is consistent with Asefa et al. (2017), Alemayehu (2011) and Kunene et al. (2007). Southern district sheep were the heaviest (38.93±0.55 kg) and Kweneng sheep were the lightest (34.14±0.53 kg). Generally, Tswana sheep were comparable in body weight to Zulu sheep (39.76 to 40.26 kg) (Kunene et al., 2007) and heavier than several indigenous sheep of Ethiopia (Asaminew et al., 2016; Michael et al., 2016; Mohammed et al., 2017). Tswana sheep are however lighter than Balami and Uda sheep types from South, Middle belt and North West districts of Nigeria (Agaviezor et al., 2012).
Southern district sheep had significantly (P<0.05) higher heart girth than sheep in Kgatleng, Kweneng and South-East districts of Botswana (Table 3). The difference in heart girth between sheep in different regions has also been reported by Asaminew et al. (2016) who found higher heart girth in Soddo Zuria and Damote Gale sheep than Damote Sore sheep of Ethiopia. Generally Tswana sheep had similar heart girth with Uda sheep of Nigeria (Agaviezor et al., 2012) and Hulet eju sheep in Ethiopia (Michael et al., 2016). The heart girth of Tswana sheep in the Southern district was higher than the heart girth of sheep in Wogide, Borena and Legambo districts of Ethiopia (Mohammed et al., 2017) and Sinan and Hulet eju sheep of Ethiopia (Michael et al., 2016). Southern district Tswana sheep had the longest body length than sheep from other districts while Kweneng district sheep were the shortest. Body length of Tswana sheep in Kgatleng, Kweneng and Southern sheep were similar to those of Gozamen, Sinan and Hulet-eju sheep of Ethiopia (Michael et al., 2016) and longer than that of Borena and Legambo sheep of Ethiopia (Mohammed et al., 2017). Generally, Tswana sheep across the districts were shorter than Soddo Zuria and Damote Gale of Southern Ethiopia (Asaminew et al., 2016).
South-East and Southern district Tswana sheep had similar wither height and were significantly (P<0.05) taller than Kgatleng and Kweneng sheep which also had similar wither height. South-East and Southern district Tswana sheep had similar wither height to Borena sheep of Ethiopia (Mohammed et al., 2017) and Agarfa sheep of Ethiopia (Asefa et al., 2017). Generally, Tswana sheep are taller than Metta, Gorogutu and Deder sheep of Ethiopia (Shibabaw, 2012) and Damote Sore sheep of Ethiopia (Asaminew et al., 2016) and shorter than Gozamen, Sinan and Hulet eju sheep of Ethiopia (Michael et al., 2016). The differences in body weight and other linear body measurements could be as a result of differences breed structure emanating from in the influences of evolutionary forces of the world. Other differences in body weight and other linear body measurements could be due to nutritional and management practices between districts. In females, the pregnancy status of the animal could be another reason for variations in body weight and other linear body measurements especially body length (Kunene et al., 2007). The wide variations in body weight and other linear body measurements of sheep between districts indicate healthy diversity which could be exploited in genetic improvement programmes of indigenous Tswana sheep (Berhanu and Haile, 2009).
Kgatleng and Kweneng rams had similar scrotal circumference that were significantly higher than scrotal circumference of sheep in South-East and Southern districts. The significant influence of district on scrotal circumference of Tswana sheep found in the current study is contrary to Mohammed et al. (2017) for Wogide, Borena and Legambo rams of Ethiopia and Michael et al. (2016) for Gozamen, Sinan and Hulet eju rams of Ethiopia who reported a non-significant influence of district on ram scrotal circumference. Scrotal circumference of Tswana sheep in Southern and South-East districts of Botswana is similar to that of Wogide and Legambo rams (Mohammed et al., 2017) and Zulu rams (Kunene et al., 2007). Generally, the scrotal circumference of Tswana rams was higher than that of Borena rams of Ethiopia (Mohammed et al., 2017) and Soddo Zuria, Damote Gale and Damote Sore rams of Ethiopia (Asaminew et al., 2016). Differences in scrotal circumference between breeds and districts might be due to the fact that testicular size varies with breed, age of an animal and season/time of the year (Söderquist and Hulten, 2006). Kunene et al. (2007) reported larger scrotal circumference in autumn and in summer compared to winter and spring in Zulu rams, probably because of fluctuations in fodder quality and quantity with changes in times of seasons of the year. Furthermore, Dana et al. (2000) reported reduced scrotal circumference by up to 10% in Ethiopian highland sheep fed low quality diet than those fed good quality diet. Low quality diet causes loss of fat from scrotal tissue of rams resulting in reduced testicular size (Coulter and Kozub, 1984).
Age by sex interaction
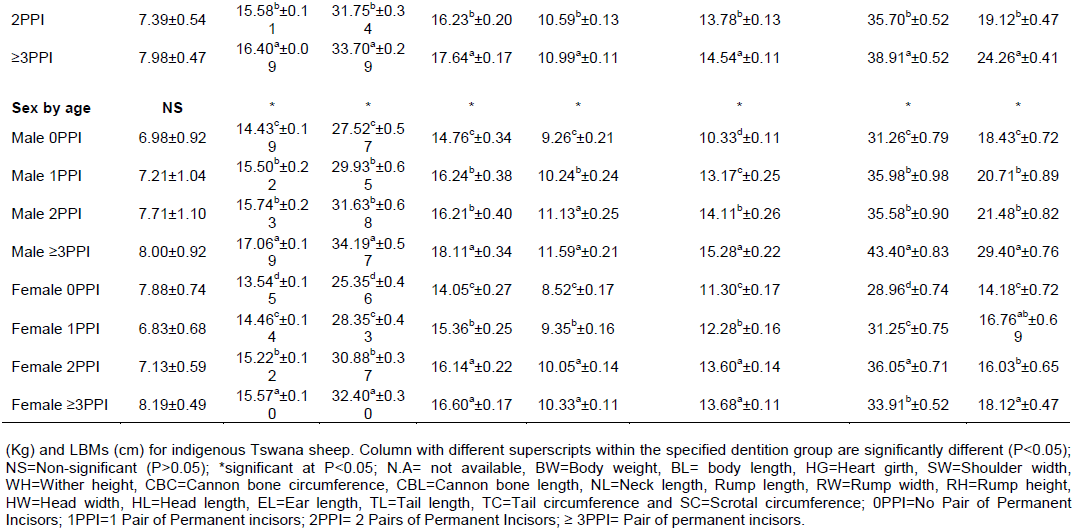
The least square means and standard errors for the effect of sex, age group and their interaction on body weight and other LBMs of indigenous Tswana sheep are presented in Table 3. The interaction of sex and age group was significant (P<0.05) for body weight and most LBMs (BL, HL, HW, SW, CBL, HG, WH, RL, RH, RW, TL and TC). However, the interaction effect was not significant (P>0.05) for cannon bone circumference and ear length, implying that these parameters were not affected by the sex X age group interaction in the current study. In consonance with Tassew (2012) the sex X age group interaction was significant (P<0.05) for most LBMs except ear length in Habru and Gubalafto sheep. Contrary to the current findings, Michael et al. (2016) and Alemayehu (2011) reported a significant sex by age group interaction (P<0.01) only for body weight in Dawro and Konta Special Woreda zones of Ethiopia. Kunene et al. (2007) also reported a non-significant sex by age interaction for LBMs between Zulu rams and ewes lambs of South Africa at their milk stage.
The sex by age interaction found in the current study revealed that the differences in live body weight between males and females increased with the age of the animals from 4.47 to 5.20 kg and 7.90 kg at one, two and three pairs of permanent teeth, respectively (Table 3). Body weights obtained at 0PPI, 1PPI and ≥3PPI age groups in the current study were slightly higher than those previously reported by Nsoso et al. (2004b). The 2PPI age group in the current study had lower body weights than those reported by Nsoso et al. (2004b). The discrepancies might be mainly due to differences in management and evolving production systems and changing breeding goals of Tswana sheep farmers.
Generally, males were heavier and had higher LBMs than females across all age groups except ear length at 2PPI and ≥3PPI. This is consistent with Getachew et al. (2010) for Menz and Afar sheep and Tibbo et al. (2004) for Menz and Horro sheep in Ethiopia. The superiority of males over females in body weight and other LBMs might be attributed to differences in hormonal profiles between the sexes with males having hormones that promote rapid weight gain and muscularity than females, consequently resulting in superior body weight and higher LBMs in males than females (Gebreyowhens and Tasfay, 2016). In females, estrogen inhibits growth of long bones of the body resulting in slower growth rate and the reaching of puberty at a relatively smaller body size (Sowande and Sobola, 2007).
Correlation between body weight and other LBMs
Animal live body weight, linear body measurements, their interrelationships and correlations are very important in determining genetic potential for co-current improvement of traits in genetic improvement programs. The phenotypic correlation coefficients between live body weight and linear body measurements are presented in Table 4. In males, significant (P<0.05), positive and strong correlations were found between body weight and heart girth (r=0.97), followed distantly by rump height (r=0.79) and wither height (r=0.76). These linear body measurements were highly affected by the changes in body weight, hence important in predicting body weight of indigenous Tswana sheep males. Body length (r=0.67), shoulder width (r=0.67), rump length (r=0.66), cannon bone circumference (r=0.69), cannon bone length (r=0.72), neck length (r=0.69), rump width (r=0.72), head width (r=0.60), head length (r=70), tail length (r=55) and tail circumference (r=0.50) showed significant (P<0.05) moderate and positive correlations with body weight. Ear length showed a significant (P < 0.05) low and positive correlation while scrotal circumference did not show any significant correlation with body weight.
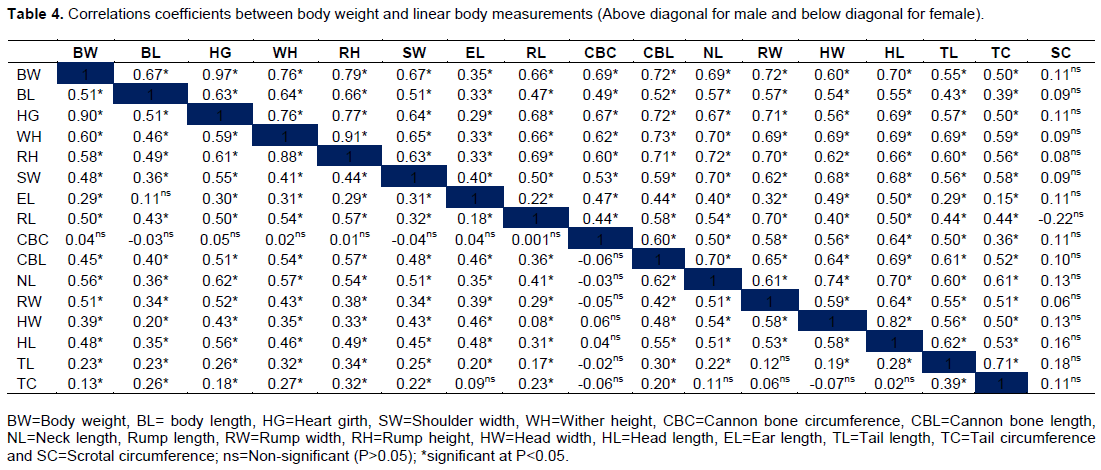
Likewise, in females, heart girth (r=0.90) showed the strongest significant (P<0.05) and positive correlation with body weight. Most linear body measurements, body length (r=0.51), wither height (r=0.60), rump height (r=0.58), shoulder width (r=0.48), rump length (r=0.50), neck length (r=0.56), rump width (r=0.51) and head length (r=0.48) had significant (P<0.05), moderate and positive correlations with body weight. Ear length (r=0.29), cannon bone length (r=0.45), head width (r=0.39), tail length (r=0.23) and tail circumference (r=0.13) showed significant low correlation with body weight whereas cannon bone circumference had no significant (P>0.05) correlation with body weight. The highest association of heart girth with body weight than other linear body measurements is consistent with Afolayan et al. (2006) and Asaminew et al. (2016).
Generally, higher correlations coefficients between body weight and other linear body measurements were found in males than females. Selection for body weight will thus result in highest co-current improvements of linear body measurements in males than females. Traits that will benefit more from selecting for higher body weight in both male and female Tswana sheep include heart girth, rump height and wither height. Asefa et al. (2017) and Asaminew et al. (2016) similarly reported heart girth as one of the traits highly correlated to body weight and will thus be significantly improved if selection is based on body weight.
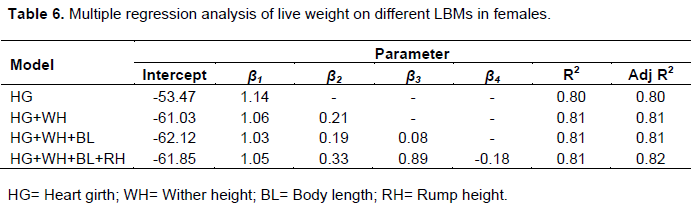
Prediction of live body weight from LBMs
Body weight is an important growth indicator and trait of economic importance that influence management interventions (drug doses) and determines the final market value of an animal (Otoikhian et al., 2008). The accuracy of functions used to predict live body weight from linear body measurements is of paramount importance in livestock production enterprises (Mohammed et al., 2017). For this study, in order to develop the prediction equation, only four linear body measurements (HG, WH, SW and RL) were selected in the prediction equation for rams (Table 5) and four linear body measurements (HG, WH, BL and RH) were selected in the prediction equation for ewes (Table 6). After comparing all coefficient values for all the relationships between body weight and other LBMs in males and females, heart girth showed the highest association with body weight in both males (0.97) and females (0.90), and thus was selected to predict body weight in each sex. This might be true because heart girth is made up of muscles, some fat and bone structure which are the main constituents of live body weight of an animal (Okpeku et al., 2011). Heart girth is deemed an easy variable to measure and is amongst the least affected by the posture of the animal (Mohammed et al., 2017). Therefore, low errors are incurred by individuals taking heart girth measurements under field conditions compared to other LBMs.
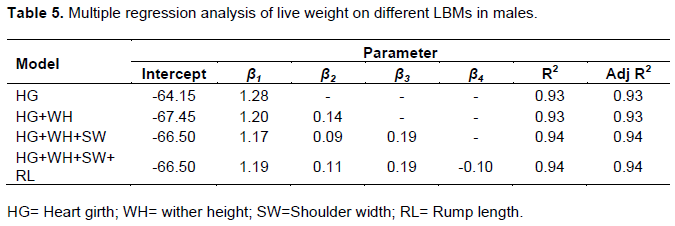
The prediction of body weight in males can be based on the prediction equation y= -64.15+1.28x and for females y= -53.47+1.14x where, x and y are heart girth and body weight respectively. The fitted prediction models were selected based on higher of coefficient of determination (R2) and smaller coefficient of variation (CV %). The R2 is a proportion of the total variability explained by the predicted model. Using heart girth in the model gave R2 values of 93% in males and 80% in females meaning that heart girth accounted for large proportions of changes in body weight in males and females, respectively. Although there is a slight increase in adjusted R2 values whenever a new variable is added to the model (Tables 5 and 6), using heart girth alone to predict body weight might be sufficient and preferable to combinations with other LBMs due to simplicity. This could prove very useful particularly under field conditions where animal restraint might be difficult during measurements. Several authors have reported that heart girth can be a sole predictor of live body weight based on its high correlation coefficients with body weight (Asefa et al., 2017; Asaminew et al., 2016).
CONCLUSIONS
The most dominant coat color patterns on indigenous Tswana sheep were plain. The white dominant and plain white was the most dominant coat colors in Tswana sheep. Most Tswana sheep were characterized by short fat tails with a straight tip pointing downwards at the end. The predominant ear orientation in Tswana sheep was horizontal. Most Tswana sheep did not have horns and wattles. The study revealed variability in LBMs between sheep in different districts of Botswana. Tswana sheep in the Southern district of Botswana displayed superiority in body weight, body length and heart girth over sheep in other districts. Generally, higher correlation coefficients between body weight and LBMs were observed between in males than females. Heart girth accounted for most of the variability in body weight than other LBMs in both males and females and was thus used as the sole predictor of body weight. The prediction equation for body weight in Tswana sheep males was y= -64.15+1.28x and for females was y= -53.47+1.14x where, x and y are heart girth and body weight, respectively.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors appreciate Botswana University of Agriculture and Natural Resources for funding this study. They are also grateful to Chandapiwa Moses, Malebogo Radikara and Josephine Motseothata who provided assistance in animal handling during data collection. They appreciate the meticulous work by the government extension staff in the districts in organizing and liaising with farmers for data collection, and are forever indebted to the smallholder farmers who allowed their animals to be used for body measurements.
REFERENCES
|
Afolayan RA, Adeyinka IA, Lakpinin CAM (2006). The estimation of live weight from body measurements in Yankasa sheep. Czech Journal of Animal Science 51(8):343-348. |
|
|
Agaviezor BO, Peters SO, Adefenwa MA, Yakubu A, Adebambo OA, Ozoje MO, Ikeobi CON, Wheto M, Ajayi OO, Amusan SA, Ekundayo OJ, Sanni TM, Okpeku M, Onasanya GO, De Donato M, Ilori BM, Kizilkaya K, Imumorin IG (2012). Morphological and microsatellite DNA diversity of Nigerian indigenous sheep. Journal of Animal Science and Biotechnology 3(1):1-16. |
|
|
Alemayehu A (2011). Phenotypic characterization of indigenous sheep types of Dawuro zone and Konta special woreda of SNNPR, Ethiopia. M.Sc. Thesis presented to the School of Graduate Studies of Haramaya University, Dire Dawa, Ethiopia. |
|
|
Asaminew M, Kebede K, Mekasha Y (2016). On farm phenotypic characterization of indigenous sheep types in Woliata Zone, Southern Ethiopia. Journal of Biology, Agriculture and Healthcare 6(11):114-124. |
|
|
Asefa B, Abate T, Adunga E (2017). Phenotypic characterization of indigenous sheep types in Bale Zone, Oromia Regional State, Ethiopia. Journal of Veterinary Science and Technology 8(4):1-7. |
|
|
Baker RL, Gray GD (2004). Appropriate breeds and breeding schemes for sheep and goats in the tropics. In. Sani RA, Gray GD, Baker RL (Eds.), Worm Control for Small Ruminants in Tropical Asia. Canberra: ACIAR Monograph Press. pp. 63-96. |
|
|
Berhanu B, Haile A (2009). Reproductive performance of traditionally managed sheep in the South Western part of Ethiopia. Livestock Research for Rural Development 21:9. |
|
|
Berihulay H, Abied A, He X, Jiang L, Ma Y (2019). Adaptation Mechanisms of Small Ruminants to Environmental Heat Stress-A Review. Animals 75(9):1-9. |
|
|
Botswana Statistics (2016). Agricultural census stats brief of 2015. Gaborone, Botswana. View. |
|
|
Coulter GH, Kozub GC (1984). Testicular development, epididymal sperm reserves and seminal quality in 2-year-old Hereford and Angus bulls: Effect of two levels dietary energy. Journal of Animal Science 59(2):432-440. |
|
|
Dana N, Shenkoru T, Tegegne A (2000). Growth rates and testicular characteristics of Ethiopian highland sheep offered chickpea haulm supplemented with incremental levels of Leucaena leucocephala leaf hay. Livestock Production Science 65(3):209-217. |
|
|
Edea Z, Haile A, Tibbo M, Sharma AK, Assefa D, Sölkner J, Wurzinger M (2010). Morphological characterization of Bonga and Horro indigenous sheep breeds under smallholder conditions in Ethiopia. Ethiopian Journal of Animal Production 9(1):117-133. |
|
|
Ermias E, Yami A, Rege JEO (2002). Fat deposition in tropical sheep as adaptive attribute to periodic feed fluctuation. Journal of Animal Breeding and Genetics 119(4):235-246 |
|
|
Fasae OA, Chineke AC, Alokan JA (2006). Relationship between some Physical Parameters of Grazing Yankasa Ewes in the Humid Zone of Nigeria. Department of Animal Production and Health. University of Agriculturehttps://www.redalyc.org/articulo.oa?id=49520807 |
|
|
Food and Agriculture Organization (FAO) (2012). Phenotypic characterization of animal genetic resources. FAO Animal Production and Health Guidelines No.11. FAO, Rome, Italy. |
|
|
Gebreyowhens W, Tesfay Y (2016). Morphological Characterization of Indigenous Highland Sheep Population of Tigray, Northern Ethiopia. Journal of Natural Sciences Research 6(1):96-104. |
|
|
Getachew T, Haile A, Tibbo M, Sharma A, Kifle A, Terefe E, Wurzinger M, Sölkner J (2010). Morphological characters and body weight of Menz and Afar sheep within their production system. Ethiopian Journal of Animal Production 9(1):99-115. |
|
|
Giwaz S, Van Arendonk JAM, Komen H, Windig JJ, Hanotte O (2007). Population structure, genetic variation and morphological diversity in indigenous sheep of Ethiopia. Animal Genetics 38(6):621-628. |
|
|
Hailemariam F, Gebremicheal D, Hadgu H (2018). Phenotypic characterization of sheep breeds in Gamogofa zone. Agriculture and Food Security 7(1):1-7. |
|
|
Katongole JBD, Sebolai B, Madimabe MJ (1996). Morphological characteristics of the Tswana goat. In. Lebbie SHB, Kagwini E (Eds.), Proceedings of the Third Biennial Conference on the African Small Ruminant Research Network. Kampala, Uganda. pp. 43-47. |
|
|
Kunene NW, Nesamvuni EA, Fossey A (2007). Characterization of Zulu (Nguni) sheep using linear body measurements and some environmental factors affecting these measurements. South African Journal of Animal Science 37(1):11-20. |
|
|
Makhabu SW, Marotsi B, Perkins J (2002). Vegetation gradients around artificial water points in the Central Kalahari Game Reserve of Botswana. African Journal of Ecology 40(2):103-109. |
|
|
Melesse A, Banerjee S, Lakew A, Mersha F, Hailemariam F, Tsegaye S, Makebo T (2013). Morphological characterization of indigenous sheep in Southern Regional State, Ethiopia. International Journal of Animal Genetic Resources 52:39-50. |
|
|
Michael A, Kefelegn K, Yoseph M (2016). Phenotypic Characterization of Indigenous Sheep Types In Northern Ethiopia. Journal of Natural Sciences Research 6(15):16-27. |
|
|
Mohammed N, Alemayehu K, Getachew T (2017). On-farm phenotypic characterisation of indigenous sheep population and its cross with Awassi in Selected Districts of South Wollo, Amhara, Ethiopia. Tropical Drylands 1(1):1-11. |
|
|
Monau PI, Visser C, Nsoso SJ, Van Marle-Köster E (2018). Phenotypic and genetic characterization of indigenous Tswana goats. South African Journal of Animal Science 48(5):925-934. |
|
|
Msanga YN, Mwakilembe PL, Sendalo D (2012). The indigenous cattle of the Southern Highlands of Tanzania: distinct phenotypic features, performance and uses. Livestock Research for Rural Development 24:7. |
|
|
Nsoso SJ, Podisi B, Otsogile E, Mokhutshwane BS, Ahmadu B (2004b). Phenotypic Characterization of Indigenous Tswana Goats and Sheep Breeds in Botswana: Continuous Traits. Tropical Animal Health and Production 36(8):789-800. |
|
|
Nsoso SJ, Madimabe MJ (1999). A review of the sheep industry in Botswana: Promoting the Karakul sheep industry. South African Journal of Animal Science 29(3):258-262. |
|
|
Okpeku M, Yakubu A, Sunday P, Ozoje, MO, Ikeobi C, Adebambo O, Ikhide I (2011). Application of multivariate principal component analysis to morphological characterization of indigenous goats in Southern Nigeria. Acta agriculturae Slovenica 98(2):101-109. |
|
|
Otoikhian CSO, Otoikhian AM, Akporhuarho OP, Oyefia VE, Isidahomen CE (2008). Body measurement parameters as a function of assessing body weight in goats under on-farm research environment. African Journal of General Agriculture 4(3):135-140. |
|
|
Schierenbeck KA (2017). Population-level genetic variation and climate change in a biodiversity hotspot. Annals of Botany 119(2):215-228. |
|
|
Shibabaw W (2012). On-farm Phenotypic Characterization of Hararghe Highland Sheep and Their Production Practices in Eastern Hararghe Zone, Ethiopia. [Thesis]. School of Graduate Studies of Haramaya University, Ethiopia. |
|
|
Söderquist L, Hulten F (2006). Normal values for the scrotal circumference in rams of Gotlandic Breed. Reproduction in Domestic Animals 41(1):61-62. |
|
|
Sölkner J, Nakimbugwe H, Valle ZA (1998). Analysis of determinants for success and failure of village breeding programmes. In. Proceedings of the Sixth World Congress on Genetics Applied to Livestock Production. Australia pp. 273-280. |
|
|
Soma P, Kotze A, Grobler JP, Van Wyk JB (2012). South African sheep breeds: Population genetic structure and conservation implications. Small Ruminant Research 103(2-3):112-119. |
|
|
Song S, Dey DK, Holsinger KE (2006). Differentiation among populations with migration, mutation and drift: implications for genetic inference. Evolution 60(1):1-12. |
|
|
Sowande OS, Sobola OS (2007). Body measurements of West African dwarf sheep as parameters for estimation of live weight. Tropical Animal Health Production 40(6):433-439. |
|
|
Tassew M (2012). On-Farm Phenotypic Characterization of Native Sheep Types and Their Husbandry Practices in North Wollo Zone of the Amhara Region. M.Sc. Thesis presented to the School of Graduate Studies of Haramaya University. |
|
|
Tesfaye G (2008). Characterization of Menz and Afar Indigenous Sheep Breeds of Smallholders and Pastoralist for Designing Community Based Breeding Strategies in Ethiopia. An M.Sc. Thesis presented to the School of Graduate Studies of Haramaya University, Dire Dawa, Ethiopia. |
|
|
Tibbo M, Ginbar T (2004). Phenotypic characterization of the Bonga sheep in Kaffa zone: Field mission Report 8 p |
|
|
Tibbo M, Ayalew W, Awgichew K, Ermias E, Rege JEO (2004). On-station characterization of indigenous Menz and Horro sheep breeds in the central highlands of Ethiopia. Animal Genetic Resources Information 35:61-74. |
|
|
Wilson RT, Durkin JW (1984). Age at permanent incisor eruption in indigenous goats and sheep in semi-arid Africa. Livestock Production Science 11(4):451-455. |
|
|
Yoseph M (2007). Reproductive traits in Ethiopian male goats, with special reference to breed and nutrition PhD dissertation. Department of Clinical Sciences, Faculty of Veterinary Medicine and Animal Sciences, Swedish University of Agricultural Science (SLU), Uppsala, Sweden. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


