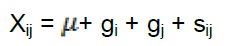ABSTRACT
Verticillium wilt disease causes significant losses to cotton yield and varietal development for resistance is critical in combating this threat. A study was carried out to determine the underlying genetic pattern controlling disease resistance and identify suitable parental lines to use in varietal tolerance development. Five cotton varieties were crossed in a half diallel mating system to produce ten crosses and five selfed parental lines which were screened against Verticillium wilt by artificial inoculation. There were significant variations on the severity scores, vascular brown index scores, morphological and agronomical traits under Verticillium wilt pressure. The Verticillium wilt severity scores ranged from 1.22 to 3.07 and Vascular Brown Index scores from 0.78 to 2.77. The mean squares of general combining ability (GCA) for the various characteristics of the parents were significant and also for the specific combining ability (SCA) of the vascular brown index score which was 0.27 was also significant. Three parental lines were identified as breeding material with good GCA, morphological and agronomical performance and these were CRI-MS-1, SZ9314 and BC853. This study implies that varietal tolerance can be developed by incorporating genotypes with resistance genes in breeding programmes. Future work should focus on developing resistant varieties suitable for production in Verticillium wilt prone areas.
Key words: Verticillium wilt, general combining ability, specific combing ability, cotton.
Cotton productivity has generally been low in Africa due to the effect of climatic variations, biotic stress and poor crop management by farmers (International Cotton Advisory Committee, 2012). In the Southern African region, cotton industry plays a pivotal role in the national economies of Zimbabwe, South Africa, Mozambique, Zambia and Malawi (Baffes, 2004). Annually in Zimbabwe, 300 000 ha of land is put under cotton and about 250 000 tonnes of seed cotton is realised with a national average yield of around 800 kg per hectare (Mudada, 2013) which is low compared to the expected yield of 2300 kg per hectare in local varieties (Mudada and Muripira, 2010). Smallholder farmers who account for about 99% of cotton production in Zimbabwe contribute to the low yields because of poor production management practices (Mubvekeri and Nobanda, 2012).
Cotton diseases including Verticillium wilt caused by Verticillium dahliae Kleb have also been cited as a major reason for the low yields attained by the smallholder farmers (Mapope, 2001). The disease was also reported to be a problem in South Africa, Tanzania, Uganda, Ethiopia and Malawi in a survey report made by International Cotton Advisory Committee (2003).
Cotton has many characteristics that are quantitatively inherited and the most interesting ones being for biotic stress tolerance (Meredith, 1998), especially for disease resistance. Understanding genetic variation and disease resistance, therefore, is important for plant breeders and pathologists because the selection of resistant genotypes depends on high heritability of the traits of the crop influencing resistance (Ashraf and Ahmad, 2000). In breeding programmes of self-pollinated crops, combining desirable genes from different germplasm through recombination is important so as to develop a single superior genotype (Mudada, 2013).
More specific, in plant breeding, identifying suitable parents for the crossing programme is important. Lines or parents in hybrid combinations can be compared on their performance through combining ability analysis (Griffing, 1956). The combining ability is a powerful tool used to determine hybrid vigour in a population and thereby aid in selecting parents for producing crosses and segregating populations (Meredith, 1998). General combining ability and specific combining ability analysis are tools used to separate good and poor combiners followed by selection of the best cross combination. One of the techniques widely used for this purpose in different crops, including cotton, is diallel analysis (Hayman, 1954). Diallel mating helps in choosing parents to be used in hybridisation or population breeding programmes (Mather and Jinks, 1982). The aims of this study were to study the underlying genetic pattern controlling disease resistance by determining the combining abilities, phenotypic correlations and yield of the cotton lines and identify suitable parents to be used in cotton breeding programme for the improvement of Verticillium wilt tolerance.
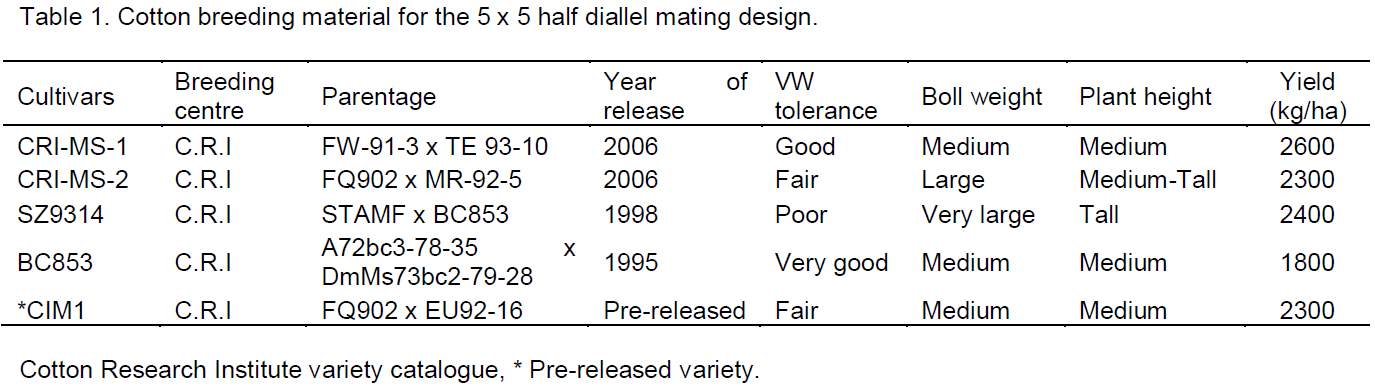
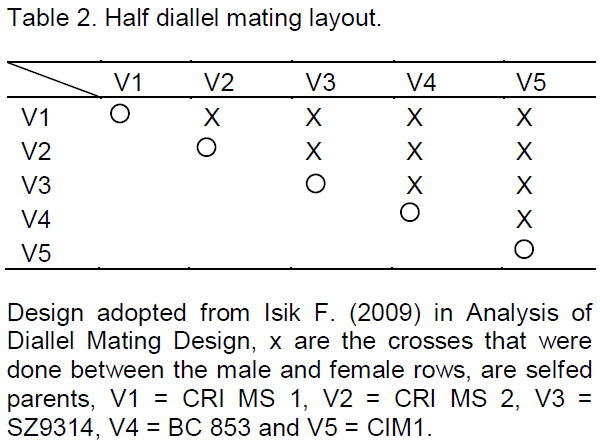
The experiment was conducted at Cotton Research Institute (C.R.I) in Zimbabwe which lies at 18° 20′ South and 29° 54′ East at an altitude of 1156 m above sea level. The institute receives an average rainfall of 750 mm per annum and the temperature during the summer season averages 28oC maximum and 14°C minimum ranges (Mubvekeri and Nobanda, 2012). Five diverse upland cotton Zimbabwean genotypes (CRI-MS-1, CRI-MS-2, SZ9314, BC 853 and CIM1) developed by the Cotton Research Institute varying in year of release, verticillium wilt response and seed cotton yield traits (Table 1) were crossed in a half diallel mating fashion. The half diallel mating design which included parent lines was carried out in the greenhouse during the winter period of 2013. The genotypes were crossed to each other and the parent lines selfed but reciprocals excluded in a non-replicated crossing block with four pots per cultivar as shown in the layout in Table 2 as described by Isik (2009).
The 10 F1 hybrids produced and their five selfed parents were evaluated under Verticillium wilt pressure in the greenhouse at C.R.I in 2014. The 15 genotypes were laid out in a randomised complete block design with three replications each. The plot sizes were composed of three pots containing single cotton plants that were spaced at 1m between rows and 0.3 m within rows. The blocking factor was shading imposed by one brick walled side of the greenhouse. Artificial inoculation of Verticillium wilt was done on the cotton plants to induce uniform infection in the plants at six weeks after crop emergence as follows:
A collection of five cotton plants showing foliar symptoms of verticillium wilt were collected from the field at C.R.I. Leaves from these cotton plants were used for isolation of the pathogen by surface sterilising them using 1% Sodium hypochlorite and cleansed with running distilled water. Twenty small sections of 5 x 5 mm were then cut from the margin of the lesions on the leaves such that the leaf piece contained both the healthy and diseased leaf tissue. The five tissues samples were placed in a petri dish containing Potato Dextrose Agar prepared in an autoclave at 120°C for 20 min.
Twenty petri dishes were prepared under a lamina airflow cabinet and they were incubated for 48 h at 26°C under dark conditions. Purification of the fungal growth was done by sub-culturing in Czapek Dox Broth media agar containing 3% solution of lactic acid to inhibit bacterial growth (Hillocks, 1991). Microsclerotia developed on the culture and the inoculum for each isolate was adjusted to a concentration of 2x106 conidia/ml using repeated serial dilutions in distilled water. Six weeks old cotton seedlings were inoculated at the base of the plant using sewing needles dipped in the culture suspension as described by Mapope (2001). The fungal isolates used for inoculation had been cultured in the broth media for 48 h.
Comparison of varietal performance of the genotypes under Verticillium wilt pressure was carried out on disease expression (infection scores), Vascular Brown Index (VBI), plant height, fruit retention and bolls per plant, boll weight and seed cotton yield expected per hectare. The severity of the disease and resistance was measured by means of scoring foliar symptoms and using VBI determined on the cross sectional discolouration of stems cut after harvesting according to the Fusarium and Verticillium Resistance Ranking Protocol for Cotton Varieties (2008), Australian Cotton CRC. The scoring system that was used was given in Tables 3 and 4.


The general agronomic practices as recommended in the Cotton handbook (Mubvekeri and Nobanda, 2012) were carried out on the cotton crops. Seed cotton picking was done at the end of the season on the whole plot. The data recorded on disease infection score expression, Vascular Brown Index, plant height, fruit retention and bolls per plant, boll weight and seed cotton yield expected per hectare was subjected to analysis of variance (ANOVA) to test with the F-test using Genstat 14th edition statistical package from VSN International bioscience software and consultancy.
Genetic analysis
GCA and SCA were estimated based on the fixed model method II as described by Griffing (1956) as used by Machikowa et al. (2011) as follows:
Where Xij = the mean phenotypic value; = the general mean; gi gj, = GCA effects of the ith and jth parents respectively; sij = SCA effects of the cross i x j.
The estimates of GCA and SCA parents and hybrids were obtained as:
GCA effects gi = 1/n(n-2)[nXi.-2X..]
SCA effects sij = Xij-1/n(n-2)(Xi+X.j) + 2/(n-1)(n-2)X..
Where X
i., X.
j = means of the i
th and j
th parents respectively; X..
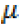
= grand mean; n = number of parent lines.
GCA:SCA ratio was calculated using ratios of the mean squares of GCA and SCA.
Correlation
Phenotypic correlation estimates were calculated using the following formula as used by Lukonge in 2005:
Negative value of correlation coefficient (r) indicates dominant genes, while if its value was positive then recessive genes were responsible for the phenotypic expression of the trait. Genstat 14th edition statistical package was applied for correlation analysis.
Morphological traits
Analysis of variance for the cotton lines revealed presence of significant differences on morphological characteristics (Table 5). The plant height was significantly different at P£0.05 between the 15 cotton lines where CRI-MS-2 was taller than all the genotypes (160.3cm) but was comparable to BC853, the cross between CRI-MS-2 and SZ9314, the cross between SZ9314 and BC853 and the cross between BC853 and CIM1. The cross between SZ9314 and CIM1 yielded the shortest progeny (121.1 cm) and this was comparable to the parental lines SZ9314, CRI-MS-1, CIM1, the crosses CRI-MS-1 x CRI-MS-2, CRI-MS-1 x SZ9314, CRI-MS-1 x SZ9314, CRI-MS-1 x BC853, CRI-MS-1 x CIM1 and CRI-MS-2 x BC853.
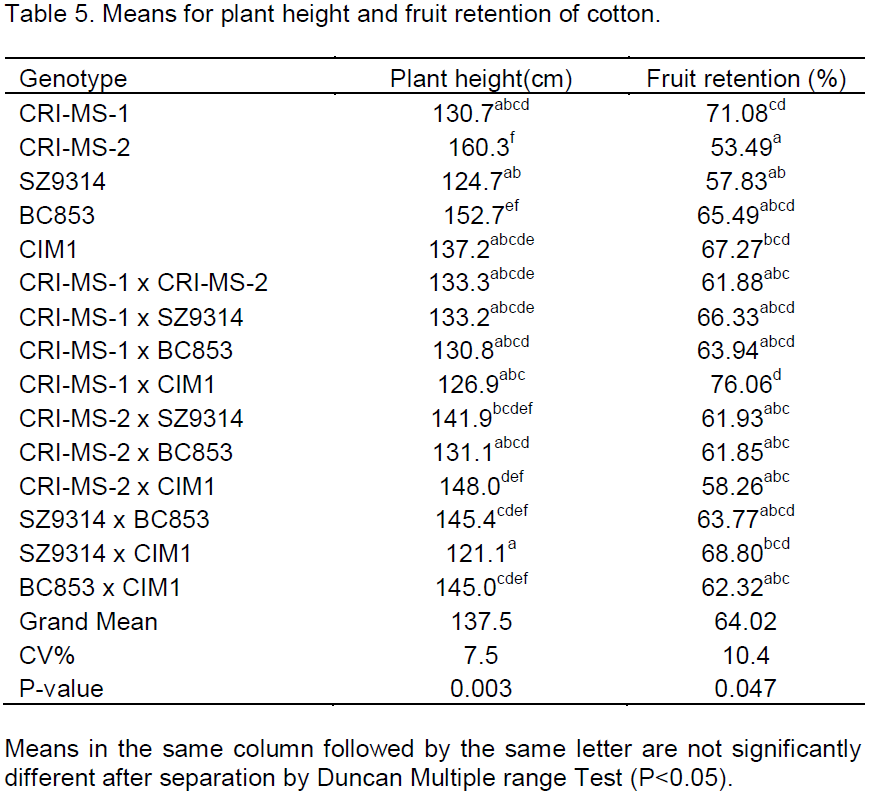
Fruit retention among the 15 genotypes was significantly different (P£0.05). The cross between CRI-MS-1 x CIM1 retained most of its fruiting bolls and it was comparable to both parents CRI-MS-1 and CIM1 and also to BC853 and the crosses CRI-MS-1 x SZ9314 and CRI-MS-1 x BC853. CRI-MS-2 shedded most of its bolls as it had the lowest fruit retention percent along with the parental lines SZ9314 and BC853 as shown in Table 5.
Yield and yield components
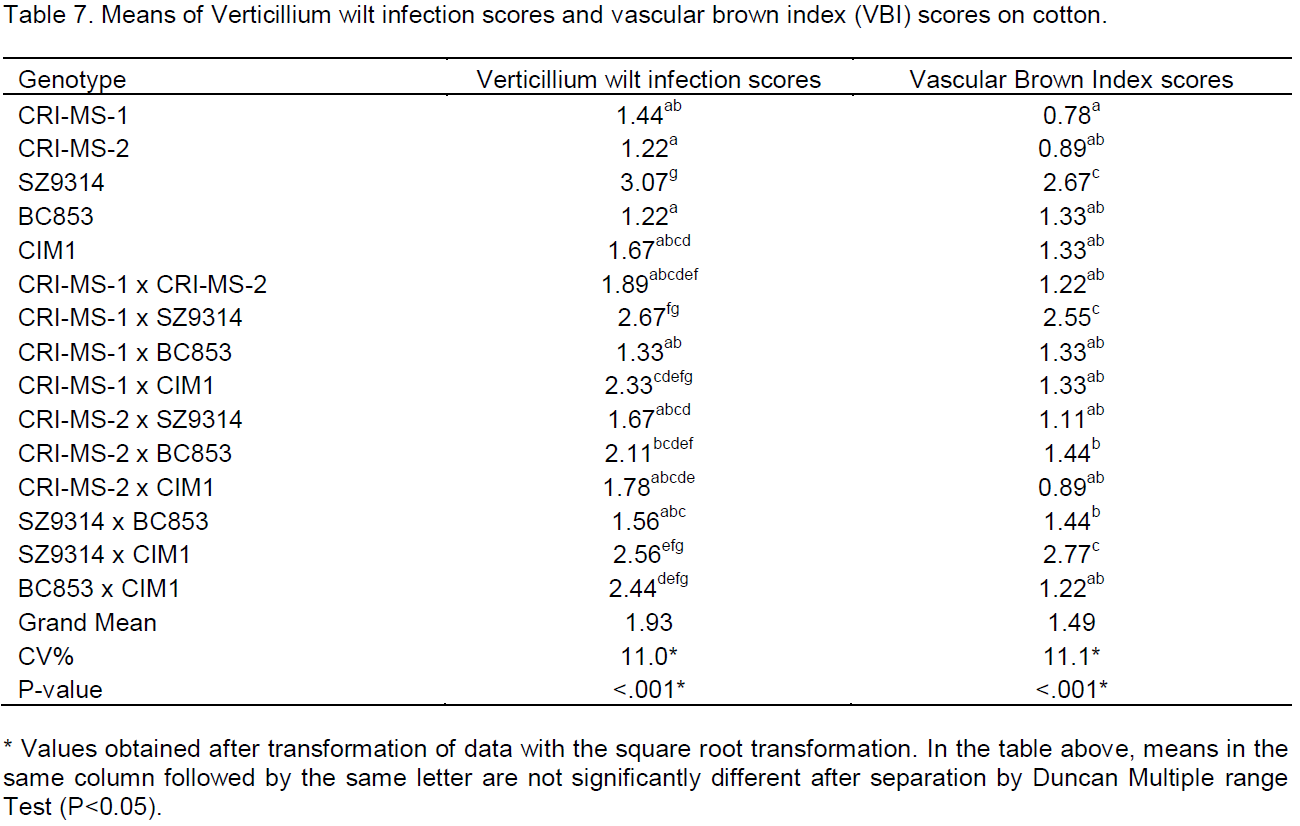
Table 6 also shows varietal performance under the disease pressure in terms of number of bolls produced. The number of bolls per plant also did not vary largely for all the genotypes (P£0.05). The boll number ranged between 12 and 19. There were also no noticeable differences on the average boll weight of the different cotton genotypes which ranged from 3 and 6 g to that which were used in the study (Table 6). There were significant differences in the mean squares of replications for this trait indicating the importance of blocking in this study.
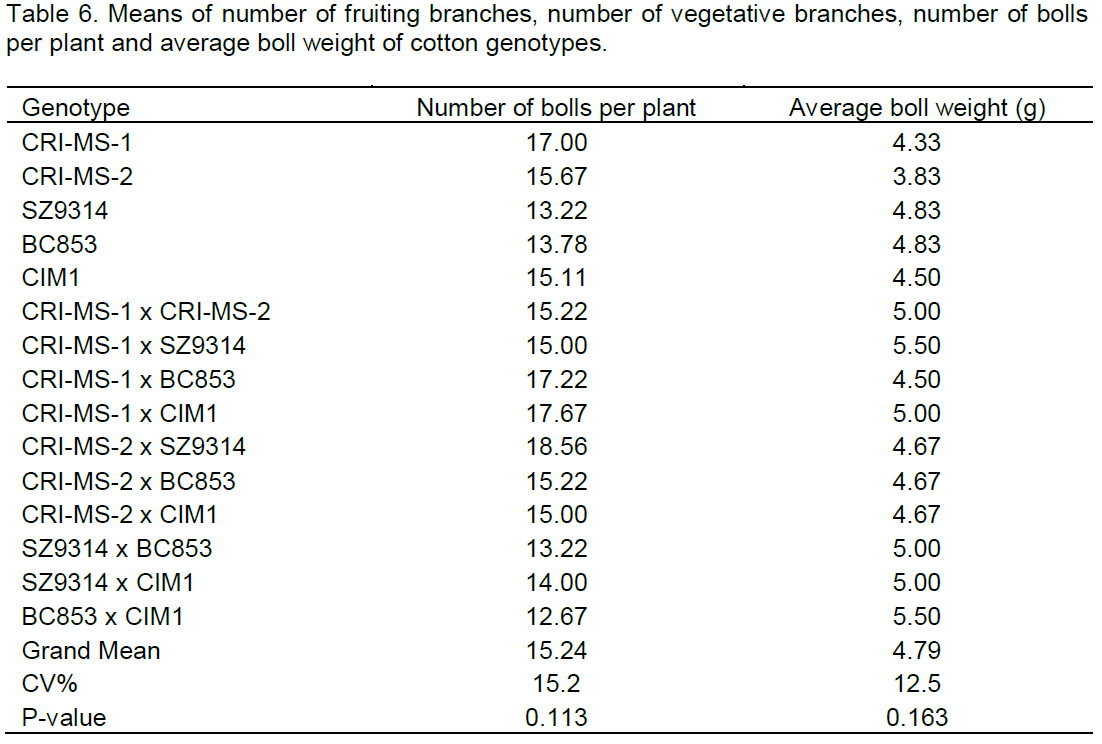
The seed cotton yield of the cotton lines under Verticillium wilt pressure varied largely as the results revealed that the yield means were significantly different at P<0.05 (Figure 1). The cross CRI-MS-1 x SZ9314 had significantly higher yield (2259 kg/ha) although it was comparable to two parent lines CRI-MS-1 and BC853 and six other crosses. The cross between CRI-MS-2 and BC853 yielded the least seed cotton (1296 kg/ha) but the yield was comparable to four parent lines (CRI-MS-2, SZ9314, BC853 and CIM1) and four other crosses.
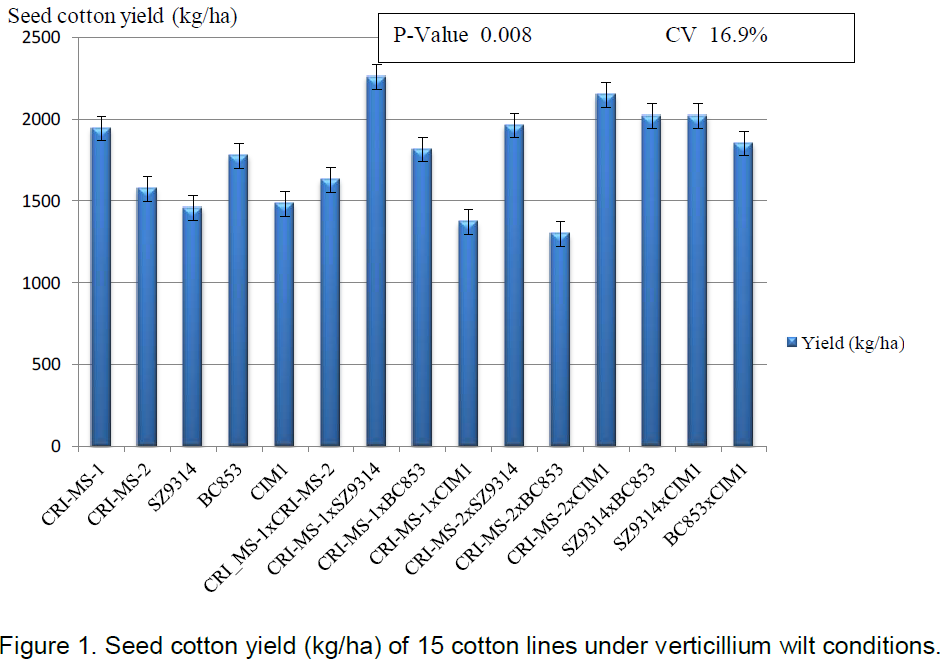
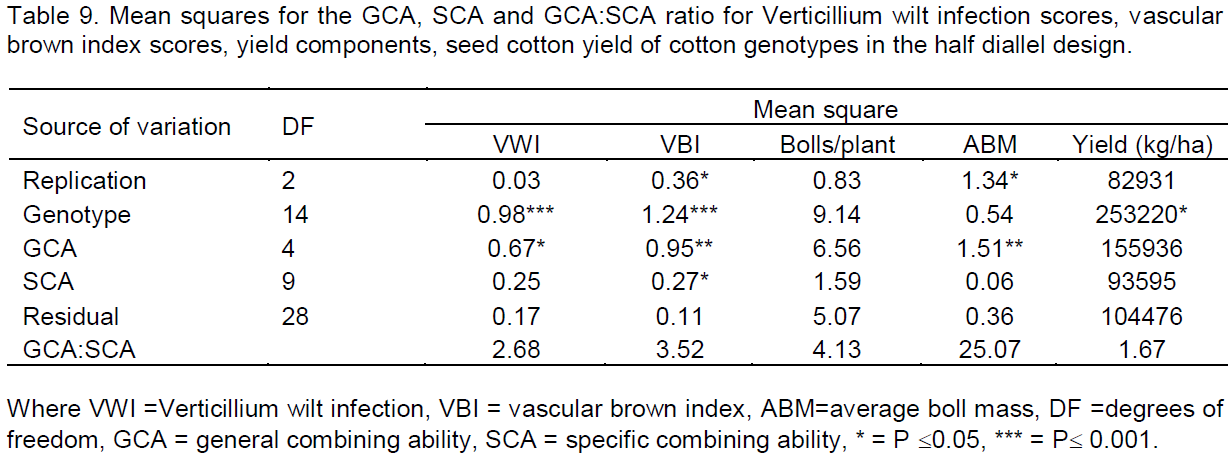
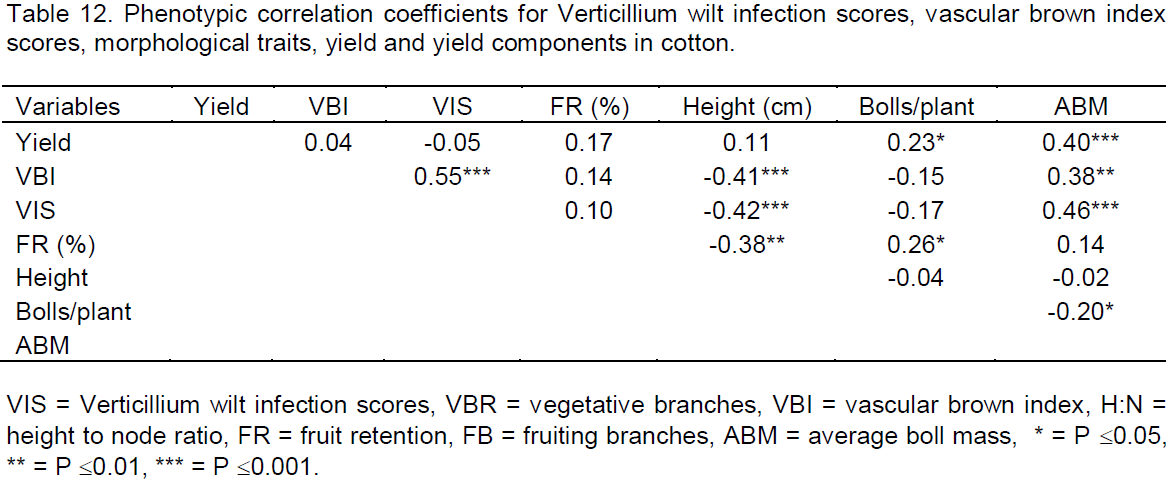
Analysis of variance on the 15 cotton genotypes developed in the diallel mating design revealed highly significance differences among the genotypes when exposed to the same inoculum level of Verticillium wilt disease (P£0.001) in terms of foliar infection scores and vascular brown index scores as shown in Table 7. SZ9314 was affected the most with the disease as shown by the high infection score (3.07) and high vascular brown index score (2.67). The genotype was comparable to its cross with CRI-MS-1 (infection score of 2.67 and high vascular brown index score of 2.55) and CIM1 (infection score of 2.56 and high vascular brown index score of 2.77). CRI-MS-2 and BC853 were the most tolerant genotypes as shown by their low infection scores (1.22) and low vascular brown index scores (0.89 and 1.33 respectively). All the genotypes were infected by the disease to some extent as indicated by varying scores. The Verticillium wilt scores ranged from 1.22 to 3.07 and the vascular brown index scores ranged from 0.78 to 2.77.
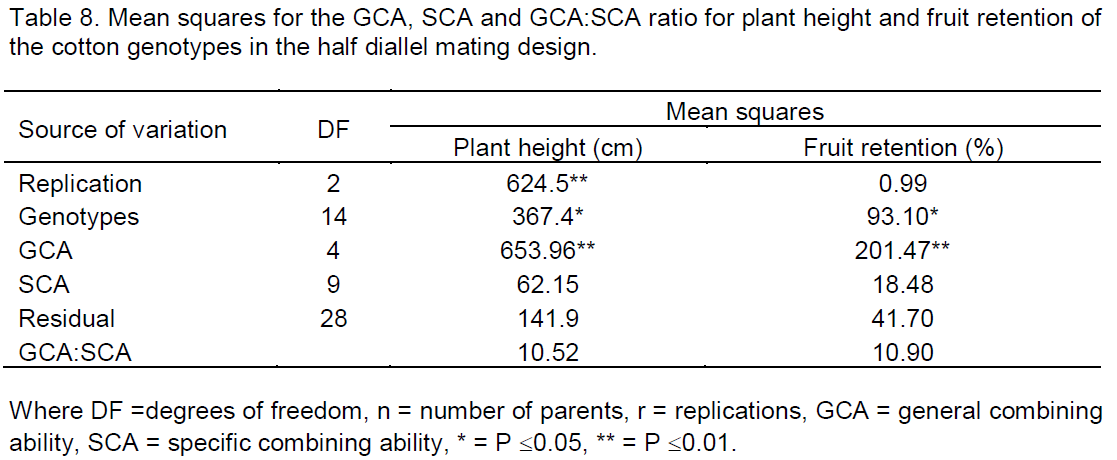
Analysis of parents and F1s using the Griffing’s approach were employed to study the GCA effects and SCA effects for exploring the possibilities of isolating the best recombination related to Verticillium wilt resistance. Mean squares of GCA were significant for plant height, height to node ratio, fruit retention, number of fruiting branches, vascular brown index scores and the average boll mass at 0.01 significance level whilst Verticillium wilt infection scores was significant at 0.05 significance level (Tables 8 and 9). The significant mean square GCA effects indicated that additive genetic effect was controlling most of the characteristics. Mean squares of SCA were significant for vascular brown index scores at 0.05 significance level (Table 9) indicating non additive genetic effect influenced the expression of the scores.
GCA effects
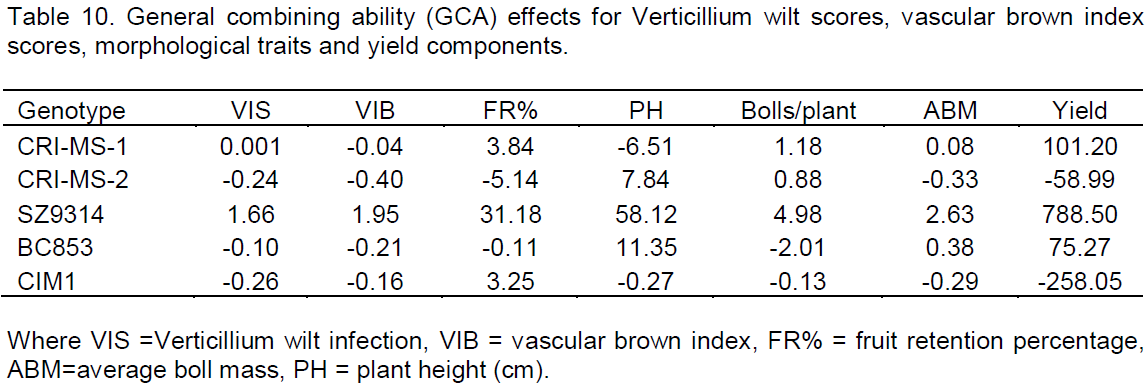
The GCA contributions of each parent to the various characters influencing resistance against Verticillium wilt are shown in Table 10. Variability of the GCA was observed between the five parental lines with SZ9314 showing the highest positive GCA values for all the seven parameters tested. CRI-MS-1 which was the second best combiner, had positive GCA values for Verticillium wilt scores, fruit retention, bolls per plant, average boll mass and seed cotton yield. BC853 also had better GCA values on plant height, average boll mass and seed cotton yield. CRI-MS-2 had better combining abilities for plant height and number of bolls per plant. CIM1 had negative GCA effects for all the characteristics except for fruit retention. CRI-MS-2 and BC853 both had negative GCA effects on Verticillium wilt scores, vascular brown index scores and fruit retention. BC853 also had a negative GCA value for number of bolls per plant.
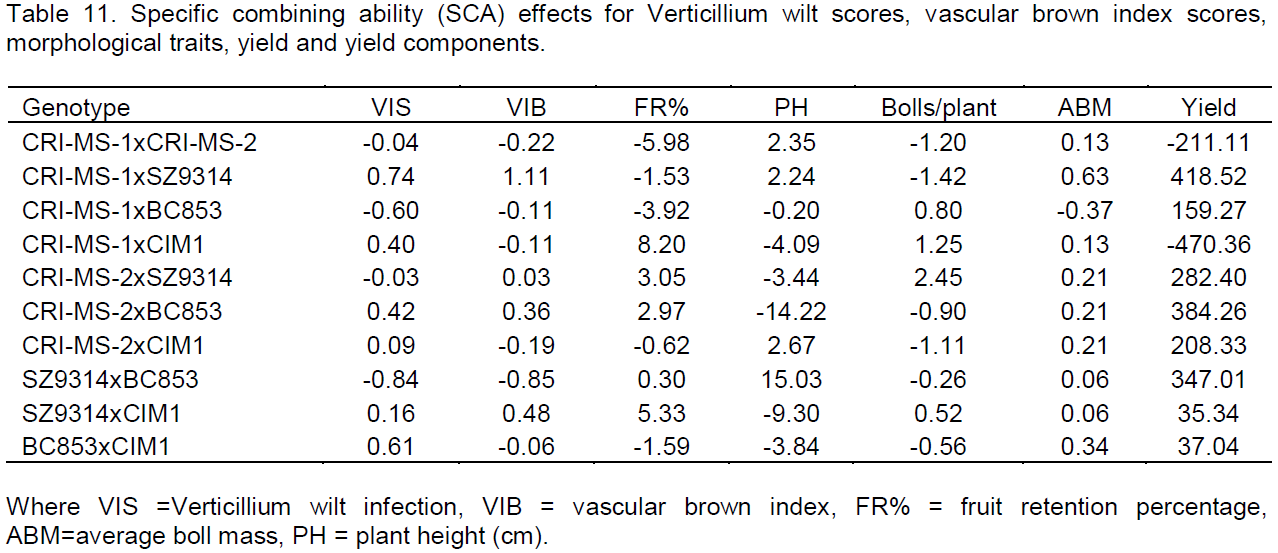
SCA effects
SCA estimates (Table 11) indicate that CRI-MS-1xSZ9314 was a best cross (0.74) and SZ9314xBC853 being the worst cross (-0.84) in terms of Verticillium infection scores. The vascular brown index scores also revealed the same (CRI-MS-1xSZ9314 having a positive SCA value of 1.11 and SZ9314xBC853 having a negative SCA value of -0.85). In terms of fruit retention, CRI-MS-1xCIM1 proved to be the best combination with a positive SCA effect (8.20) and CRI-MS-1xCRI-MS-2 being the worst (-5.98). The plant height performance of the crosses revealed that SZ9314xBC853 was the best combination (15.03 SCA value) and the cross CRI-MS-2xBC853 being the worst (-14.22). The cross CRI-MS-2xSZ9314 was the best combination for the number of bolls per plant (2.45) and CRI-MS-1xSZ9314 being the worst (-1.42). For the average boll mass, all the crosses had positive SCA effects with the cross CRI-MS-1xSZ9314 being the best (0.63) whilst CRI-MS-1xBC853 was the only cross with a negative SCA effect. The cross CRI-MS-1xSZ9314 had a superior SCA effect in terms of seed cotton yield (418.52) whilst CRI-MS-1xCIM1 was the worst combination (-470.36).
Correlation
Phenotypic correlations for most of the characteristics were positive and but not significant as shown in Table 12. Most of the characteristics that were negatively correlated were significant. Vascular brown index scores was significantly correlated to Verticillium wilt infection scores (0.55) at P£ 0.001. There was a significant positive correlation between the seed cotton yield and the number of bolls per plant (0.23) at P£ 0.05 and highly significant association between the seed cotton yield and the average boll mass (0.40) at P£0.01. This indicate that average boll mass is important to cotton yield. Verticillium wilt infection scores had a high significant negative correlation with height (-0.42) at P£ 0.001. Fruiting retention was not highly associated with the plant height (-0.38). The seed cotton yield was positively correlated with all the traits except for Verticillium wilt infection scores where it had a negative association.
There were some noticeable level of variations observed for the morphological and yield components studied among the c Botton parental lines and their F1 progeny. The study revealed that selection of cotton lines under Verticillium wilt could be done for varietal resistance against the disease, plant height, fruit retention and seed cotton yield. The parental lines CRI-MS-1 and BC853 had better yields under the Verticillium wilt pressure. These findings are in agreement with the information indicated in the variety catalogue by Mudada and Muripira (2010) which showed that the varieties are resistant to the disease. These varieties can also be used as breeding material for varietal tolerance development programme against Verticillium wilt in cotton as indicated by superior crosses including at least one of the parental lines on almost all traits that were studied. Verticillium wilt infections and vascular brown index scores were highly significant indicating differences in genetic make-up of the parental lines and the F1 progeny in terms of reaction to disease infection. SZ9314 was the only parental line that was highly affected by the disease indicating the lack of dominant genes that supressed the progression of the disease in the cotton plants of the variety. The crosses involving this parental line with other tolerant varieties however yielded better performing progenies against the disease and better seed cotton yields in the presence of the diseases.
Significant mean squares of the GCA effects that were observed revealed the presence of additive genetic effect controlling the expression of most of the traits that were studied except for the number of bolls per plant and seed cotton yield. This is in contrast with studies by Lukonge (2005) which revealed significant mean squares for bolls per plant and seed cotton yield which suggests that these could be used to improve yield under the disease pressure. The results from the diallel study also indicated that there was high mean square contribution of GCA effects to the total mean squares as revealed by GCA:SCA ratios greater than one which confirms the predominant role of additive gene effects to the role of non-additive gene effects for all the characteristics under study. These results were also obtained by El-Dahan et al. (2003). In this case normal, breeding methods such as backcross, pedigree or recurrent selection can be used to fix these additive genes. Significant mean squares of SCA for the vascular brown index scores indicated non-additive genetic influence on the extent discolouration occurred on the vascular bundles due to disease infection. This is rather a product of genetic and environmental interactions where the amount of phytoalexins produced to avoid the upward movement of the pathogen are determined by the amount level of pathogen inoculum and temperature (Mysore and Ryu, 2004).
The cotton parental lines used in this study varied for GCA estimates on all the parameters tested in the experiment. SZ9314 had positive GCA effects for all the traits that were studied indicating that it is a good combiner if used in breeding programmes for varietal tolerance development in cotton under Verticillium wilt conditions. This is in agreement with varietal performance of the crosses including the variety in terms of seed cotton yield, Verticillium wilt infection scores, vascular browning index scores and the other morphological traits. CRI-MS-1 had some characteristics that exhibited positive GCA estimates with vascular brown index, number of fruiting branches, plant height and height to node ratio having negative GCA values. This indicates that these traits are negatively correlated and could lead to challenges during breeding of varietal tolerance against Verticillium wilt. BC853 had five positive GCA estimates and five negative GCA estimates signifying the same challenge as noted for CRI-MS-1. These varieties however were the best performers in terms of combining ability if incorporated in breeding programmes that are targeted at reducing the impact of the disease in cotton especially if yield is to be considered. CRI-MS-2 and CIM1 had most negative GCA estimates for most characteristics indicating that they are poor combiners if used in breeding programmes. According to Griffing (1956), GCA estimates close to zero show that genotype does not differ much from the general mean of all crosses and positive and negative values indicate good and poor parent when used in the diallel.
Positive and negative SCA estimates were observed in the crosses of cotton produced in the diallel mating system under Verticillium wilt conditions. Positive SCA effects were observed for crosses that included CRI-MS-1, SZ9314 and BC853 parental lines for most of the characteristics. This is a confirmation that the parental lines can be used in the breeding programmes with high chances of improving the characteristics so that they can be fixed and used in the development of tolerant lines against Verticillium wilt. Lukonge (2005) suggested that self-pollinating crops like cotton fixes the additive by additive type of component at later stages of inbreeding.
In cotton breeding, the phenomenon of negative correlation exists whereby improvement of certain economic important traits results in breakdown of other traits (Mudada, 2013). This is the case with the results from this study where negative and positive correlation values where observed. The negative correlation of the Verticillium wilt infection scores with yield, plant height and number of bolls per plant will further derail the efforts of breeding for tolerance against the disease in cotton. However, these results suggest that the phenotypic variance for most of the characteristics was mostly additive and those with low values being non-additive.
Three parental cotton lines with high and positive GCA, namely CRI-MS-1, SZ9314 and BC853, and positive SCA combination value for the characteristics studied were identified in this study. This implies that varietal tolerance development in cotton can be carried out using parental lines with good combining abilities in terms of disease resistance and high yielding. The best hybrids that can be further advanced to fix the genes controlling the resistance traits were also identified in the form of the crosses CRI-MS-1xSZ9314, CRI-MS-1xBC853 and SZ9314xBC853. These were recommended for further use as breeding material through selfing in subsequent filial generations in the Verticillium wilt tolerance development programme.
The authors have not declared any conflict of interest.
REFERENCES
|
Baffes J (2004). Tanzania's cotton sector: Constraints and challenges in a global environment. African Region Working Paper Series No. 42. |
|
|
|
El-Dahan MAA, Leid EO, Lopez M, Gutierrez JC (2003). Combining Ability of Yield and Yield Components in Upland Cotton (Gossypium hirsutum) Under Drought Stress Conditions. Proceedings World Cotton Research Conference. Cape Town, South Africa. |
|
|
|
Griffing B (1956). Concept of general and specific combining ability in relation to diallel crossing system. Aust. J. Biol. Sci. 9:463-493. |
|
|
|
Hayman BI (1954). The theory and analysis of diallel crosses. J. Genet. 39:789-810. |
|
|
|
Hillocks RJ (1992). Cotton Diseases. London, UK: CAB International International Cotton Advisory Committee (ICAC) (2003). Assessment of the Impact of the Main Dynamics of Cotton Diseases Affecting in Particular Small-scale Production Systems in Southern and Eastern Africa. 1629 K Street NW, Suite 702, Washington, DC 20006 USA. |
|
|
|
International Cotton Advisory Committee (ICAC) (2012). World cotton situation. 1629 K Street NW, Suite 702, Washington, DC 20006 USA. |
|
|
|
International Cotton Advisory Committee (ICAC) (2014). World Prices High Despite Excess Production. 1629 K Street NW, Suite 702, Washington, DC 20006 USA. |
|
|
|
Isik F (2009). Analysis of Diallel Mating Designs. North Carolina State University, Raleigh, USA. |
|
|
|
Lukonge EP (2005). Characterisation and Diallel Analysis of Commercially Planted Cotton (Gossypium hirsutum L.) Germplasm in Tanzania. PhD Dissertation, University of the Free State, South Africa. |
|
|
|
Machikowa T, Saeteng C, Funpeng K (2011). General and Specific Combining Ability for Quantitative Characters in Sunflower. J. Agric. Sci. 3:1. |
|
|
|
Mapope N (2001). Verticillium Wilt Survey in Cotton Research Institute Annual Report, Department of Research and Specialist Services, Ministry of Agriculture, Zimbabwe. |
|
|
|
Mather K, Jinks JL (1982). Introduction to Biometrical Genetics. Chapman & Hall Ltd. London. McCarty JC, Jenkins JN. DOl: 10.1007/978-1-4613-3387-6 |
|
|
|
Meredith WR (1998). Continued Progress in Plant Breeding for Yield in the U.S.A. Proceedings World Cotton Research Conference Athens, Greece. pp. 97-101. |
|
|
|
Mubvekeri W, Nobanda S (2012). Cotton Agronomy. CRI, Bag 765, Kadoma, Zimbabwe. |
|
|
|
Mudada N (2013). Genotype by Environmental Interaction on Yield, Yield Components and Fibre Quality Parameters of Gossypium hirsutum varieties. MSc Dissertation, Midlands State University, Zimbabwe. Unpublished. |
|
|
|
Mudada N, Muripira V (2010). Cotton Variety Catalogue. Unpublished. Cotton Research Institute, P. Bag 765, Kadoma, Zimbabwe. |
|
|
Mysore KS, Ryu CM (2004). Non host resistance: how much do we know? Trends Plant Sci. 9(2):97-104.
Crossref |