Studies on morphology and occupancy area of trees provide knowledge on features and functions of forest ecosystem, especially in terms of competition and growth of trees. The main purpose of this work was to develop models for shape and size relationships of Araucaria angustifolia in natural forest remains of species. Dendrometric and morphometric variables from 210 trees from three sites were used. Results showed that the species present different shape and size, vigor and productive capacity in evaluated sites. Fitted models indicated that there was transition in competition mode (symmetric or asymmetric) for resources, which is result of available growing space for trees. Relationship between shape and size is derived from slope of regression line and regarded as an indication of need for silvicultural interventions to reduce competition, which is a key information for obtaining better practices in forest management.
Biometric principles of size, shape and growth of trees in forest, and changes of these variables over time are important factors on an individual scale, in forest and ecosystem dynamics. Conclusions on these relationships and forest-level performance are necessary for forest management, since they help to identify the interactions of competition and its effect on growth over time (Pretzsch and Dieler, 2010).
The study on the growth-size relationship and environmental conditions is of particular interest since growth of trees and forest dynamics are strongly affected by trends of changes in growth conditions and stress events (Spiecker et al., 1996; Jentsch et al., 2007; Matyssek and Sandermann, 2003; Pretzsch et al., 2010).
Studies have demonstrated that competition mode and relation between growth and size may vary spatially over ecological gradients (Pretzsch and Biber, 2010), as well as with time as forest develops. According to same authors, forest structure parameters are used to analyze and model its dynamics.
Developing of morphometric variables and changes in forest structure for long periods affects to a large extent, processes such as light absorption, water interception, evapotranspiration, photosynthesis and respiration, and in turn, these processes affect the growth of trees and life cycle of organisms around (Pretzsch, 2009).
Canopy structure is crucial to understand relationships between functionality, structure and environment in pure stands, but more importantly in mixed stands, where different species demonstrate their ability to adapt their structure to capture scarce resources efficiently or deny access to competitors for same resources (Preztsch, 2009).
Shape and size relationships of trees can predict space required by one during its development, inferring to its competitors and allowing conclusions on stability, vitality and productivity of each individual, acting as a practical tool in forestry interventions, especially when age of the plants is not known (Durlo, 2001).
Therefore, developing accurate models of morphometric relationships for tree species in its natural area of occurrence allows identifying trends for sustainable forest management, ensuring ecological and structural sustainability of forests, and related benefits.
The purpose of this study was to evaluate morphometric characteristics and shape and size relationships in individual trees, developing models to compare effect of site on trees to assist in forest management strategies and silvicultural interventions in natural patches with Araucaria angustifolia (Bertol.) Kuntze in Southern Brazil.
Study area
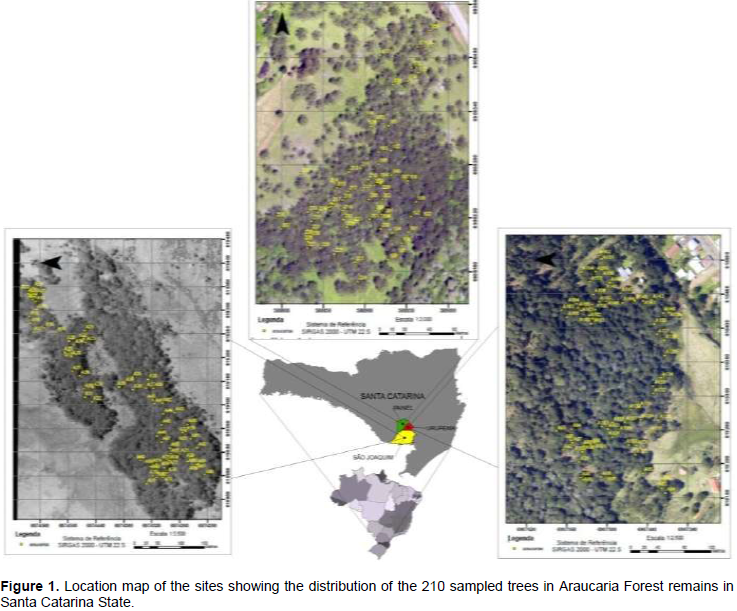
The study was done in natural occurrence region of Araucaria angustifolia, Southern Brazil. Three sites were evaluated: São Joaquim (1,353 masl); Painel (1.145 masl); and Urupema (1.245 masl) counties (Figure 1).
Araucaria angustifolia is an evergreen species. The shape crown shape has cone to hemispheric form, and at maturity stage, it takes umbel shape. Due to Brazilian Legislation, which has forbidden this species management, at present, Araucaria Forest remains are not subject to management and silvicultural intervention for 40 years, presenting an increased competition.
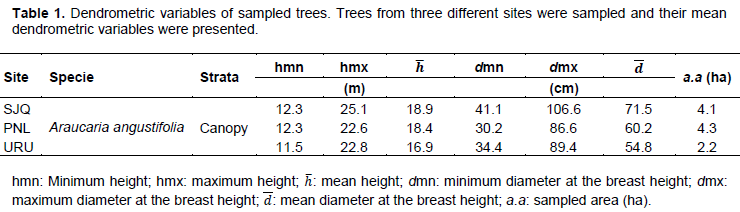
A total of 210 trees were sampled, 70 from each site. Sampling intensity was evaluated accordingly to Sanquetta et al. (2009), assuming a maximum sampling error of 10%. Dendrometric characteristics of sampled trees are shown at Table 1.
Climate classification of the region according to Koppen is predominantly characterized as Cfb, or temperate. The annual mean temperature is 15.5°C, and minimum mean temperatures range between 5 and 8°C, maximum mean between 22 and 31°C and minimum absolute of -12°C, with annual mean rainfall of 1,400 mm (EPAGRI, 2002).
Predominant vegetation type in the region is the Araucaria Forest (IBGE 2012), which is the typical vegetation in the Southern Brazilian Plateau, it has floristic disjunctions in refuges located in mountain ranges from Serra do Mar and Serra da Mantiqueira. In the past, the species had a distribution even northward, since the Araucariaceae family is believed to have paleogeographic dispersion, suggesting different occupation from the present (IBGE, 2012).
Data collection and analysis
In each study site, sample area was defined and sampled trees were label and their positions were established with GPS. The following variables were measure in each tree: diameter at breast height (d), using a diameter tape; the insertion height of crown (
hic); total height (
ht); and crown radius, taken in four directions (North, South, East and West). Height variables and crown radius were measured using a clinometer. Based on the four radius measured, crown mean radius values:
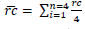
was obtained. Multiplying mean radius, crown diameter:
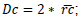
; crown projected area:
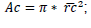
; crown length:
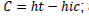
; and crown proportion
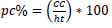
were obtained.
In this study, the hypothesis that morphometric indexes and their variation are indicators of past competition was assumed, which had caused changes in shape-size relationships and can be express by regression functions (Cunha and Finger, 2013). Therefore, considering two trees with same diameter at breast height, that one with smaller crown diameter is one under more competition.
Using sampled data, morphometric variables were calculated and fit of equations of shape and size relationships was carry out. A covariance analysis was performed using morphometric variables as dependent, and their variability explained by interaction with site and diameter as a continuous and independent variable. Thus, difference in level and inclination of regression line was evaluated from the relation between insertion height of crown (hic), crown proportion (pc%) and crown diameter (dc), as a function of diameter at breast height (d) in studied sites, aiming to assess whether a single equation could be used for both sites or multiple equations would be required.
Residuals from regression were tested according to regression assumptions, as proposed by Schneider et al. (2009). The goodness of fit was evaluated by the coefficient of determination (R²), standard error of estimate (Sxy) and graphical analysis of residuals.
Biometric and morphometric adjustments
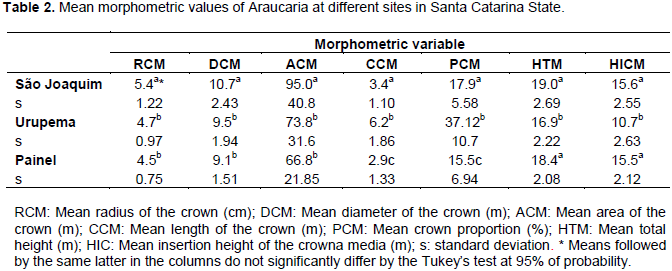
Sample size assessment indicated that 59 trees would be required at the São Joaquim site, 28 trees at Urupema and 20 trees at Painel, in order to accurately represent the populations. Therefore, the sample composed by 70 trees from each site is considered sufficient. Table 2 shows the mean morphometric values from the 70 sampled trees in their respective sites.
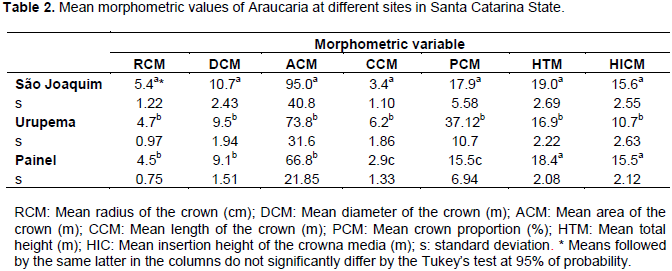
Results indicate that trees from São Joaquim and Painel have larger mean diameter, higher mean total height and higher mean insertion height of the crown. Consequently, these trees have shorter crown and lower crown proportion, which empirically allows one to conclude that, regarding the crown pattern of Araucaria, sampled trees in these sites are older than those from Urupema. Durlo (2001), studying Cabralea canjerana also observed a higher insertion height of the crown and crown diameter as the trees grow. However, this increase in morphometric variables is also associated with space, competition and density of individuals.
Trees from Urupema presented larger mean crown length, indicating that for this site, trees have more growth space and less competition between individuals, since competition effect implies directly the crown length and, consequently, crown proportion. These variables might be regarded as measures of vigor (Assmann, 1970), which are required for models of mortality, growth and yield (Weiskittel et al., 2011).
Wink et al. (2012), studying Eucalyptus sp. Plantations, also noted that proportion of the crown decreases as trees age. According to the authors, crown proportion was significantly different between young stands (20 months) and intermediate stands (44 months), and the late was not different from mature stands (240 months).
It is understood that the greater the crown length, the higher the proportion of the crown, while the inverse relationship is also true. Therefore, the higher the proportion of the crown, the greater will be the photosynthesis capability, and possibly, better growth rates and less age. Santos et al. (2012) pointed out that trees crown is composed by several elements, and it is responsible for the processes of growth and production. Thus, parameters such as area, diameter and crown length are strongly correlated with these physiological processes.
In these cases, the ability to take up space available for the expansion of branches, along with the maintenance of a crown above the space occupied by the stem appears to be important in inter-crown competition situations (Santos et al., 2012).
Covariance analysis and shape and size relationships
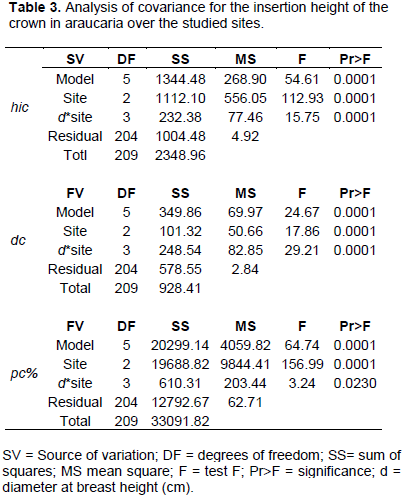
The fitted models presented here aimed to identify and explain relations between shape and size and do not have prediction purposes. Covariance analysis (Table 3) evidenced the presence of differences between inclination levels of regression lines for hic, dc and pc%, thus indicating that the shape and size relationships between sites, morphometry of trees, productivity rates, age and vigor varies with tree’s diameter and characteristics, such as competition and social position.
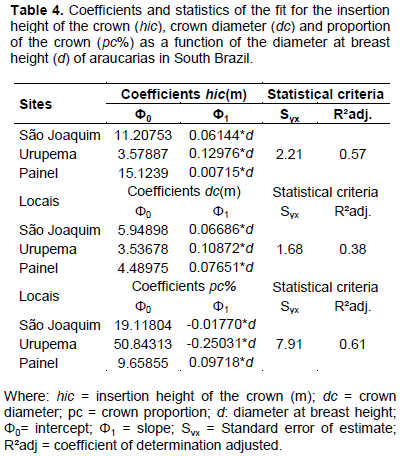
A significant result of the analysis of covariance indicate that the range of variation in shape of trees is regarded to the variation in trees size, as well as to the sort of competition levels at which trees were subject in the forest throughout their development. Such characteristics demand different management practices and morphometric models for the sites, as presented in Table 4.
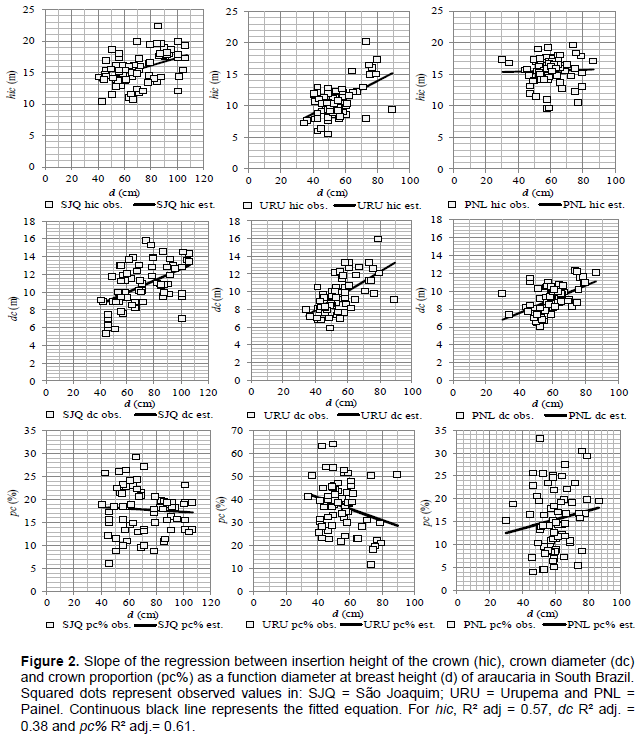
Insertion height of the crown and crown diameter were positively correlated with diameter at breast height, which explained 57 and 38% of the variability in the dependents variable, respectively. For proportion of the crown, the regression line presented a negative slope and diameter at breast height accounted for 61% on its variability.
The fit of equations can be seen in Figure 2, which presents the difference in the level and slope of the shape-size of araucaria. Urupema trees have smaller mean diameter and younger ages, as well as the highest crown proportion (37.2%), which implies larger photosynthetic area and higher growth rates.
In São Joaquim and Painel, trees are larger and older, which contributes to the increase in the insertion height of the crown. Such trees present, therefore, shorter crown and lower growth rates, indicating that they have reached maturity and are in the upper strata of the forest. This means that in these sites, environmental conditions and forest dynamics contributed to accelerate growth and development of forest structure over the past decades (Pretzsch et al., 2014).
The fitted equation for the estimate of hic shows that as araucaria grows and ages, the crown decreases in length, while its diameter increases, which is common for species that in the early stages of development have higher competitive capacity, fast leaf expansion, higher height and root expansion (Zanine and Santos, 2004).
Regression fit indicates that as trees grow in diameter, insertion height of the crown and crown diameter increase, providing an increased usable bole and indicating that trees were able to stand out from competition in the past and expanded their crown.
However, despite the increase in crown diameter, crown proportion decreases, as results of ontogenetic patterns on the species growth. Such reduction shows that growth rates tend to decline with age and reduced crown proportion.
It is understood that trees in an active and accelerated growth phase are shorter and smaller (younger), which present the highest crown proportion as compared to those higher (older) (Wink et al., 2012), varying from 26.2 to 76.7% in this study, for the older and the younger individuals, respectively. The amplitude of the crown proportion is the result of different degrees of competition to which the trees are submitted to and may also be associated with the size of individuals (Roman et al., 2009).
Correlation between morphometric variables of araucaria refers to the individuals position in the canopy, competition (density), size and time. The inverse relationship between diameter at breast height and crown proportion is an indication that as the height of trees stabilize, in large and old trees, there is higher mortality of lower branches, reducing, consequently, crown proportion.
Results show that the relationships in trees’ size are strongly related to the growth in height, diameter and site characteristics. Such information can be helpful to build structure scenarios, to be used in morphometric and environmental models in order to define the size, stock and density of individuals (Pretzsch et al., 2014).
Determination of this variable is very important because it allows the projection of the space to be reserved for a tree to have successful development, as it grows. Knowledge of the demanded space for each individual, in turn, allows one to predict the number of individuals to be selected and conducted up to the end of rotation, when a target diameter is established.
Since the studied forests are composed by a great amount of large individuals, one can assume that it is an indication that in the initial process of forest formation, trees were able to grow fast, taking priority in the use of environmental resources (Gustafson et al., 2004). Therefore, the predominance of old individuals indicates the necessity of interventions to allow the developing of young individual, for the success of forest succession.
Positive correlation in the relationship shape-size can be regarded as an indication of trees ability to compete, since individuals will develop their crown structure faster in order to intercept light, growing higher and occupying sooner the forest canopy. Thus, crown structure is a key factor influencing competition for light and species coexistence patterns (Peltzer and Kochi, 2001).
Such growth pattern in the relationship between shape and size is expected in fertile sites, where light is the most limiting factor to tree growth. This study also shows that the competition patterns and the relationship between shape and size can spatially vary across ecological gradients and with the stage of forest development (Pretzsch and Dieler, 2010).
Trees from São Joaquim and Painel sites presented larger crown diameter and crown proportion, higher total height and higher insertion height of the crown, indicating that trees from these sites have reached maturity.
A significant result of the analysis of covariance indicate that the range of variation in shape of trees is regarded as the variation in trees size, as well as the sort of competition levels at which trees were subject in the forest throughout their development. The study of morphometric relationships with shape and size of trees may allow the identification of the need for intervention inthe forest.
Results from this work are helpful in understanding resources distribution, diametric growth in forest populations, as well as the changes in the structural dynamics with different patterns of resource availability in time.

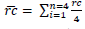 was obtained. Multiplying mean radius, crown diameter:
was obtained. Multiplying mean radius, crown diameter: 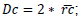 ; crown projected area:
; crown projected area: 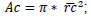 ; crown length:
; crown length: 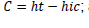 ; and crown proportion
; and crown proportion 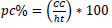 were obtained.
were obtained.