Full Length Research Paper

ABSTRACT
The objective of this study is to evaluate the interference of reduced spacing in the physiological and characteristics production components of maize hybrids (Zea mays L.). Four line spacings (95, 80, 60 and 40 cm) were tested using three commercial hybrids (H1, H2 and H3) in Senador Guiomard, Acre, Brazil. The experimental design of randomized complete blocks was used in subdivided plots. The experiment lasted four months, where the parameters measured were CO2 assimilation and gas exchange, water use and carboxylation efficiencies and productivity. Reduction in spacing did not influence the photosynthesis, with slightly differences at stomatal conductance and transpiration. However, it was observed that the H3 presented higher efficiency of water use and carboxylation at the larger spacings. Moreover, there was an increase in the number of ears and grain yield per hectare in reduced spacing, with H3 plants presenting a superior mass of 100 grains among the cultivars. It is concluded that H3 (P4285YHR) presents essential physiological characteristics favouring yields at reduced spacings compared to crops requiring larger spacing. In addition, reduced spacing lines for the maize crop do not reduce atmospheric CO2 assimilation, resulting in larger productivity per cultivated area among the three tested hybrids.
Key words: Production, photosynthesis, Zea mays.
INTRODUCTION
Maize is widely cultivated around the world, in many different soils and climate, mainly because of its large domestication. Nevertheless, its origin is from America (Werle et al., 2011). At the level of world production, Brazil takes the third place, standing behind United States and China (ABIMILHO, 2017).
Brazilian states with the largest production of maize grains in 2015/16 were Mato Grosso do Sul (9000 kg ha-1), followed by Paraná (7953 kg ha-1) and Goiás (7800 kg ha-1). At the North region, Acre state is in an intermediary position, which had a productivity of 2350 kg ha-1, in the same period (CONAB, 2017). The low productivity of Acre can be attributed to use of low-yield cultivars by the producers in addition to inadequate use of plant spacing among lines, reaching one meter (Queiroz et al., 2015).
If it is possible to increase the plant density using reductions in the plant spacing with the objective to improve grain production; thus, the whole system can be more efficient and competitive with the same planted area (Testa et al., 2016). However, high plant density of maize might increase the negative effects of drought conditions (Ferreira et al., 2014), in addition to more intraspecific competition (Sangoi et al., 2010). This rivalry among plants highly affects production amount and quality, and breaks the efficiency of utilization of environmental resources, particularly in the physiological trends associated with photosynthesis (Marenco et al., 2014).
In order to evaluate the influence of plant spacing reduction on the physiology and production of maize hybrids grown in open fields, the objective of this work is to assess physiological and production parameters of maize plants cultivated at different levels of spacing and using three maize hybrids.
MATERIALS AND METHODS
The experiment was carried out at the São Pedro Farm in 2016-17, located at Senador Guiomard, AC, Brazil (9°50,9’ S; 67°26,4’ W). The soil of the experimental area is classified as dystrophic red argisol, medium texture to clay (Rodrigues et al., 2003). Previous soil analysis performed at the 0-20 cm profile revealed the following parameters: pH = 5.4; P = 10.4 mg dm-3; K = 45.3 mg dm-3; Organic Matter = 20.6 g dm-3; Al = 0.25 cmolc dm-3; Ca = 6.5 cmolc dm-3 and Mg = 2.5 cmolc dm-3. Using the Köppen classification, the region is categorized as Am, humid and hot equatorial, with well-defined drought period, accentuated at June, July and August (Costa et al., 2012).
The experimental design was random blocks, with subdivided parcels and four replicates. Treatments were combinations of maize hybrids with different spacings. Maize hybrids were 2B 655 PW (H1); AG 7088 PRO3 (H2) e P4285 YHR (H3) while different spacings were 40, 60, 80 and 95 cm among lines, reaching a final number of 125,000; 83,333; 62,500 and 52,632 plants by hectare, respectively. The selected hybrids were in the test phase to be recommended for open field cultivation in the region. Parcels had 96 m2 of area, each one was divided in four sub-parcels of 24 m2. Plants positioned at the parcel limits were discarded from the measurements. One week before sowing, weeds in the area were controlled by using glyphosate (systemic herbicide from glycine-derivate group).
In the minimum cultivation system, seeds were sown manually, using spacing holes as explained above to set the treatments; and the line spacing was 20 cm in all treatments, using one seed per hole. Fertilizing was performed based on soil analysis and recommendations by Coelho (2006). The applications contained 60 kg P ha-1, 80 kg K ha-1 and 120 kg N ha-1 (20% basis, 35% in V4 state and 35% in V8 state) adjusting the fertilizers distribution among the different spacings. Pest and weed control were performed at December 9th of 2016 and January 21st of 2017 using glyphosate (1920 g ha-1 of the active ingredient). In order to control Deois flavopicta S., the insecticide Decis was applied, a contact product of the pyrethroid group, using 5 g ha-1 of the active ingredient.
The following gas exchange parameters were evaluated: Net photosynthesis (PN), stomatal conductance (gS), CO2 intercellular content (Ci) and leaf transpiration (E). Water use efficiency (WUE) and carboxylation efficiency (CE) were calculated by the relation between the PN and E or Ci, respectively. Gas exchange measurements were performed using an infrared gas analyzer - IRGA, LI-6400XT model (Li-Cor Inc. Lincoln, USA). These evaluations were made at the flowering stage VT, between 10 A.M. and noon, at the flag leaves of each measured plant. During the measurements, photosynthetically active photon flux density (PPFD) was maintained at 1000 µmol m-2 s-1 and the CO2 concentration in the gas analyzer chamber was 380 µmol mol-1.
Morphological characterization was performed when the plants were with the male inflorescence completely visible. The following parameters were evaluated: leaf area (cm2), plant height (m), ear insertion height (m) and stalk diameter (mm). Leaf area was evaluated measuring every photosynthetically active leaf of a plant, considering those with more than 50% of green leaf area (Borrás et al., 2003). Photosynthetically active leaf area (A) was estimated by the Equation 1:

A – leaf area (cm2);
C – length from base to the tip of the leaf (cm);
L – larger width of the leaf (cm).
At the same time, the average height of the plants, average height of the ear insertion and stalk diameter were measured. For these measurements, a ruler and pachymeter were utilized. Ten plants within each sub-parcel were evaluated. At the time of harvest (120 days after sowing), the following parameters were evaluated: total ears (ears ha-1), grains per ear (grains ear-1), mass of 100 grains (g) and grain yields (t ha-1). After the ears were harvested, the grains were weighed and water content was determined, making it possible to estimate the grain weight corrected to 13% of humidity. The results were estimated to 100 grains. Grains yield was measured and corrected to the humidity content (Equation 2):
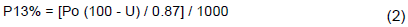
P13% - grains yield (t ha-1) corrected to the 13% humidity;
Po - [stand ha-1 x parcel yield (kg) / parcel stand];
U - grain humidity in the harvest (%).
Normality of the data was tested using a Shapiro-Wilk test and analysis of variance; and, if significant, means were compared by a Scott-Knott test (p ≤ 0.05), using ASSISTAT software (Silva and Azevedo, 2016).
RESULTS AND DISCUSSION
Concerning the CO2 assimilation and gas exchange measurements, the results were not different with respect to net photosynthesis; but they showed changes for the other evaluated parameters. The different spacing of plants or the kind of hybrids used did not affect net photosynthesis (PN) in any of the treatments. On the other hand, intercellular concentration of CO2 (Ci) was altered by hybrids and spacing. At 95 cm of spacing, H3 showed 30% less Ci compared to H1 and H2. However, when results were determined for different spacings, the Ci for H1 and H2 was not affected when the spacing was reduced; but H3 showed a larger Ci value at the 60 cm spacing (Figure 1B).
Stomatal conductance (gS) was significantly different only at the larger spacing (95 cm), with H3 plants showing lower gS in this treatment (33%) (Figure 1C). Finally, leaf transpiration (E) showed similar results to gS, with H3 presenting lower E compared with the other hybrids at 95 cm spacing, but with a slight decrease at the minor spacing as well (Figure 1D). These results were also found by Gomes et al. (2011), who found no variation among photosynthetic values in maize plants grown at 50, 70 and 90 (cm) spacings of lines. In order to maintain the photosynthesis rate when there are reductions in stomatal aperture, it has to be more efficient to absorb CO2 inside the leaf mesophyll (Lemos et al., 2012). This was observed in the H3 hybrid. Even with higher stomatal conductance and decreased CO2 concentration, phosphoenolpyruvate carboxylase (PEPC) acts efficiently, favoring maintenance of the photosynthetic potential in C4 plants such as maize (Walter et al., 2015).
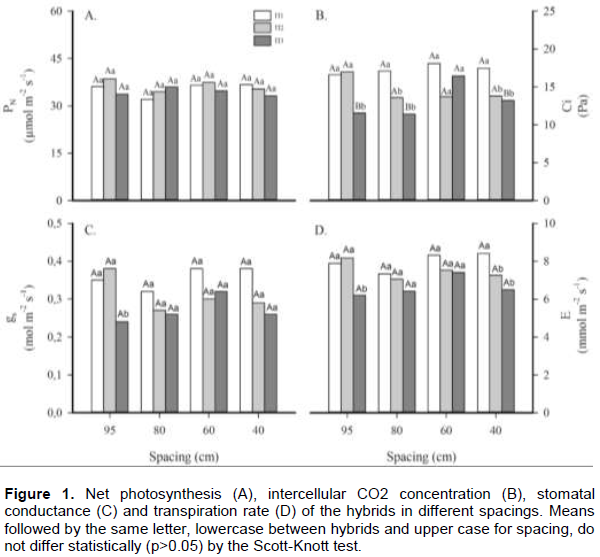
Water use and carboxylation efficiencies (WUE and CE, respectively) were calculated using collected data from PN, E and Ci. WUE was greater for H3 plants grown at the 95 and 80 cm spacings, compared with the other hybrids (Figure 2A). Probably these results are a consequence of the lower transpiration rate in these plants. However, when the spacing was reduced below 80 cm, WUE of the H3 plants was not significantly different from the other hybrids. Similarly, CE was more pronounced for H3 plants in comparison to H1 and H2 plants when grown at the 95 cm spacing, but was significantly different only in relation to H1 plants at 80 cm spacings (Figure 2B).
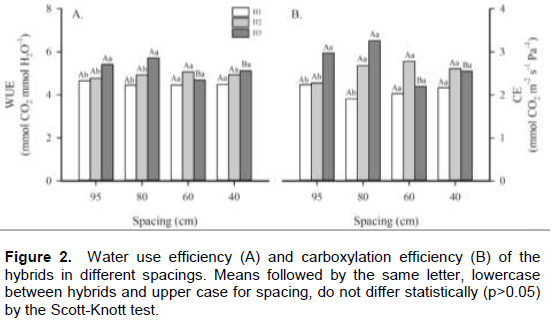
C4 plants, which contain a highly effective enzyme to assimilate atmospheric CO2 are consequently very efficient photosynthetically. However, when these plants are under stress, they reduce stomatal conductance and leaf transpiration, possibly reducing photosynthesis but increasing photosynthetic and water use efficiencies (Hartzell et al., 2018). Evaluating growth, there are some significant differences (p<0.05) among hybrids and spacing for the parameters of plant height, ear height, and stem diameter (Figure 3). However, there were no verified significant variations for leaf area. Plant hybrids achieved more than 200 cm average height. Moreover, there were statistically significant variations among hybrids in the 80 cm line spacing treatment, where H3 plants showed higher height than the other hybrids. Nevertheless, only the H1 plants showed differences in plant height for a spacing at 80 cm; with the 186.2 cm average, being lower than the height values for the other two hybrids. According to Lima et al. (2016), plant height is a characteristic that is conditioned by the plant genotype. This is similar to what we found for H2 and H3 hybrids that responded similarly in height. Calonego et al. (2011) did not find significant differences in plant height as well.
Referring to ear height, there was variation among hybrids (Figure 3B). Across all spacing treatments, H1 plants showed statistically significantly lower mean ear heights than H2 and H3, whereas the latter were similar in mean ear heights. Analysing spacing, H2 plants were the hybrid with the higher mean ear height at 60 cm, presenting 122.7 mean cm for ear height, but at 40 cm, both H2 and H3 are significantly different in mean ear height compared to H1.
According to Sangoi et al. (2010), at places with high density of plants and low line spacing, there is a high intraspecific competition to obtain light, favouring the culm elongation. This can be the reason that in this work, only the mean ear height was affected with less variation of the plant height. With regards to stem diameter, it was observed that at 95 cm spacing, hybrids were similar. But, at the 80 cm spacing, H1 showed lower stem diameter than the other hybrids. At the smaller spacings (60 and 40 cm), there was no statistically significant difference. Evaluating spacing lines, hybrid plants cultivated at the larger spacings (95 and 80 cm) showed larger mean stem diameters than the ones grown at the smaller spacings (60 and 40 cm). According to Strieder et al. (2007), plant density is the main factor that alters the stem diameter, also increasing the number of dominating plants, characterized by low development and lesser stem diameter. This situation can increase the amount of broken and bedding plants, being prejudicial especially during the mechanical harvest, with significant losses of the maize crop (Rezende et al., 2015).
In the productivity components, there were differences (p<0.05) only for the productivity and ear by hectare parameters among the different spacings (Figure 4). Nevertheless, the number of kernels by ear was variable among hybrids and spacing while the 100 grains weight was different only among hybrids. Productivity varied from 7.53 to 12.81 t ha-1, increasing while the spacing between lines was reduced. It is important to notice that H2 and H3 hybrid plants were more productive (approximately 80%) at the smaller spacings, compared with the larger spacing lines. Lima et al. (2016) observed that yield increased with the increment of the plants number by area. Similar results were found by Lana et al. (2009) and Modolo et al. (2010), who found more kernel productivity at reduced spacing.
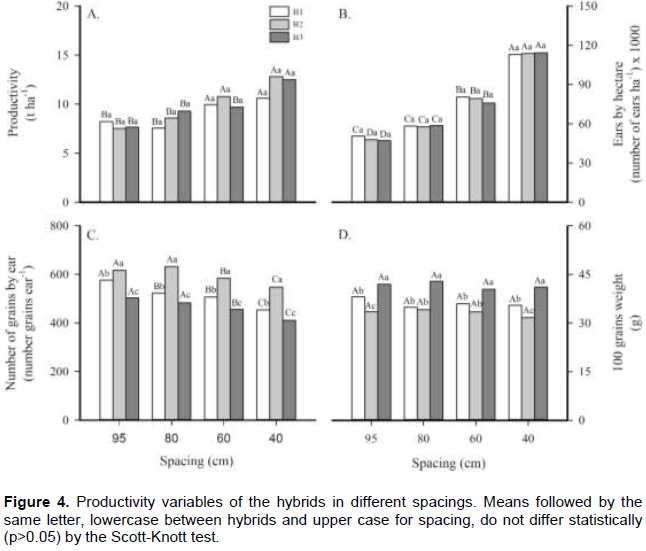
Checking the number of ears per hectare, it can be seen that hybrids did not vary at the same spacing. Nevertheless, examining only the different spacing lines, there was a 2-fold increase in mean ear number per hectare comparing the data for the smaller spacing (40 cm) with the larger one (95 cm). Serpa et al. (2012) verified that the ear number per square meter was linearly influenced by the plant density. Still, they reported that an increase in the plant’s density can cause female sterility, directly influencing the number of fertile ears per area.
The number of grains by ear was higher for the H2 hybrid, with a larger average at all spacings. Evaluating the number of grains per ear in relation to the spacing treatments, this parameter was reduced concomitantly with reduced spacing lines, independently of hybrid treatments. According to Testa et al. (2016), at reduced spacing lines and higher plant’s density, there is a reduction of the ear size, consequently reducing the grain amounts and weights. However, these same authors report that the key to increased productivity with high density of plants is the higher number of harvested grains per unit of area. The different spacing lines had no significant impact on the 100-grains weight (Figure 4D). Though, it was observed that H3 plants showed a higher average for this parameter at all spacing lines, mainly at the treatments with more density of plants. However, Li et al. (2015) reported different results when planting with higher densities. They found grains with lighter and smaller cores, reducing the plants’ productive potential.
CONCLUSIONS
(1) The hybrid P4285 YHR (H3) showed higher water use and carboxylation efficiencies at the smaller spacing lines, because they presented lower stomatal conductance, while keeping similar values of photosynthesis compared to the other tested hybrids.
(2) Reduced spacing lines of the maize crop do not reduce atmospheric CO2 assimilation, resulting in larger productivity per cultivated area among the three tested hybrids.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Associação Brasileira das Indústrias do Milho (ABIMILHO) (2017). 8º Acompanhamento da Safra de Milho Total - 2016/17. Disponível em: |
|
|
Borrás L, Maddonni GA, Otegui ME (2003). Leaf senescence in maize hybrids: plant population, row spacing and kernel set effects. Field Crops Research 82:13-26. |
|
|
Calonego JC, Poleto LC, Domingues FN, Tiritan CS (2011). Produtividade e crescimento de milho em diferentes arranjos de plantas. Revista Agrarian 4:84-90. |
|
|
Coelho AM (2006). Nutrição e Adubação do Milho. Sete Lagoas/Centro Nacional de Pesquisa de Milho e Sorgo. 10 p. |
|
|
Companhia Nacional de Abastecimento (CONAB) (2017). Acompanhamento de safra brasileira: Safra 2016/17 de grãos. 3:144 p. |
|
|
Costa FS, Amaral EF, Butzke AG, Nascimento SS (2012). Inventário de emissões antrópicas e sumidouros de gases de efeito estufa do estado do Acre: ano base 2010. Rio Branco/Embrapa Acre 144 p. |
|
|
Ferreira G, Alfonso M, Depino S, Alessandri E (2014). Effect of planting density on nutritional quality of green-chopped corn for silage. Journal of Dairy Science 97:5918-5921. |
|
|
Gomes KR, Amorim AV, Ferreira FJ, Filho FLA, Lacerda CF, Gomes Filho E (2011). Respostas de crescimento e fisiologia do milho submetido a estresse salino com diferentes espaçamentos de cultivo. Revista Brasileira de Engenharia Agrícola e Ambiental 15:375-370. |
|
|
Hartzell S, Bartlett MS, Porporato A (2018). Unified representation of the C3, C4, and CAM photosynthetic pathways with the Photo3 model. Ecological Modelling 38:173-187. |
|
|
Lana MC, Woytichoski Junior PP, Braccini AL, Scapim CA, Avila MR, Albrecht LP (2009). Arranjo espacial e adubação nitrogenada em cobertura na cultura do milho. Acta Scientiarum Agronomy 31:433-438. |
|
|
Leakey ADB, Ainsworth EA, Bernacchi CJ, Alistair R, Long SP, Ort DR (2009). Elevated CO2 effects on plant carbon, nitrogen, and water relations: six important lessons from FACE. Journal of Experimental Botany 60:2859-2876. |
|
|
Lemos JP, Galvão JCC, Silva AA, Fontanetti A, Lemos LMC (2012). Efeito da roçada das espécies Bidens pilosa e Commelina benghalensis nas características morfológicas do milho. Revista Brasileira de Agropecuária Sustentável 2:32-40. |
|
|
Li J, Xie RZ, Wang KR, Ming B, Guo YQ, Zhang GQ, Li SK (2015). Variations in maize dry matter, harvest index, and grain yield with plant density. Agronomy Journal 107:829-834. |
|
|
Lima SF, Alvarez RCF, Contardi LM (2016). Influência do espaçamento entre linhas em características fitotécnicas e acúmulo de massa seca de híbridos de milho. Ambiência 12:1027-1039. |
|
|
Marenco RA, Vera SAA, Gouvêa PRS, Camargo MAB, Oliveira MF, Santos JKS (2014). Fisiologia de espécies florestais da Amazônia: fotossíntese, respiração e relações hídricas. Revista Ceres 61:786-799. |
|
|
Modolo AJ, Carnieletto R, Kolling EM, Trogello E, Sgarbossa M (2010). Desempenho de híbridos de milho na Região Sudoeste do Paraná sob diferentes espaçamentos entre linhas. Revista Ciência Agronômica 41:435-441. |
|
|
Queiroz LR, Costa FS, Oliveira TK, Marinho JTS (2015). Aspectos da cultura do milho no Estado do Acre. Rio Branco/Embrapa Acre 30 p. |
|
|
Rezende WS, Brito CH, Brandão AM, Franco CJF, Ferreira MV, Ferreira AS (2015). Desenvolvimento e produtividade de grãos de milho submetido a níveis de desfolha. Pesquisa Agropecuária Brasileira 50:203-209. |
|
|
Rodrigues TE, Gama JRNF, Silva JML, Valente MA, Santos ES, Rolim PAM (2003). Caracterização e classificação de solos do município de Senador Guiomard, Estado do Acre. Embrapa Amazônia Oriental 69 p. |
|
|
Sangoi L, Silva PRF, Argenta G, Rambo L (2010). Ecofisiologia da cultura do milho para altos rendimentos. Lages: Graphel. 87 p. |
|
|
Serpa MS, Silva PRF, Sangoi L, Vieira VM, Marchesi DR (2012). Densidade de plantas em híbridos de milho semeados no final do inverno em ambientes irrigados e de sequeiro. Pesquisa Agropecuária Brasileira 47:541-549. |
|
|
Silva FAZ, Azevedo CAV (2016). The Assistat Software Version 7.7 and its use in the analysis of experimental data. African Journal Agricultural Research 11:3733-3740. |
|
|
Strieder ML, Silva PRF, Argenta G, Rambo L, Sangoi L, Silva AA, Endrigo PCA (2007). Resposta do milho irrigado ao espaçamento entrelinhas depende do hibrido e da densidade de plantas. Revista Ciência Rural 37:634-642. |
|
|
Testa G, Reyneri A, Blandino M (2016). Maize grain yield enhacement trough high plant density cultivation with different inter-row spacings. European Journal of Agronomy 72:28-37. |
|
|
Walter LC, Rosa HT, Streck NA (2015). Acclimation mechanisms to elevated CO2 in plants. Ciência Rural 45:1564-1571. |
|
|
Werle AJK, Nicolay RJ, Santos RF, Secco D (2011) Avaliação de híbridos de milho convencional e transgênico (Bt), com diferentes aplicações de inseticida em cultivo safrinha. Revista Brasileira de Tecnologia Aplicada nas Ciências Agrárias 4:150-159. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


