ABSTRACT
Anther culture is one of the biotechnology tools using for quick recovery of fixed breeding pure lines in rice. The development of new rice varieties is highly dependent on genetic diversity in desirable agronomic traits. Therefore, this study aimed to identify potential genotypes having the characters of Tongil-type and to study suitable traits of tested Double Haploid (DH) lines which can be applied in breeding program. Twenty advanced DH lines along with one local check were planted in a randomized complete block design with three replications during autumn seasons of 2017 and 2018. All evaluated genotypes exhibited a wide and significant variation in the twelve measured traits. The highest heritability related with high genetic advance was recorded for the number of tillerplant-1, biomass yield, plant height, and grain yield. Genotypic coefficient of variation and genetic advance were recorded for number of tillersplant-1 and plant height in both seasons. Moreover, there was a highly significant and positive correlation of grain yield with number of panicle per m2 (0.76), number of filled grain per panicle (0.71), number of tillersplant-1 (0.69), panicle weight (0.45) and biological yield (0.45). Based on productive tillers number, number of filled grainspanicle-1, number of panicle per m2, biological yield and grain yield, the lines KF323819, KF323895, KF323826, KF323893, KF323847, KF323855 and KF323885 were high potential could be considered as a new type of aerobic rice in Sudan. In additions, hybridization of these high yielding could be used to achieve higher heterosis among the genotypes and developing reliable selection indices for improvement rice breeding programs.
Key words: Double haploid, anther culture, yield components, broad sense heritability and path coefficient analysis.
Rice (Oryza sativa L.) is the world’s leading cereal crop as more than half of the world’s population is dependent
on it as their staple food (Bashir et al., 2010). The rising world population is increasing rapidly; rice production needs to be increased to meet its demands in the coming years in order to keep pace with increasing population mainly in Asia and Sub-Saharan Africa (Anyaoha et al., 2018). It is estimated that a 50% increase in rice grain yield may be required by 2050 to keep hunger away (Pang et al., 2017). This important cereal is cultivated and consumed in White Nile State in the Sudan but its production is characterized by poor yields resulting from use of low farm inputs and cultivation of unimproved cultivars with poor yield potentials (Osman et al., 2012). Hence, improving rice yield potential or its yield under various conditions is the foremost task for rice breeders (Ali et al., 2017).
The most sustainable way to increase rice production is via genetic and breeding improvements (Fischer, 2015). Perhaps even more importantly, rice breeding enables the integration of novel traits, which is essential in achieving yield stability in the changing climatic conditions we are facing (Dawson et al., 2015). The challenge for contemporary plant breeding is not only to integrate new traits into our crops, but to accelerate the genetic gain of its breeding programs at the same time, in order to achieve a doubling in speed of yield increase (Dwivedi et al., 2015).
Anther culture is a tissue culture procedure which can be applied in a breeding in order to accelerate the process of gaining pure lines (Herawati et al., 2010). With the development of doubled haploid (DH) techniques on improving the speed of genetic gain when applied in breeding programs, as well as their importance and diverse applicability in basic and applied research (Roy and Mandal, 2005). Anther culture breeding system which can shorten varietal development cycles by 3~5 years allow breeders to improve existing local varieties. DH techniques have been, and are being, used to accelerate the breeding programs of a range of crops, most notably maize, wheat, rice and barley (Seguí-Simarro, 2015).
Rice researchers in South Korea have made breakthrough in yield potential that occurred from modifying the existing high yield plant type to improve biomass, harvest index, reaching 7-8 tons/ha in polished rice. This occurred by formulating prototypes of semi-dwarf rice cultivars called ‘Tongil-type’ (TGT) through wide crosses between indica and japonica cultivars using Anther culture techniques. The desired characters of the TGT rice are compact growth, good number of productive tillers, big panicles and good grain filling (120-200 grains), semi dwarf (80-100 cm in height), erect and thick leaves with dark green colour, medium earliness (100-130 days), deep rooting system, soft and sticky endosperm and high resistance to major pests and diseases (Khush and Brar, 2002). Recently, a huge DH progenies were obtained from interspecific crosses between Tongil-type and Japonica with African varieties and wild species including Oryza glaberrima, Oryza longistaminat and Oryza rufipogon. Many of them were distributed to different African countries in order to determine the agronomic character stabilities for further selection of desirable characters, particularly in terms of yield components and yield. This will be serving as good resources for improving yield potential of local varieties particularly in the Sudan.
The magnitude of genetic variability provides useful information with regard to the possibility and extent of improvement that may be expected in the characters through breeding and selection. Breeders are interested in evaluating genetic diversity based on morphological characteristics as they are inexpensive, rapid, and simple to score. Moreover, this evaluation could be useful in developing reliable selection indices for important agronomic traits in rice. Therefore, the main objective of the present investigation was to assess the existing genetic variability and to select DH lines of aerobic rice having the characters of the Tongil-type and japonica to establish relationship in yields and its components and utilization of the available population in future rice breeding programs.
Study site and experimental design
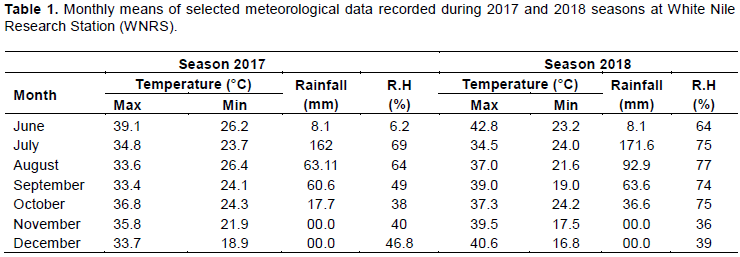
A total of 1027 independent lines, each containing one or a few plantlets that were regenerated from separated calli, were produced through anther culture of the (F2 – F3) generations derived for 14 interspecific crosses among Korean Tongil-type (Semi-dwarfism, high-yielding potential, earliness, medium amylose and medium-long grain,) and Japonica (Earliness, low-medium amylose, soft and sticky endosperm) with African varieties and wild species including O. glaberrima, O. longistaminata, and O. rufipogon (high biomass, disease resistance and low yield). They were established at Tissue Culture Laboratory Africa-Rice St. Louis. Of these 597 double Haploid lines were grown at Sahel Station in observation yield trial (OYT) then after several series of selection and fixation, in advanced generation,105 potential lines with the required adaptive traits were received and conducted in OYT; then again selected the superior 20 DH lines along with one check namely Kosti-1, were grown for two consecutive seasons (2017 and 2018) during the rainy season under irrigated condition; at White Nile Research Station Farm (latitude 14° 24 N and longitude 33° 22 E), Kosti of the Agricultural Research Corporation (ARC) Sudan. The soil of the experimental plots was classified as vertisol with high clay content (40 to 65%), less than 1% organic carbon, low in available nitrogen (0.03% total nitrogen) and medium in available P2O5 (406 to 700 ppm total phosphorus); pH values is slightly alkaline ranging from 7 to 8.2. The climate was semi-arid (Table 1).
The experimental plots were laid out in randomized complete block design (RCBD) with three replications. In both seasons deep plough, harrowing and leveling were practiced to prepare experimental area. The seeds were drilled on July 7th and 5th of 2017 and 2018 in turn, using a seed rate of 80 kg /ha. The plot size was 3 × 6 m with 0.2 m apart consisting of 15 rows giving a total area of 18 m2. Recommended fertilizers of Urea and Triple Super Phosphate (TSP) at the rate of 129 kg N ha-1 and 43 kg P2O5 ha-1, respectively were used. TSP was applied as basal dose during final land preparation. Nitrogen was top dressed in two equal split doses at 21 days after sowing and the other before panicle initiation. Hand weeding was performed three times per season. All plots were irrigated at sowing and then at weekly intervals until it reached the flowering stage and then every 3 days. All other agronomic practices were applied as per the recommendation for rice production in the two seasons to raise a healthy rice crop.
Data collection
Data were collected on days to heading (days from sowing to the time when the 50% of the plants were headed) and days to maturity (days from sowing to the time when 80% of the panicles reached full maturity). Then at harvest, plant height (cm), panicle length (cm), panicle weight (g), number of tillersplant-1, number of filled grainspanicle-1, percent of unfilled grains panicle-1, 1000 grain weight (g), grain yield (t/ha) and Biological yield (t/ha) based on grain yield per plot were recorded. The data were collected according to standard evaluation systems for rice standard evaluation system (SES) (IRRI. 2002); ten randomly selected plants in the net harvested area of each plot were used for data collection.
Statistical analysis
Analysis of variance (ANOVA) was carried out on the data to assess the genotypic effects and their interaction using general linear model (GLM) procedure for randomized complete blocks design in SAS (version 9.1). Estimates of variance components were generated and combined analysis of variance was done for the traits in which the mean squares were homogenous. The phenotypic and genotypic variances and their coefficients, heritability in the broad sense and genetic advance were computed according to the formula described by Singh and Chaudhary (1985). Means for each season were used to compute simple linear correlation coefficients. Path coefficient analysis was used to partition the simple linear correlation coefficients, combined over seasons, between grain yields/ha and four other traits; namely number of panicle per m2, number of filled grainspanicle-1, number of tillersplant-1 and plant height (cm) at maturity, into the direct and indirect effects.
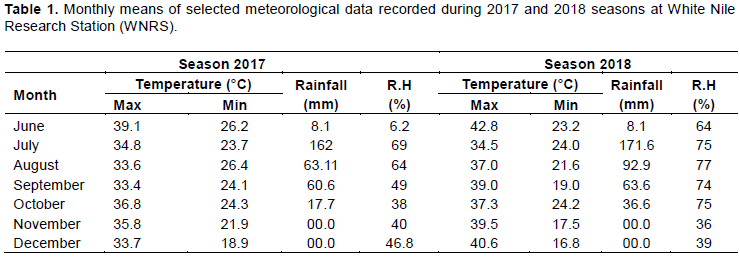
The results showed that there is a presence of acceptable amount of variability among the genotypes. This gives an opportunity for rice breeders to improve those traits through selection and hybridization to improve the desired traits. The range and mean of genotypes for all studied traits also indicated wide ranges of variation which also revealed possible amount of variability among the genotypes (Table 2).
The broad sense heritability is the relative magnitude of genotypic and phenotypic variances for the traits and it is used as a predictive role in selection procedures. This gives an idea of the total variation ascribable to genotypic effects, which are exploitable portion of variation. The characters that showed relatively high heritability estimates (≥ 90%) were number of panicle/m2 (98.1 and 97.5), panicle length (97.1 and 98.1), number of tiller/plant (95.5 and 93.5), 1000 grain weight (95.3 and 93.7), number of unfilled grain/panicle (93.4 and 94.0) and plant height (91.5 and 93.7) in both seasons respectively, indicating the optimizing of homogeneous of these lines and their improvement could be achieved by mass selection. Similar results were reported by Sohrabi et al. (2012). This indicated that selection of these traits would be more effective as compared to others. The moderate heritability estimate for grain yield was attributed to the fact that yield is a complex trait and is controlled by many genes. Since high heritability does not always indicate high genetic gain. Heritability with genetic advance considered together should be used in predicting the ultimate effect for selecting superior varieties (Anyaoha et al., 2018; Ahmed et al., 2018). High genetic advance and heritability were recorded for the number of tiller/plant, number of unfilled grain/panicle, biological yield t/ha, plant height, and grain yield t/ha. These results suggested that these traits were primarily under genetic control and selection for these traits can be achieved through their phenotypic performance. High heritability estimates with low genetic advance observed for days to heading, days to maturity and panicle weight indicated non additive type of gene action and that genotype × environment interaction played a significant role in the expression of the traits.
The genetic analysis of quantitative traits is a prerequisite for plant breeding programs, which can lead to a systemic method of design and to the appropriate planning of plant breeding strategies. The current study suggests that the packed cell volume (PCV) was higher than the Genotypic Coefficient of Variability (GCV) for all traits. This was also the case for all the traits observed in another study; Osman et al. (2012) which reported that the environmental effect on any trait is indicated by the magnitude of the differences between the genotypic and phenotypic coefficients of variation. Large differences reflect a large environmental effect, whereas small differences reveal a high genetic influence. In this study, the small differences between the PCV and GCV for most of the traits, such as days to heading, plant height, panicle length, number of unfilled grain per panicle, 1000 grain weight and grain yield t/ha, represented some degree of environmental influence on the phenotypic expression of these characters. It also suggests that selection based on these characters would be effective for future crossing programmes. The other traits, which showed a higher difference between PCV and GCV, indicated that the environmental effect on the expression of those traits is higher and that selection based on these characters independently is not effective for further yield improvement. The highest PCV recorded for number of unfilled grain/panicle (34.8-38.9) and numbers of tiller/plant (24.5-25.7) in both seasons were also recorded by the following researchers (Mulugeta et al., 2011; Abebe et al., 2017).
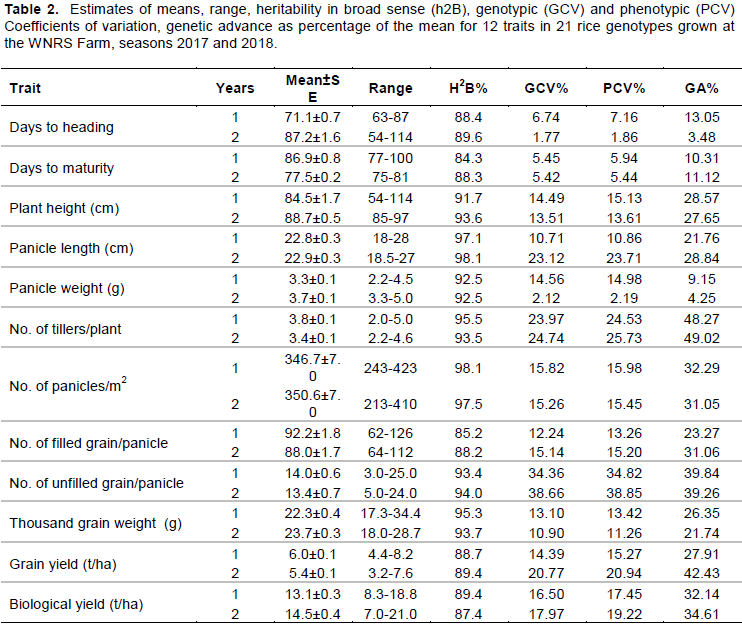
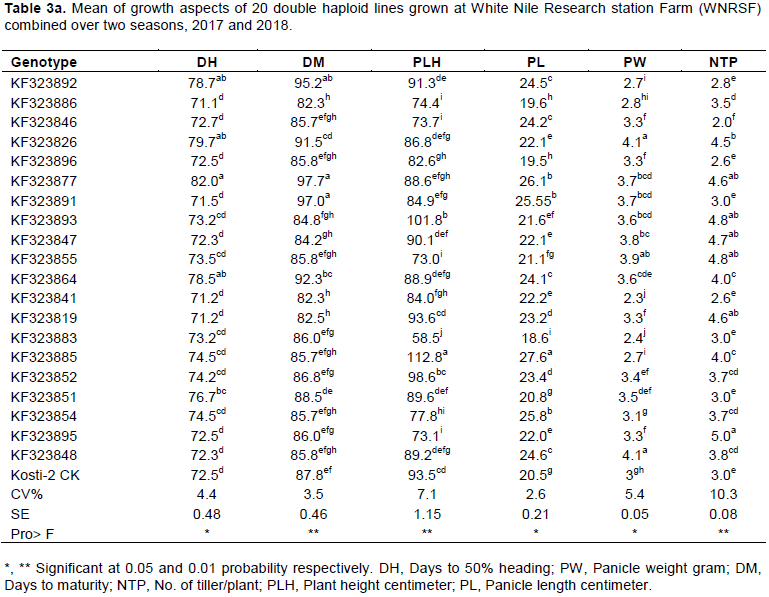
The results further revealed that most of the traits exhibited wide range of variability (Table 3a-b). The late heading and maturing genotypes were KF323877 and KF323892, while the early flowering and maturing genotype was KF323886, KF323841 and KF323819; as compared to the local check variety (Kosti-1) most of the genotypes showed extra early maturity period. However, only four genotypes were late maturing than the local check Kosti-1. This suggested the chance of selecting earliness genotypes which can escape terminal moisture and drought stress. The range for plant height was 58.5 to 112.8 cm with genotype KF323885 as the tallest and genotypes KF323883 as the shortest. According to IRRI irrigated rice plant height is classified as semi-dwarf (less than 110 cm), intermediate (110-130 cm) and tall (more than 130 cm). Based on this classification, in the present study 90.4% of the tested genotypes group are under the semi-dwarf class. This indicated that the tested genotypes had inherent variability in stature to develop lodging resistant varieties (semi-dwarf) that will have higher response to nitrogen application. Also there is reported variation in plant height by Akinwale et al. (2011), Atif and Khalid (2013) and Abebe et al. (2017). The local check panicle length recorded was 20.5cm with maximum value of 27.6 cm and minimum value of 18.6 cm for KF323885 and KF323883, respectively. Based on the IRRI irrigated rice classified argument, the present finding showed that there is enough medium variability for panicle length among the genotypes for improving panicle architecture and grain yield due to high association of this trait that determines the number of grains it can hold (Table 3a).
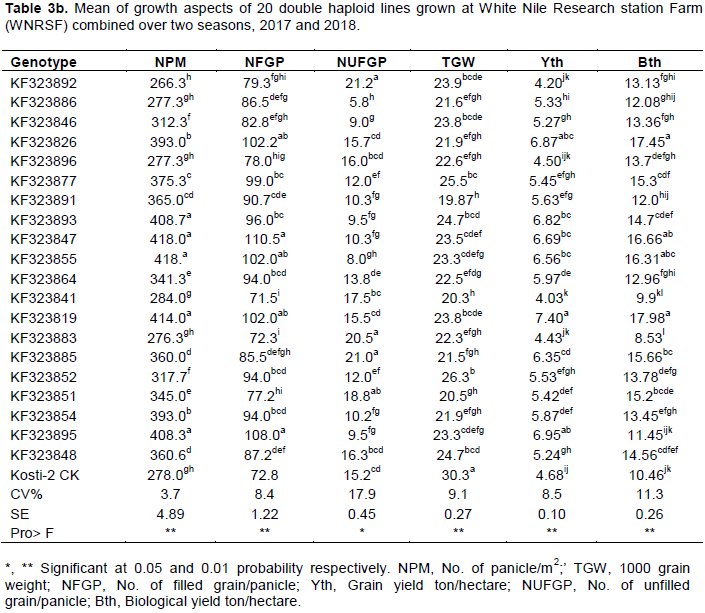
The range for number of filled grainspanicle-1 was 71.5 to 110.5 for genotypes KF323841 and KF323847, respectively. The range of percentage of unfilled grainspanicle-1 was 5.8 to 21.2 for genotypes KF323886 and KF323892 respectively (Table 3b). Adequate number of fertile grains/panicle and heavy grains are important traits, which should be considered in selection for high yield genotypes (Osman et al., 2012; Abebe et al., 2017). Relatively range performance was revealed among the studied traits, grain yield (4.03-7.40 t/ha) with a local check grain yield of 4.68 t/ha. The genotypes significantly varied for grain yield and about 76.2% of the genotypes had higher grain yield than the standard check (Kosti-1). Among the genotypes, KF323819, KF323895, KF323826, KF323893, KF323847, KF323855 and KF323885 were the top yielders with corresponding grain yield of 7.40, 6.95, 6.87, 6.82, 6.69, 6.56 and 6.35 t/ha, respectively. The local check (Kosti-1) was the medium yielder (4.68) (Table 2b). Thus, the existence of enough variability among genotypes was highly significant. Hence, it offers a better scope for further selection of promising DH genotypes in rice breeding program particularly selection based on yield is reliable.
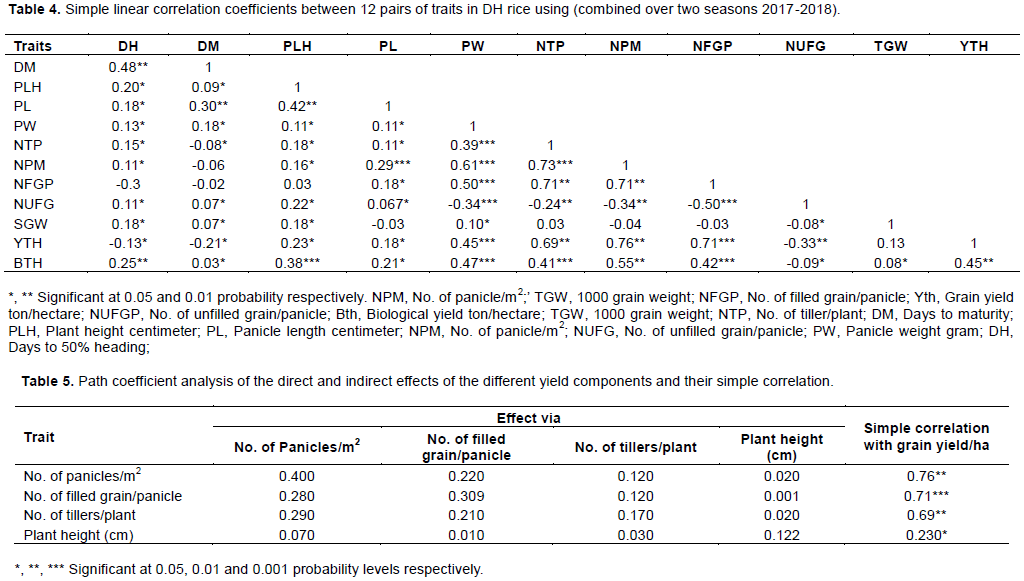
Yield is a complex product being influenced by several independent quantitative characters. Breeders always look for variation among traits to select desirable types. Some of these characters are highly associated among themselves and with seed yield. The analysis of the correlations among these characters and their associations with grain yield is essential to establish selection criteria. When more characters are involved in correlation study it becomes difficult to ascertain the characters which really contribute toward yield. The relationships existing between 12 quantitative traits represented as simple correlation coefficients are presented in Table 4. Correlation of yield and other traits is important in indirect selection for high yield improvement in crop genotypes (Idris et al., 2012). There was a highly significant and positive correlation of grain yield with number of panicle per m2 (0.76), number of filled grain per panicle (0.71), number of tillers per plant (0.69), biological yield (0.45) panicle weight (0.45), plant height (0.23) and panicle length (0.18) (Table 4). Conversely, grain yield exhibited negative significant correlation with number of unfilled grain per panicle (-0.33), days to 50% heading (-0.13), and days to maturity (-0.21). The significant and positive correlation with grain yield is a strong indication that these traits are major factors in improving grain yield; also suggests that selection directed towards these characters will be effective in ensuring high grain yield in Double haploid rice. These results collaborate with the finding of Ogunbayo et al. (2014) who observed a positive and significant correlation between grain yield and number of filled gain per panicle. Also these results were in agreement with that reported by Kato et al. (2008). The negative correlation obtained between days to heading and days to maturity and grain yield indicate that grain yield can be improved by selecting early maturing genotypes.
Partitioning of yield and yield component into direct and indirect effect revealed that number of panicle per m2 had the highest direct positive contribution (0.400) to the grain yield t/ha and also had positive indirect effect on grain yield t/ha through number of filed grain/panicle (0.220), number of tillers/plant (0.120) and plant height (0.020) (Table 4). Many research workers reported similar findings (Surek and Beser, 2003; Abdulfiyaz et al., 2011; Sanni et al., 2012). The direct effect of number of filled grains/panicle on grain yield was the second largest (0.309); it had positive indirect effect on grain yield through number of panicle per m2 (0.280) and number of tillers/plant (0.120) and plant height (0.001). The direct effect of number of tillers/plant on grain yield t/ha was the third largest (0.170), and its indirect effect through number of panicle per m2 (0.290) and number of filled grain/panicle (0.210). The direct effect of plant height on grain yield was the fourth largest (0.122). It had positive indirect effect on grain yield through number of tillers/plant (0.030), number of filled grains/panicle (0.010) and number of panicle per m2 (0.070). indicate that both correlation and path coefficient analysis suggested these four traits should be considered in further selection procedures for higher grain yield.
In conclusion, the present study identified the presence of adequate genetic variability among 21 tested genotypes. Hence, the information generated from this study, rice breeder can be exploited for future rice breeding program. The study was also carried out for one sit and two seasons. Therefore, it is advisable to select and grow the highest yielding genotype (KF323819, KF323895,KF323826, KF323893, KF323847, KF323855 and KF323885) at least in more than three locations (National Yield Trial); considering. major rice growing areas to make sound recommendations for release Moreover, it is recommended that future rice research should explore molecular means to further confirm the outcome of DH lines study findings, in order to do an efficient selection process.
The authors have not declared any conflict of interests.
The authors gratefully appreciate Rural Development Administration of Korea (RAD) for funding the research work and Korea-Africa Food and Agriculture Cooperation Initiative (KAFACI) for providing the research material through Africa Rice Center. White Nile Research Station of Agricultural Research Corporation (ARC) Sudan is also appreciated for hosting the study. Special recognition is forwarded to Dr. Kang Kyung-ho for giving the opportunity to select the superior DH materials from his research field to make breakthrough in Sudan. Finally, thanks to the research team for significant time and effort spent for data collection and field management.
REFERENCES
|
AbdulFiyaz R, Ramya KT, Chikkalingaiah1, Ajay BC, Gireesh C, Kulkarni RS (2011). Genetic variability, correlation and path coefficient analysis studies in rice (Oryza sativa L.) under alkaline soil condition. Electronic Journal of Plant Breeding 2(4):531-53.
|
|
|
|
Abebe T, Alamerew S, Tulu L (2017). Genetic Variability, Heritability and Genetic Advance for Yield and its Related Traits in Rainfed Lowland Rice (Oryza sativa L.) Genotypes at Fogera and Pawe, Ethiopia. Advance Crop Sciences 5(2):1-8.
Crossref
|
|
|
|
|
Ahmed T, Kumar A, Pandey D, Prasad B (2018). Correlation and path coefficient analysis for yield and its attributing traits in bread wheat (Triticum aestivum L.). Journal of Applied and Natural Science 10(4):1078-1084.
Crossref
|
|
|
|
|
Akinwale M, Gregorio G, Nwilene F, Akinyele B, Ogun-bayo SA, Odiyi AC (2011). Heritability and correlation coefficient analysis for yield and its components in rice (Oryza sativa L.). African Journal of Plant Science 5(3):207-212.
|
|
|
|
|
Ali J, Xu JL, Gao YM, Ma XF, Meng LJ, Wang Y, Pang YL, Guan YS, Xu MR, Revilleza JE, Franje NJ, Zhou SC, Li ZK (2017). Harnessing the hidden genetic diversity for improving multiple abiotic stress tolerance in rice (Oryza sativa L.). PLoS ONE 12(3):e0172515.
Crossref
|
|
|
|
|
Anyaoha C, Felix A, Uyokei U, Bosede P, Vernon G, Semon M, Mamadou F (2018). Genetic Diversity of Selected Upland Rice Genotypes (Oryza sativa L.) for Grain Yield and Related Traits. International Journal of Plant and Soil Science 22(5):1-9.
Crossref
|
|
|
|
|
Atif EI, Khalid AM (2013). Estimation of Genetic Variability and Correlation for Grain Yield Components in Rice (Oryza sativa L.). Global Journal of Plant Ecophysiology 3(1):1-6.
|
|
|
|
|
Bashir MU, Akber N, Iqbal A, Zaman H (2010). Effect of different sowing dates on yield and yield components of direct seeded coarse rice (Oryza sativa L.) Pakistan Journal of Agricultural Science 47(4):361-365.
|
|
|
|
|
Dawson IK, Russell J, Powell W, Steffenson B, Thomas WTB, Waugh R, BarleyA (2015). Translational model for adaptation to climate change.New Phytologist 206(3):913-931.
Crossref
|
|
|
|
|
Dwivedi SL, Britt AB, Tripathi L, Sharma S, Upadhyaya HD, Ortiz R (2015). Haploids: Constraints and opportunities in plant breeding. Biotechnology Advances 33(6):812-829.
Crossref
|
|
|
|
|
Fischer RA (2015). Definitions and determination of crop yield, yield gaps, and of rates of change. Field Crops Research 182:9-18.
Crossref
|
|
|
|
|
Herawati R, Bambang S, Purwok, Iswari SD (2010). Characterization of Doubled Haploid Derived from Anther Culture for New Type Upland Rice. Indonesian Journal of Agronomy 38(3):170-176.
|
|
|
|
|
Idris AE, Justin FJ, Dagash YMI, Abuali AI (2012). Genetic Variability and Inter Relationship between Yield and Yield Components in some Rice Genotypes. American Journal of Experimental Agriculture 2(2):233-239.
Crossref
|
|
|
|
|
IRRI (2002). Standard Evaluation System for rice (SES). International Rice Research Institute (IRRI), Manila, November 2002.
|
|
|
|
|
Kato Y, Kamoshita A, Yamagishi J (2008). Preflowering abortion reduces spikelet number in upland rice (Oryza sativa L.) under water stress. Crop Science 48:2389-2395.
Crossref
|
|
|
|
|
Khush GS, Brar DS (2002). Biotechnology for rice breeding: progress and impact. In: Sustainable rice production for food security. Proceedings of 20th Session Rice Communications, pp. 23-26.
|
|
|
|
|
Mulugeta S, Sentayehu A, Kassahun B (2011). Evaluation of upland NERICA rice (Oryza sativa L.) genotype for grain yield and yield components along an altitude gradient in Southwest Ethiopia. Journal of Agronomy 10(4):105-111. ISSN 1812-5379.
Crossref
|
|
|
|
|
Ogunbayo SA, Sie M, Ojo DK, Sanni KA, Akinwale MG, Toulou B, Shuttu A, Idehen EO, Popoola AR, Daniel IO, Gregorio GB (2014). Genetic variation and heritability of yield and related traits in promising rice genotypes (Oryza sativa L). Journal of Plant Breeding and Crop Science 6(11):153-159.
|
|
|
|
|
Osman KA, Mustafa AM, Ali A, Yonglain Z, Fazhan Q (2012). Genetic variability for yield and related attributes of upland rice genotypes in semi-arid zone (Sudan). African Journal of Agricultural Research 7(33):4613-4619.
Crossref
|
|
|
|
|
Pang Y, Chen K, Wang X (2017).Recurrent selection breeding by dominant male sterility for multiple abiotic stresses tolerant rice cultivars. Euphytica 213:268.
Crossref
|
|
|
|
|
Roy B, Mandal BA (2005). Anther culture response in indica and variations in major agronomic characters among the androclones of a scented cultivar, Karnal local. African Journal of Biotechnology 4(2):235-240.
|
|
|
|
|
Sanni KA, Fawole I, Ogunbayo SA, Tia DD, Somado EA, Futakuchi K, Sie M, Nwilen FE, Guei RG (2012). Multivariate analysis of diversity of landrace rice germplasm. Crop Science 52(2):494-504.
Crossref
|
|
|
|
|
SAS Institute (2000). SAS Linear Model: A Guide to ANOVA and GLM Procedures. SAS Inst. Inc., Cary, NC, USA.
|
|
|
|
|
Seguí-Simarro JM (2015). Editorial: Doubled haploidy in model and recalcitrant species. Front. Plant Sciences 6:1-2.
Crossref
|
|
|
|
|
Singh RK, Chaudhary BD (1985). Biometrical Methods in Quantitative Genetic Analysis. Third edition. Kalyani Publisher, New Delhi.
|
|
|
|
|
Sohrabi M, Rafii MY, Hanafi MM, Akmar ASN, Latif MA (2012). Genetic diversity of upland rice germplasm in Malaysia based on quantitative traits. The Scientific World Journal, Article ID: 416291.
Crossref
|
|
|
|
|
Surek H, Beser N (2003). Correlation and Path Coefficient Analysis for Some Yield_Related triats in Rice (Oryza Sativa L.). Turkish Journal of Agriculture and Forestry 27(2):77-83.
|
|