Full Length Research Paper

ABSTRACT
To study the possible use of fungi to control Varroa destructor Anderson & Trueman, a parasitic mite of Apis mellifera L., without effects on its pollinator’s behavior, the entomopathogenic fungi Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae (Metschnikoff) were applied at 30ºC in the dark, at 108 conidia/mL, a concentration known previously for its pathogenicity on V. destructor. Both fungi did not affect diverse parameters of the normal behavior of nursing honey bees in the laboratory, including the duration of walking periods, antennae tapping, resting, feeding, drinking, communication with the antennae (touching other bee), and grooming, as observed using a PC with the Observer® software. These results indicate that it would be possible to use both fungi against the mite, without any side effects on the bee. However, as pathogenicity varies greatly in diverse strains, further studies are recommended.
Key words: Beauveria bassiana, hives, honey bees, Metarhizium anisopliae, observer, varroasis.
INTRODUCTION
Severe infestation by Varroa destructor, the main parasitic mite of the honey bee can lead to reduction in bee population, survival and overall hive productivity, and if not controlled, leads to the collapse of the colony (Sima, 2013). This mite feeds the haemolymph and debilitates the larvae, pupae, and adults (Sammataro et al., 2000; Chandler et al., 2001; Kanga et al., 2003; Márquez et al., 2003), which also emerge deformed (De Jong, 1997; Shaw et al., 2002). Although the bees remove the infested larvae and pupae, the hive weakens, and as the mite density increases for the larvae available, V. destructor can affect severely and rapidly a colony, and often eliminate it in the fall (Meikle et al., 2007). Varroa prefers young to mature bees, probably these last have a greater concentration of geraniol, that strongly repels the mite (Hoppe and Ritter, 1989), and commonly there are females of the parasite on foraging bees, that disperse them (phoresis), while serving as short term hosts (Meikle et al., 2007). The mite bores soft parts between the abdominal sclerites or behind the neck of the bee, and feeds its haemolymph. When arriving to an active colony, the parasite gets off and looks for rearing cells with 3rd stage larvae (Meikle et al., 2007), it enters them before they are sealed and hides in them (Donzé and Guerin, 1997), it sticks to the larval abdomen while this weaves its cocoon, and begin to feed from it when the prepupa develops (Meikle et al., 2007).
The traditional control of the mite with chemicals leaves residues that contaminate the honey and wax (Shaw et al., 2002; Kanga et al., 2003, Sima, 2013), it achieves insufficient results, and their repeated use has developed resistance (Gerson et al., 1991; Milani, 1995; Colin et al., 1997; Neira et al., 2003). Plant oils are also used (Sammataro et al., 1998; Menn, 1999; Chandler et al., 2001), with varied efficacy and a narrow range of selective doses (Kanga et al., 2003). They are also complex compounds that may cause undesired effects on honeybees and beekeepers (Schaller and Korting, 1995), and could also contaminate the hive products (Sammataro et al., 2000). Other control possibility is the use of pathogenic fungi that do not affect A. mellifera. Among them stand Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae (Metschnikoff) Sorokin, species that act on a wide range of arthropods (Poinar and Thomas, 1984; Khetan, 2001), including V. destructor (Kanga et al., 2002, 2003; Sterk et al., 2002; Meikle et al., 2007). Although the hymenopterans can be susceptible, the social species have behavior mechanisms that avoid infection or minimize their effects (Lord, 2000; Aumeier, 2001). In A. mellifera, the nursing bees remove dead and weak juveniles (Büchler et al., 1992 (1993); Boecking et al., 1993], a very variable hygienic conduct (Büchler, 1994) that reduces varroasis in untreated hives, that require then less chemical treatments (Spivak, 1996).
According to Hamiduzzaman et al. (2012), entomopathogenic fungi could reduce varroa mite damage to honey bee brood by both infecting the parasite and preventing varroa-associated suppression of honey bee immunity.
In laboratory bioassays by Ramírez and Gerding (no publication date), M. anisopliae strain Qu-M845 has obtained 98% control of Varroa, and 67% in field trials when applied in the fall on and between the combs. Sprays in the spring increased 50% the fall of the mite from the level in untreated hives. Further, two B. bassiana isolates caused varroa mortality (20 to 30%) after 6 days of having been treated (Rodríguez et al., 2009).
Meikle et al. (2007) found that B. bassiana had no effect on colony weight, adult bee weight or honey production.
With the hypothesis that the use of isolates of B. bassiana and M. anisopliae does not affect the normal conduct of nursing bees, a study was done to compare the conduct of healthy nursing bees with others exposed to isolates of both fungi, in order to evaluate their possible use for control of V. destructor.
MATERIALS AND METHODS
In this study, developed in the spring of 2006 in the Technological Centre of Biological Control of INIA in Quilamapu, Chillán, Chile, 1-day old healthy bees were used, selected at sunset from healthy hives in glass cages, and incubated a night at 35ºC to stimulate emergence from the cells. From these bees, ten nursing workers were selected at random, that were set in a glass cage with an 8 × 8 cm comb piece with larvae, cut with a 90% alcohol disinfected knife to avoid rotting dead larvae at the borders.
To obtain the inoculum, the fungi B. bassiana strain Qu-B303 and M. anisopliae strain Qu-M845, from the INIA >800 isolate collection at Quilamapu, Chillán, Chile (Rodríguez and Gerding, 2005), were inoculated onto Petri dishes with PDA and incubated at 25ºC until sporulation. The conidia were collected from the surface of these dishes, and suspensions were made on sterile distilled water with 1% Tween 80 moisturizer at 0.1%, and homogenized with a magnetic stirrer. The concentration of conidia in each solution, prepared in 10 mL solution of 30% honey in distilled water, was verified in 108 conidia mL-1, that according to Kanga et al. (2003) kill V. destructor, using a BOECO Neubauer counting chamber (Neubauer, Germany).
To evaluate the effect of the fungi on the nursing bees, 2 mL were poured of each spore solution on chicken water providers with small sponge pieces to allow drinking and avoid the bees from drowning. The bees were provided also with 3 cm diameter 1:1 pollen granules with honey to feed them. As control for comparisons bees unexposed to the fungi were used. Ten nursing bees were used for each treatment in 23.5 x 19.5 x 18.5 cm glass cages with red light and the corresponding drinking water provider, and were set in the dark at 30ºC, in a completely random experiment design, with 5 replicates. At 18 h from the beginning of the bioassay a bee was selected at random from each cage for observation under red light (not visible for the bees) during 3 min, during which conduct parameters were registered using the Observer® software, that allows to collect, analyze, and present observations measurements of behavior through activities, postures, gestures, expressions, movements, and interactions, both in humans and animals (Suazo et al., 2003). The conduct parameters evaluated were the time spent walking, antennal tapping, feeding, drinking, resting, communication (repeated movements of the antennae in front of another bee), and grooming. To facilitate observations, each activity was registered in a PC with the Observer program using separate keys. The records were made of 15 bees per treatment during 9 days, period which they behave as nursing bees (Lesser, 2004). The results obtained graphs for each activity during the bioassay at 18, 42, 66, 90, 114, 138, 162, 186, and 192 h exposure to the treatments were normalized and subjected to analysis of Variance and Dunnett tests.
RESULTS
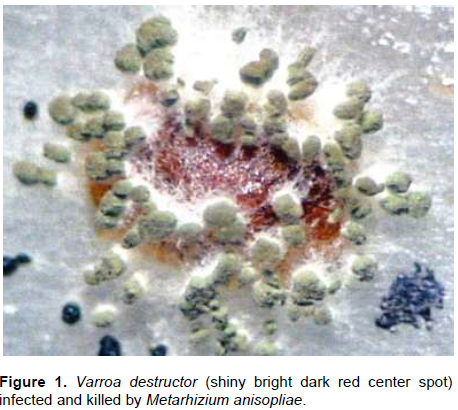
The results of exposing the nursing bees to M. anisopliae are presented in Figure 1, where the V. destructor mites killed by the fungus are easily identified.
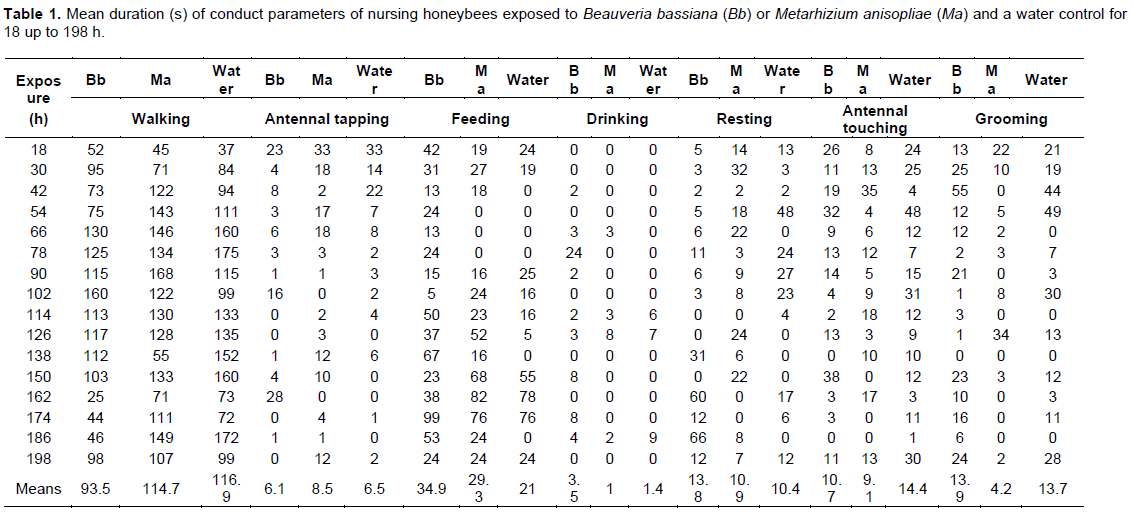
The results obtained when registering each conduct parameter are presented in Table 1, where no significant differences occurred between the treatments in all parameters, thus indicating just small variation throughout the exposure bioassay, but importantly, no effects on nursing honey bees.
Walking
The results presented in Table 1 varied numerically from 25 to 175 s.
Antennal tapping
The results in Table 1 neither presented significant differences between the control and the treatments with both fungi throughout the bioassay. Antennal tapping was very variable (0-33 s), but insufficient to cause statistical differences.
Duration of feeding
As in the previous parameters, the results of the feeding period in Table 1 were not significantly different between the control and the exposure treatments with both fungi, but presented wide numerical variations (0-99 sec).
Duration of drinking
Again, the results of the time spent drinking in Table 1 varied only numerically between the control and the exposure treatments with both fungi (0-24 s).
Duration of resting periods
The same as for the displacements and antennal tapping, the results of the resting time in Table 1 were not significantly different but presented wide numerical variation differences (0-66 s) between the control and the exposure treatments with both fungi.
Duration of antennal communication activities (touching)
The mean registered values in Table 1 were also not significantly different between the control and the exposure treatments with both fungi, although they varied numerically from 0 to 48 s.
Duration of grooming
These activities, estimated as very important for the sanitary condition of the hive (Büchler et al., 1992 (1993); Boecking et al., 1993), were again not significantly different between the control and the exposure treatments with both fungi, but varied greatly during our bioassay (0-55 s).
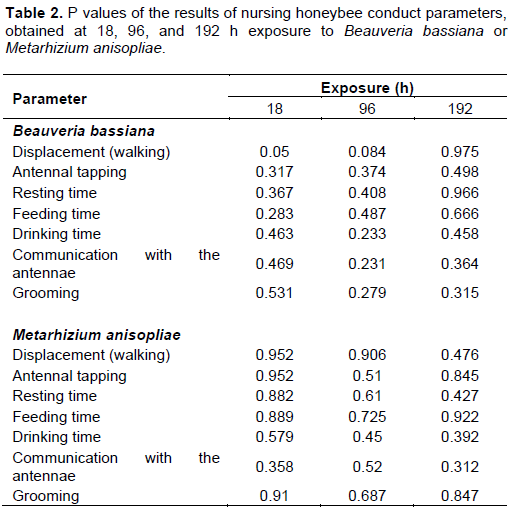
Despite the lack of statistical differences in all conduct parameter between the treatments, the P values of the results are presented at 18, 96 and 192 h exposure in Table 2.
DISCUSSION
The absence of significant differences in the conduct parameters of nursing bees exposed to both fungi indicate that it is possible to use B. bassiana or M. anisopliae in treatments against V. jacobsoni. The honeybee has a series of behavior characteristics that protect it of fungal infections, such as a hard and non-permeable cuticle, gut pH and peripheral membrane, that together with the tracheal system form an effective mechanical and physiological barrier (Gliñski and Buczek, 2003). The honeybee is characterized by the rapid detection and removal of sick and dead individuals, and the detailed cleansing of the hive by nursing workers (Flores et al., 1998). For example, this behavior is important in the resistance of the bees chalkbrood an aspergillosis, caused by the fungi Ascosphaera apis (Maasen ex Clausen) Olive & Spiltoir, and Aspergillus flavus Link, respectively; the workers retire with their mandibles the mummified larvae and carry them out of the hive (Gliñski and Buczek, 2003).
Our results agree with Lord (2000), who pointed that although many hymenopterans are susceptible to B. bassiana, their behavior including mechanisms of grooming, hygiene care of the nest, secretion of antimicrobial compounds and temperature regulation, avoid or minimize infection by pathogens, which is in line with the results of Flores et al. (1998), and Ibrahim and Spivak (2006) for bacteria, fungi, and arthropods (like V. destructor). This mite is highly susceptible to B. bassiana and M. anisopliae at a concentration of 108 conidia/mL and temperature similar to that reached in the hives of A. mellifera. The fungi during their reproductive phase do not infect V. destructor into the operculate cells of A. mellifera, but this occurs when the mites emerge to keep feeding from a newly emerged bee, which is not affected by the microorganisms (Kanga et al., 2002). The death of the mite caused by the fungi is due to the mechanical destruction of its tissues, the loss of water, and the effect of toxins (Chandler et al., 2001).
Kanga et al. (2002) indicated that the effects of B.
bassiana and M. anisopliae on honeybees and the environment of the hive require more investigation, because these fungi present a high potential as a biological control alternative against V. destructor, as they do not affect the normal behavior of the bees. In field studies with B. bassiana in southern France, Meikle et al. (2007) observed a significant increase in the percentage of infection on V. destructor that increased also in untreated hives suggesting certain conidial movement between colonies, probably derived from flying workers.
Sima (2013) determined in laboratory bioassays the pathogenicity of nine Canadian isolates of fungi of the genera Metarhizium, Beauveria, and Clonostachys on the parasitic mite V. destructor. All isolates were pathogenic to V. destructor with M. anisopliae UAMH9198, C. rosea UAMH 9161 and B. bassiana GHA being the most pathogenic within each species, with LC50 values of 1.6 ×105, 9.6 × 106, and 5.4 × 106 conidia/mL, respectively. M. anisopliae and B. bassiana affected brood and adult honey bee survivorship and their immune responses; however, they affected the bees, with LC50s of 3.70 × 106 and 2.62 × 105 conidia/mL, respectively. Combined treatments of fungi and thymol caused significantly greater mite mortality than single fungal treatments, which obtained ≤61% control levels.
CONCLUSION
Exposure to B. bassiana Qu-B303 or M. anisopliae Qu-M845 at 10 conidia/mL at 30ºC in darkness did not affect the normal behavior of nursing A. mellifera bees, thus the use of both fungi strains for control of V. jacobsoni should continue to be studied, including other best strains in further research.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Aumeier P (2001). Bioassay for grooming effectiveness towards Varroa destructor mites in Africanized and Carniolan honey bees. Apidologie 32:81-90. |
|
|
Boecking OW Rath W Drescher (1993).Grooming and removal behavior-strategies of Apis mellifera and Apis cerana bees against Varroa jacobsoni. American Bee Journal 133:117-119. |
|
|
Büchler R (1994). Varroa tolerance in honey bees occurrence, characters and breeding. Bee World 75:54-70. |
|
|
Büchler RW (1992) Drescher I, Tornier (199-93).Grooming behaviour of Apis cerana, A. mellifera and A. dorsata and its effect on the parasitic mites Varroa jacobsoni and Tropilaelaps clareae. Experimental and Applied Acarology 16(4):313-319. |
|
|
Chandler D, Sunderland KD, Ball BV, Davidson G (2001). Prospective biological control agents of Varroa destructor n.sp., an important pest of the European honey bee, Apis mellifera. Biocontrol Science and Technology 11(4):429-448. |
|
|
Colin ME, Vandame R, Jourdan P, Pasquale SDi (1997). Fluvalinate resistance of Varroa jacobsoni (Acari: Varroidae) in Mediterranean apiaries of France. Apidologie 28(6):375-384. |
|
|
De Jong D (1997). Mites: varroa and other parasites of brood. Chapter 2:13-31 In, Morse, RM and PK Flottum (eds.), Honey bee pests, predators, and diseases, 3rd ed., Root, Medina, Ohio. |
|
|
Donzé G, Guerin PM (1997). Time-activity budgets and space structuring by the different life stages of Varroa jacobsoni in capped brood of the honey bee, Apis mellifera. Journal of Insect Behavior 10(3):371-393. |
|
|
Flores JM, Ruiz JA, Ruz JM, Puerta F, Campano F, Padilla F, Bustos M (1998). El grooming en pis mellifera iberica frente a Varroa jacobsoni. Archivos de Zootecnia 47:213-218. |
|
|
Gerson U, Mozes-Koch R, Cohen E (1991). Enzyme levels used to monitor pesticide resistance in Varroa jacobsoni. Journal of Apicultural Research 30(1):17-20. |
|
|
Gliñski Z, Buczek K (2003). Response of the Apoidea to fungal infections. Apiacta 38:183-189. |
|
|
Hamiduzzaman MM, Sinia A, Guzmán-Novoa E, Goodwin PH (2012). Entomopathogenic fungi as potential biocontrol agents of the ecto-parasitic mite, Varroa destructor, and their effect on the immune response of honey bees (Apis mellifera L.). Journal of invertebrate pathology 111(3):237-243. |
|
|
Hoppe H, Ritter W (1989). The influence of the Nasonov pheromone on the recognition of house bees and foragers by Varroa jacobsoni. Apidologie 19(2):165-172. |
|
|
Ibrahim A, Spivak M (2006). The relationship between hygienic behavior and suppression of mite reproduction as honey bee (Apis mellifera) mechanisms of resistance to Varroa destructor. Apidologie 37(1):31-40. |
|
|
Kanga LHB, RR James, DG Boucias. (2002). Hirsutella thompsonii and Metarhizium anisopliae as potential microbial control agents of Varroa destructor, a honey bee parasite. Journal of Invertebrate Pathology 81(3):175-184. |
|
|
Kanga LHB, Jones WA, James RR (2003). Field trials using the fungal pathogen Metarhizium anisopliae (Deuteromycetes: Hyphomycetes) to control the ectoparasitic mite, Varroa destructor (Acari: Varroidae) in honey bee, Apis mellifera. (Hymenoptera: Apidae) colonies. Journal of Economic Entomology 96(4):1091-1099. |
|
|
Khetan S (200)1. Microbiological pest control. Chapter 9:211-221. Marcel Dekker, New York, USA. 300p. |
|
|
Lesser R (2004). Manual de apicultura moderna. Ed. Universitaria, Santiago, Chile. 213p. |
|
|
Lord J (2000). Response of the wasp Cephalonomia tarsalis (Hymenoptera: Bethylidae) to Beauveria bassiana (Hyphomycetes: Moniliales) as free conidia or infection in its host, the sawthooted grain beetle, Oryzaephilus surinamensis (Coleoptera: Silvanidae). Biological Control 21(3):300-304. |
|
|
Márquez ME, Fernández-Larrea O, Díaz D, Díaz A, Carrera B (2003). Evaluación de un producto de Bacillus thuringiensis para el control de la varroasis. Fitosanidad 7(1):3-8. |
|
|
Meikle WG, Mercadier G, Holst N, Nnansen C, Girod V (2007). Duration and spread of an entomopathogenic fungus, Beauveria bassiana (Deuteromycota: Hyphomycetes), used to treat varroa mites (Acari: Varroidae) in honey bee (Hymenoptera: Apidae) hives. Journal of Economic entomology 100(1):1-10. |
|
|
Menn J (1999). Biopesticides: present status and future prospects. Chapter 1: 1-10 in, Biopesticides, use and delivery. Humana Press, New Jersey, USA. 626p. |
|
|
Milani N (1995). The resistance of Varroa jacobsoni Oud. to pyrethroids: a laboratory assay. Apidologie 26(5):415-429. |
|
|
Neira M, Heinsohn P, Carrillo R, Báez A, Fuentealba J (2003). Efecto de aceites esenciales de lavanda y laurel sobre el ácaro Varroa destructor Anderson & Trueman (Acari: Varroidae). Agricultura Técnica 64(3):238-244. |
|
|
Poinar GO, Thomas G (1984). Laboratory guide to insect pathogens and parasites. Chapter 10: 105-110. Plenum Press, New York, USA. 392 p. |
|
|
Rodríguez M, Gerding M (2005). Control biológico de varroa. Tierra Adentro 65:20. |
|
|
Rodríguez M,Gerding M, France A (2009). Selection of entomopathogenic fungi to control Varroa destructor (Acari: Varroidae). Chilean Journal of Agricultural Research 69(4):534-540. |
|
|
Sammataro D, Degrandi-Hoffman G, Needham GR, Wardell G (1998). Some volatile plant oils as potential control agents for varroa mites (Acari: Varroidae) in honey bee colonies (Hymenoptera: Apidae). American Bee Journal 138:681-685. |
|
|
Sammataro D, Gerson U, Needham G (2000). Parasitic mites of honey bees: Life history, implications, and impact. Annual Review of Entomology 45(1):519-548. |
|
|
Schaller M, Korting HC (1995). Allergic airborne contact dermatitis from essential oils used in aromatherapy. Clinical and Experimental Dermatology 20(2): 143-145. |
|
|
Shaw K, Davidson G, Clark S, Ball B, Pell J, Chandler D, Sunderland K (2002). Laboratory bioassays to assess the pathogenicity of mitosporic fungi to Varroa destructor (Acari: Mesostigmata), an ectoparasitic mite of the honey bee, Apis mellifera. Biological Control 24(3):266-276. |
|
|
Sima A (2013). Evaluation of the fungi Beauveria bassiana, Metarhizium anisopliae, and Clonostachys rosea as bio-control agents against the honey bee parasitic mite, Varroa destructor. PhD diss. The University of Guelph, Ontario, Canada 181 p. |
|
|
Spivak M (1996). Honey bee hygienic behavior and defense against Varroa jacobsoni. Apidologie 27(4):245-260. |
|
|
Sterk G, Heutz F, Merck N, Bock J (2002). Sensitivity of non-target arthropods and beneficial fungal species to chemical and biological plant protection products: results of laboratory and field trials. 1st International Symp. International Symposium on Biological Control of Arthropods, Honolulu pp. 14-18. |
|
|
Suazo A, Baldwyn T, Teal PEA, Tumlinson JH (2003). Response of the small hive beetle (Aethina numida) to honey bee (Apis mellifera) and beehive-produced volatiles. Apidologie 34(6):525-533. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


