Full Length Research Paper

ABSTRACT
The effect of drought stress on Abscisic acid (ABA) on roots and root characteristics of three alfalfa varieties (Aohan, Zhongmu No.1 and Suntory) was studied. The study was conducted in greenhouse of the Chinese Academy of Agricultural Sciences (CAAS), Beijing, China from September 2014 to May 2015. Alfalfa varieties were exposed to four irrigation levels of water-holding capacity viz 100% (w1) (control); 85% (w2); 70% (w3) and 55% (w4), which were considered as deficit irrigation treatments. The results showed that ABA was significantly affected by deficit irrigation treatments during different plant growth stages. With the exception of 90 and 105 days after transplanting, ABA increased after transplanting up to harvest in all treatments. Increasing deficit irrigation showed significant increase in ABA content in roots of alfalfa. The minimum content of ABA was obtained from w1 (44 ng/g.FW) and then increased to 56.6, 64.6 and 94.4 ng/g.FW for w2, w3 and w4 respectively on 105 days after transplanting. Moreover, the results showed that ABA content was differently affected among different varieties of alfalfa under different water stress levels. The maximum ABA content was 83.2, 61.7 and 49.9 ng/g.FW obtained with Aohan, Suntory and Zhongmu No.1 varieties respectively, at 105 days after transplanting. Highest water stress w4 (55%) reduced root length by 20.92%, lateral roots by 20.71%, root fresh biomass by 43.79% and root dry biomass by 37.96%. The root to shoot ratio was 1.9 times higher in water stressed plants compared with the control, indicating that water stress in alfalfa is affected more shoot growth than root one.
Key words: alfalfa, deficit irrigation, Abscisic acid (ABA), root characteristics.
INTRODUCTION
In recent years, drought has becomes a leading threat to agriculture in the world (Loss and Siddique 1994; Grando and Ceccarelli 1995). It is one of the main environmental stresses that limits plant growth and production (Lawlor and Cornic, 2002; Martínez et al., 2003; Tadina et al., 2007; Ren et al., 2007; Wu et al., 2009). Drought affects many Medicago species (Chebouti et al., 2001; Bouizgaren et al., 2011). Plants can avoid drought by having a root system adapted to drought (Morgan et al., 1986; Liu et al., 2005).
Roots are the first part of a plant to sense water stress and its growth affects the answer to drought cues (Shimazaki et al., 2005). Root characteristics are defined as root length, root laterals number and root diameter are very important for good plant stand (Nanjo et al., 1999). Effect of water stress on root growth was studied in different crops such as wheat (Pritchard et al., 1991), maize (Wu and Cosgrove, 2000; Shimazaki et al., 2005) and barley (Sahnoune et al., 2004). One adaptation by plants to drought is the change in root to shoot ratio (Turner, 1997). Drought decreases growth of amaranth (Amaranthus spp.) in both root and shoot, but with less effect on root growth (Liu and Stützel, 2004). Lateral roots are one of the root system components which play an important role in water absorption. The development of lateral roots and growth are affected by complex interactions of different factors such as, environment, development and hormones (Casimiro et al., 2003; López-Bucio et al., 2003; Malamy, 2005). One important factor that affects the growth and development of roots to tolerate the environment such as water, drought, salt, cold, light, and temperature is ABA hormone (Signora et al., 2001; Sharp and LeNoble, 2002). ABA hormone is a key hormone that confers tolerance to environmental stresses. Plants increased ABA content when placed under water stress which effectively closes stomata to protect plants from adverse effects of drought (Schroeder et al., 2001; Rock et al., 2010; Mori and Murata., 2011) and to enhance drought resistance in crops (Schroeder et al., 2001; Wang et al., 2005; Shinozaki and Yamaguchi-Shinozaki, 2007). This study was conducted to evaluate the effects of deficit irrigation on abscisic acid content in roots and root characteristics of alfalfa.
MATERIALS AND METHODS
Plant material and destructive samples
The experiment was conducted in the green house of Animal Science Institute, Chinese Academy of Agricultural Sciences (CAAS), Beijing, China from September 2014 to May 2015.The treatments comprised of three varieties of alfalfa (Medicago sativa L.) viz Aohan, Zhongmu No.1 and Suntory. Seeds were germinated first on wet filter paper in petri-dishes. Four plants seeding were transplanted to each pots-culture (25 x 30 cm) with10 kg clay soil (pH 7.7%, organic matter 9.2 g kg-1, total P 0.7 g kg-1, total K 19 g kg-1, bulk density 1.3 g cm-3 and field capacity 30%). The varieties were evaluated under four irrigation levels of water holding capacity viz: w1 (100%), w2 (85%), w3 (70%) and w4 (55%). The full water holding capacity w1 was considered as normal irrigation, while the other treatments w2, w3 and w4 were considered as deficit irrigation. Irrigation water was application (manually) commenced from the first day of transplanting and continued for the whole growing period of alfalfa (195 days).
Pot moisture contents measured daily by using HH2 moisture meter version 4.0 (Delta- T Devices Ltd. UK) to maintained the percentage of water-holding capacity (100, 85, 70 and 55%). Samples of roots were collected every 15 days and immediately frozen in liquid nitrogen and then stored at -80 0C for further ABA extraction and analysis. Samples of roots were carefully removed from the pot and washed several times in water to obtain the complete roots. ABA was then extracted and purified according to Dobrev and Kaminek (2002) method and analyzed as described by Albacete et al. (2008). One homogenized gram of fresh root weight was placed in 5 ml of cold mixture of methanol, water and formic acid at pH 2.5 and was separated by centrifuging (20 000 g for 15 min after overnight extraction at -20°C. The root material was then subjected to generate more extract in 5 ml of the same solution for 30 min. Supernatants were filtered through Sep-Pak Plus†C18 (®Waters, Milford, MA, USA) to remove plant pigments and interfering lipids and evaporated to dryness. Residues were dissolved in 5 ml of methanol/water solution (1 to 4 ratio) using an ultrasonic bath and filtered by nylon membrane Millex filters (Ø 0.22 μm) (®Millipore, Bedford, MA, USA) then put in tubes adding extraction solution adjusting to 1.5 ml. Analyses were carried out on an HPLC/MS system consisting of an Agilent 1100 Series HPLC (Agilent Technologies, Santa Clara, CA, USA). Equipped with autosampler and connected to an Agilent Ion Trap XCT Plus mass spectrometer using ESI (an electrospray interface). It was filtered with Millex filters and injection, 100 μl of each fraction. For quantification of ABA calibration curves were constructed for component analyzed using internal standards: [2H6]cis,trans-abscisic acid (©Olchemin Ltd, Olomouc). Root length (cm) and lateral roots number/plant were measured by using Scan Maker i800 plus (MICROTEK, shanghai, China). Fresh root biomass (g) was weighted with digital balance directly after sample of root collection and then dried at oven 60°C until a constant weight for determining dry biomass of roots. Root to shoot ratio were calculated by dividing fresh root weight by the fresh shoot weight.
Experimental design and statistical analysis
The experiment was laid out in complete randomized design (CRD) with four replications. Treatments were three varieties of alfalfa and four irrigation level of water holding capacity. Data were calculated and arranged as means and analysis using one way analysis of variance (ANOVA) followed by Duncan’s multiple range test (P < 0.05).
RESULTS
Deficit irrigation and Abscisic acid (ABA) content
The effects of deficit irrigation treatments on the content of ABA in roots were shown in Figure 1. The results show that the content of ABA was affected by deficit irrigation at different plant growth stages. Increasing deficit irrigation showed a significant increase in ABA content. ABA content gradually increased with time under all deficit irrigation treatments until 75 days after transplanting. From 75 days after transplanting, ABA content hastily decreased under w1, w2 and w3 for 30 days until it reached the lowest ABA content at 105 days after transplanting. From 105 days after transplanting ABA content tended to increase until harvest. Moreover, under w4, ABA content increased from 30 to 75 days after transplanting, and then declined until 90 days and remained to 105 days after transplanting, after that tended to increase until harvest. The minimum ABA content was obtained under w1 (44 ng/g.FW) and then increased to 56.6, 64.6 and 94.4 ng/g FW for w2, w3 and w4 at 105 days after transplanting, respectively.
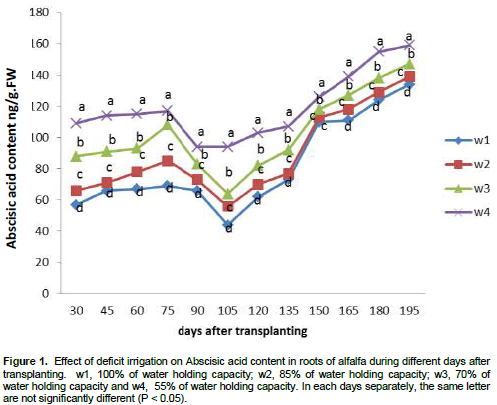
Abscisic acid (ABA) content and varietal response
The effect of alfalfa varieties on ABA content are presented in Figure 2. ABA content was affected by different varieties of alfalfa. ABA content progressively increased with time in all varieties up to 75 days after transplanting. ABA content under Zhongmu No.1 and Suntory decreased rapidly after 75 days after transplanting until reached its lowest activity level at 105 days after transplanting while Aohan decreased gradually. After that all the three varieties started to increase in ABA content and continued with that trend until harvest.
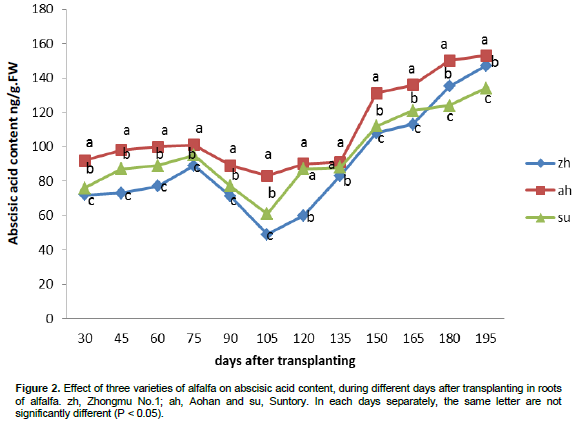
The minimum ABA content was 83.2, 61.7 and 49.9 ng/g .FW which was obtained by Aohan, Suntory and Zhongmu No.1 at 105 days after transplanting respectively. However, Zhongmu No.1 presented the lowest ABA content in all times, except for 180 and 195 days after transplanting in which was higher than Suntory while Aohan recorded the highest ABA content compared to the other two varieties.
Deficit irrigation and root characteristics
The effect of deficit irrigation treatments on root length, number of root laterals, fresh and dry weights of roots and root to shoots ratio are shown in Table 1. The results reveal that root length, number of root laterals, fresh and dry weights of roots were significantly reduced by increasing the deficit irrigation (P<0.05). The lowest reduction in root length was 9.3% followed by 12.7 and 20.9%. The least reduction in number of root laterals was 14.5% followed by 15.7 and 20.7%. Whereas the lowest reduction in root fresh weight was 9.9% followed by 11.1 and 43.8%. However, the least reduction in root dry weight was 7.3% followed by 21.7 and 38% was obtained by w2, w3 and w4 respectively, compared with control plants. Root to shoot ratio was significantly increased by increasing the deficit irrigation (P<0.05). The maximum increase in root to shoot ratio was 94.3%, followed by 78.6 and 30% in w4, w3 and w2 respectively compared to control treatment.
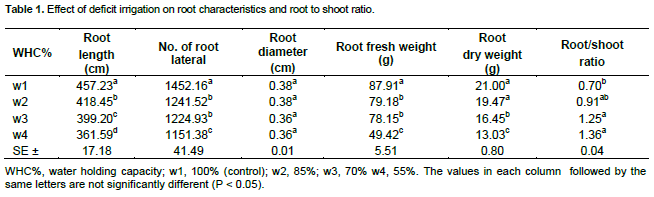
Root diameter was not significantly increased by increasing deficit irrigation. Irrigation of alfalfa plants by 55% of water holding capacity (W4) resulted in highest reduction in root diameter (5.3%) in comparison to the control plants (100% of water holding capacity).
Alfalfa varieties and root characteristics
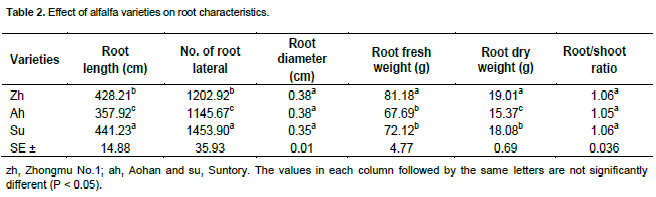
The effect of alfalfa varieties on root length, number of root laterals, root diameter, fresh and dry weights of roots are shown in Table 2. Aohan resulted in significantly different root length, number of root laterals and dry root biomass, while the minimum root diameter was observed in Suntory. Moreover, Zhongmu No.1 presented higher root fresh and dry weights compared with other two varieties. The highest root laterals number was observed in Suntory while no significant difference was observed in root to shoot ratio among all varieties.
DISCUSSION
Abscisic acid (ABA) is a hormone that indicates to endure environmental conditions such as drought and high salinity. Our study indicated that increasing deficit irrigation significantly increased ABA content. These results are in agreement with those reported by Brodribb and McAdam (2011) and McAdam and Brodribb (2012) on ferns (Pteridium esculentum and D. antarctica) and a lycophyte (Selaginella kraussiana). Outlaw (2003) reported that during drought stress ABA concentration was increased up to 30- fold.
Deficit irrigation reduced root length. This result is similar to that reported by Benlaribi et al. (1990); Ali Dib and Monneveux (1992) who attributed the reduction in root length and root fresh and dry weight to the reduction of turgor pressure in wheat. Similar results were observed in Albizzia seedlings (Nanjo et al., 1999), Erythrina seedlings (Nativ et al, 1999), Eucalyptus microtheca seedlings (Marron et al., 2002), and Populus species (Nautiyal et al., 2002). Moreover, Sacks et al. (1997) and Rao et al. (1993) found that under deficit irrigation, root growth of wheat and maize was not significantly reduced. The results showed that fresh and dry weights of roots were significantly reduced by increasing the deficit irrigation. Similar results were reported by Manivannan et al. (2007) and Lowlor and Cornic (2002) in pearl millet. Nicholas (1998) observed that in Avocado cultivars, water stress reduced the fibrous roots biomass. Under mild and severe water stress conditions, Populus species decreased root dry weight (Wullschleger et al., 2005). Similar results were observed in sugar beet (Pan et al., 2002). Furthermore, Amina et al. (2014) reported that drought reduced significantly the vegetative mass of the shoots and roots of wheat. Liu and Stützel (2004) observed that the effect of water stress was less in root growth than shoot one.
Our results showed that root to shoot ratio was significantly increased by increasing the deficit irrigation. Other studies demonstrated that under water-stress conditions, root-to-shoot ratio was increased to facilitate water absorption (Morgan 1984; Nicholas, 1998). Also Sharp and LeNoble (2002) and Manivannan et al. (2007) related this increase in water absorption to ABA content in roots and shoots.
CONCLUSION
Our results demonstrate that ABA was significantly affected by deficit irrigation treatments and Alfalfa varieties. Increasing deficit irrigation showed significant increase in roots ABA content. Moreover, root length, laterals number, fresh and dry weights of roots were significantly reduced by increasing the deficit irrigation. The results showed that the root to shoot ratio was significantly increased by increasing the deficit irrigation, indicating that water stress in alfalfa crop affected more shoot growth than roots. Aohan presented the lowest root length, number of root laterals, dry root biomass and less root to shoot ratio. In contrast this variety had the highest ABA content.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This work was supported by National Natural Science Foundation of China (31372370) and National Forages Industry and Technology System (CARS- 35-12).
REFERENCES
|
Albacete A, Ghanem M E, Martinez-Andujar C, Acosta M, Sanchez- Bravo J, Martinez V, Lutts S, Dodd IC, Perez-Alfocea F (2008). Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 59:4119-4131. |
|
|
Ali Dib T, Monneveux P (1992). Adaptation à la sécheresse et notion d'idéotype chez le blé dur. Caractères Morphologiques d'enracinement. Agronomie 12:371-379. |
|
|
Amina L, Ahmed A, Youcef H, Samira S (2014). Study of the Mechanisms involved In the tolerance of Durum Wheat (Triticum durum desf.) to Drought to Improve productivity. Turk. J. Agric. Nat. Sci. 1:682- 687. |
|
|
Benlaribi M, Monneveux P, Grignac P (1990). Etude des caractères d'enracinement et de leur rôle dans l'adaptation au déficit hydrique chez le blé dur (Triticum durum Desf.). Agronomie 10:305-322. |
|
|
Bouizgaren A, Farissi M, Khalida R, Ghoulam C, Barakate M, Al Feddy MN (2011). Assessment of summer drought tolerance variability in Mediterranean alfalfa (Medicago sativa L.) cultivars under Moroccan fields conditions. Arch. Agro. Soil Sci. 1-14. |
|
|
Brodribb TJ, McAdam SAM (2011). Passive origins of stomatal control in vascular plants. Science 331:582-585. |
|
|
Casimiro IT, Beeckman N, Graham R, Bhalerao H, Zhang P, Casero G, Sandberg M J, Bennett (2003). Dissecting Arabidopsis lateral root development. Trends Plant Sci. 8:165-17. |
|
|
Chebouti A, Mefti M, Abdelguerfi A (2001). Effet du stress hydrique sur le rendement en gousses et en grains chez trois espèces de luzernes annuelles: Medicago aculeteata, Medicago orbicularis Medicago truncatula. Cah. Opt. Médit. 45:163-166. |
|
|
Dobrev PI, Kaminek M (2002). Fast and efficient separation of cytokinins from auxin and abscisic acid and their purification using mixed-mode solid-phase extraction. J. Chromatogr. A. 950:21-29. |
|
|
Grando S, Ceccarelli S (1995). Seminal root morphology and coleoptile length in wild (Hordeum vulgare ssp. spontaneum) and cultivated (Hordeum vulgare ssp. vulgare) barley. Euphytica 86:73-80. |
|
|
Liu F, Stützel H (2004). Biomass partitioning, specific leaf area and water use efficiency of vegetable amaranth (Amaranthus spp.) in response to water stress. Sci. Hortic. 102:15–27. |
|
|
Liu F, Christian R, Shahanzari JA, Andersen MN, Jacobsen EE (2005). ABA regulated stomata control and photosynthetic water use efficiency of potato (Solanum tuberosum L.) during progressive soil drying. Plant Sci. 168:831-836. |
|
|
López-Bucio J, Cruz-Ramírez A, Herrera-Estrella L (2003). The role of nutrient availability in regulating root architecture. Curr. Opin. Plant Biol. 6:280-287. |
|
|
Loss SP, Siddique KHM (1994). Morphological and physiological traits associated with wheat yield increases in Mediterranean environments. Adv. Agron. 52:229-276. |
|
|
Lowlor DW, Cornic G (2002). Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant. Cell Environ. 25:275-294. |
|
|
Malamy JE (2005). Intrinsic and environmental response pathways that regulate root system architecture. Plant Cell Environ. 28:67-77. |
|
|
Manivannan P, Jaleel CA, Kishorekumar A, Sankar B, Somasundaram R, Sridharan R, Panneerselvam R (2007). Drought stress induced changes in the biochemical parameters and photosynthetic pigments of cotton (Gossypium hirsutum L.). Indian J. Appl. Pure Biol. 52:369-372. |
|
|
Marron N, Delay D, Petit JM, Dreyer E, Kahlem G, Delmotte FM, Brignolas F (2002). Physiological traits of two Populus × euramericana clones, Luisa avanzo and dorskamp, during water stress and re watering cycle. Tree Physiol. 22:849-858. |
|
|
Martínez JP, Ledent JF, Bajji M, Kinet JM, Lutts S (2003). Ef¬fect of water stress on growth, Na+ and K+ accumulation and water use efficiency in relation to osmotic adjustment in two populations of Atriplex halimus L. Plant Growth Regul. 41: 63-73. |
|
|
McAdam SAM, Brodribb TJ (2012). Fern and lycophyte guard cells do not respond to endogenous abscisic acid. Plant Cell 24:1510-1521. |
|
|
Morgan JM (1984). Osmoregulation and water stress in higher plants, Annu. Rev. Plant Physiol. 35:299-319. |
|
|
Morgan JM, Hare RA, Fletcher RJ (1986). Genetic variation in osmoregulation in bread and durum wheats and its relationship to grain yield in a range of field environments. Aust. J. Agric. Res. 37:449-457. |
|
|
Mori IC, Murata Y (2011). ABA signaling in stomatal guard cells: lessons from Commelina and Vicia. J. Plant Res. 124:477-487. |
|
|
Nanjo T, Kobayashi M, Yoshiba Y, Sanada Y, Wada K, Tsukaya H, Kakubari Y, Yamaguchi Shinozaki K, Shinozaki K (1999). Biological functions of proline in morphogenesis and osmotolerance revealed in antisense transgenic Arabidopsis thaliana. Plant J. 18:185-193. |
|
|
Nativ R, Ephrath JE, Berliner PR, Saranga Y (1999). Drought resistance and water use efficiency in Acacia saligna. Aust. J. Bot. 47:577–586. |
|
|
Nautiyal PC, Ravindra V, Joshi YC (2002). Dry matter partitioning and water use efficiency under water deficit during various growth stages in groundnut. Indian J. Plant Physiol. 7:135-139. |
|
|
Nicholas S (1998). Plant resistance to environmental stress. Curr. Opin. Biotechnol. 9:214-219. |
|
|
Outlaw WH (2003). Integration of cellular and physiological functions of guard cells. Crit. Rev. Plant Sci. 22:503-529. |
|
|
Pan XY, Wang YF, Wang GK, Cao RD, Wang J (2002). Relationship between growth redundancy and size inequality in spring wheat populations mulched with clear plastic film. Acta Phytoecol. Sin. 26:177-184. |
|
|
Pritchard J, Wyn Jones RG, Tomos AD (1991). Turgor, growth and rheological gradients of wheat roots following osmotic stress. J. Exp. Botany 42:1043-1049. |
|
|
Rao RCN, Williams JH, Wadia KDR, Hubikk KT, Fraquhar GD (1993). Crop growth, water use efficiency and carbon isotope discrimination in groundnut genotypes under end season drought conditions. Ann. Appl. Biol. 122:357-367. |
|
|
Ren J, Dai WR, Xuan ZY, Yao YA, Korpelainen H, Li CY (2007). The effect of drought and enhanced UV-B radiation on the growth and physiological traits of two contrasting poplar species. For. Ecol. Manage. 239:112-119. |
|
|
Rock CD, Sakata Y, Quatrano RS (2010). Stress signaling I: the role of abscisic acid (ABA). In: Pareek A, Sopory SA, Bohner HJ, Govindjee (eds) Abiotic stress adaptation in plants. Springer Dordrecht pp. 33-73. |
|
|
Sacks MM, Silk WK, Burman P (1997). Effect of water stress on cortical cell division rates within the apical meristem of primary roots of maize. Plant Physiol. 114:519-527. |
|
|
Sahnoune M, Adda A, Soualem S, Kaid-Harche M, Merah O (2004). Early water deficit effect on seminal root barley. C.R. Bilogies III. Agron. 327:389-398. |
|
|
Schroeder JI, Allen GJ, Hugouvieux V, Kwak JM, Waner D (2001). Guard cell signal transduction. Annu. Rev. Plant Physiol. Plant Mol. Biol. 52:627-658. |
|
|
Sharp RE, LeNoble ME (2002). ABA, ethylene and the control of shoot and root growth under water stress. J. Exp. Bot. 53: 33-37. |
|
|
Shimazaki Y, Ookawa T, Hirazawa T (2005). The root tip and accelerating region supress elongation of the decelerating region without any effects on cell turgor in primary roots of maize under water stress. Plant Physiol. 139:458-465. |
|
|
Shinozaki K, Yamaguchi-Shinozaki K (2007). Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 58:221-227. |
|
|
Signora L, De Smet I, Foyer CH, Zhang H (2001). ABA plays a central role in mediating the regulatory effects of nitrate on root branching in Arabidopsis. Plant J. 28:655-662. |
|
|
Tadina N, Germ M, Kreft I, Breznik B, GaberšÄik A (2007). Effects of water deficit and selenium on common buckwheat (Fagopyrum esculentum Moench.) plants. Photosynthetica 45:472-476. |
|
|
Turner NC (1997). Further progress in crop water relations. Adv. Agron. 58:293-338. |
|
|
Wang Y, Ying J, Kuzma M, Chalifoux M, Sample A, McArthur C, Uchacz T, Sarvas C, Wan J, Dennis DT, McCourt P, Huang Y (2005). Molecular tailoring of farnesylation for plant drought tolerance and yield protection. Plant J. 43:413-24. |
|
|
Wu TY, Feng DL, Bai ZY, Yang XJ, Liu XJ, Zheng YP (2009). Advances of research on drought-resistant mechanism of wheat. Agric. Res. Arid Areas 27:97-100. |
|
|
Wu Y, Cosgrove Dj (2000). Adaptation of roots to low water potentials by changes in cell wall extensibility and cell wall proteins. J. Exp. Bot. 51:1543-1553 |
|
|
Wullschleger SD, Yin TM, DiFazio SP, Tschaplinski TJ, Gunter LE, Davis MF, Tuskan GA (2005). Phenotypic variation in growth and biomass distribution for two advanced-generation pedigrees of hybrid poplar. Can. J. For. Res. 35:1779-1789. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


