Full Length Research Paper

ABSTRACT
Competition interferes with the growth and development of plants, whether of the same species or not. The aim of this work is to evaluate the intra-specific interference of slender amaranth plants (Amaranthus viridis) at different densities and distances, and in an inter-specific interaction with red pepper (Capsicum baccatum var. pendulum). The experiment was conducted in four replications and in a randomized complete block design using a factorial scheme of 2x4 + 2 controls, which represent two distances (5 and 10 cm) between the weeds and pepper (transplanted in the center of the box) and 4 densities of slender amaranth (3, 6, 9, and 12 plants m-2). Cement-asbestos boxes with a capacity of 90 L were filled with clayey soils (Red Dark Latosol). The pepper seedlings were transferred to the boxes after having three fully expanded leaves, whereas the slender amaranth seedlings were transplanted when they were 5 cm in height. Growth and yield characteristics of the crop and weeds were evaluated. As a result, it was observed that as the density increased, both species suffered more damage; the red pepper showed etiolation and reduced production, and the weeds showed a reduction in growth parameters. The distance between the plants did not interfere with their intra- and inter-specific coexistence.
Key words: Capsicum baccatum, Amaranthus viridis, density, competition, weed.
INTRODUCTION
Red pepper (Capsicum baccatum L.), belonging to the family Solanaceae, is a shrubby plant, and cultivated as an annual plant (Filgueira, 2008); it has indeterminate growth, continuous flowering and fruiting, and bears fruits at different stages of maturation (Pereira et al., 2014). It is produced mainly in family farms, where value can be added to the product, and in integration systems between the farmer and the industry (Rufino and Penteado, 2006).
The presence of weeds is one of the main factors that reduce the yield and quality of pepper fruits (Eure and Culpepperhtps, 2017). Among the plants found in the areas cultivated with vegetables, species of the genus Amaranthus, popularly known in Brazil as “caruru” (amaranth), stand out. Belonging to the family Amaranthaceae, this genus has about 60 plant species, some being cultivated and others growing as weeds: A. viridis (slender amaranth), A. spinosus (spiny amaranth), A. retroflexus (giant amaranth), and A. hybridus (purple amaranth) (Lorenzi, 2000). These plants can produce up to 235000 seeds (Lorenzi, 2000). This shows the importance of the management of these plants in agricultural areas to avoid competition with crops and to minimize the effects on the seed bank, especially considering the difficulty of their control (Robinson et al., 2008).
Amaranthus is one of the major genera of weeds infesting peppers in Mexico and Canada (Amador-Ramirez, 2002; Robinson et al., 2008). It is also one of the most important weeds for various bean cropping areas (Barroso et al., 2012) and vegetable cultivation sites (Marcolini et al., 2010). Barroso et al. (2012), in studies of inter-specific competition between beans and A. viridis, reported that the presence of weed decreased yield by 26%, negatively affecting the crop.
Knowledge of weed interference on crops is important in management decision making. The degree of weed interference on crops can be influenced by factors related to the weed community and the crop itself, such as the species present, the distribution, and the density (Barroso et al., 2012; Marcolini et al., 2010).
Competition is the main interference of one plant over another (Pitelli, 2014), and occurs when one or more of the essential resources for the growth and development of a plant becomes limited to meet the number of individuals present at the site (Rigoli et al., 2008). In this context, the density (number of individuals) and the spatial arrangement (plant spacing) of plants are factors that directly interfere with the relationship between plants (Bezerra et al., 2014). According to Carvalho and Christoffoleti (2008), high density is a more important factor for competition than the intrinsic competitive ability of the species. Thus, it is important that plants can remain under adequate conditions and in environments with available resources, so that competition is not established.
Most studies on the competition between plants are focused on comparisons between weeds and cultivated plants (inter-specific competition), seeking to quantify the effect on yield (Christoffoleti and Victoria Filho, 1996). Notwithstanding, intra-specific competition, that is, between plants of the same species, is also of great importance, since the plants have the same vegetative habit and the same needs of resources. Crops such as sunflower (Bezerra et al., 2014) and beans (Carvalho and Christoffoleti, 2008; Barroso et al., 2012) showed a decrease in yield when under intra-specific competition. However, it should be borne in mind that species agronomically considered as weeds may also suffer from intra-specific competition under conditions where resources are limited.
The hypothesis of this work is that amaranth can interfere with pepper and this interference is dependent on the density and distance of the weed crop, which may also undergo density-dependent intra-specific interference. Thus, the aim of this study is to evaluate the intra-specific interference of slender amaranth (Amaranthus viridis) plants at different densities and distances, and in an inter-specific interaction with red pepper (C. baccatum).
MATERIALS AND METHODS
Two experiments were carried out. The first consisted of the coexistence of A. viridis (slender amaranth) and C. baccatum; in the second, A. viridis plants remained in intra-specific competition.
The seedlings of A. viridis and C. baccatum were produced in 128-cell trays filled with commercial substrate. The plants for both experiments were transplanted to the experimental unit on the same day, on August 3, 2015, according to each treatment. The pepper showed three fully expanded leaves, and the weeds were approximately 5 cm high. The transplantation was done in cement boxes with a capacity of 90.0 L, and dimensions of 60x60x25 cm, filled with clayey soil (Dark Red Latosol). Fertilizations were performed according to the results of the soil analysis (Table 1), based on Bulletin 100 (Van Raij et al., 1997);h the first fertilization was done 10 days before the transplant of the plants, and the cover fertilization 30 days after transplantation (DAT). Irrigation was done daily, and the insecticide Deltamethrin (concentration of 25 g L-1), with commercial product Decis (BAYER) was applied as recommended by the manufacturer and when necessary for crop protection.

The inter-specific experiment was performed in a randomized complete block design, with a factorial scheme of 2x4 + 2 controls, representing the two distances (5 and 10 cm) between the weeds and the pepper (transplanted in the center of the box), and the four densities of slender amaranth (1, 2, 3, and 4 plants box-1, representing 3, 6, 9, and 12 plants m-2, respectively). A pepper control was maintained without the presence of weeds, and a slender amaranth control without any crop.
For the intra-specific experiment, only the factorial scheme was modified to 2x3 + control, representing the two distances from the center of the box (5 and 10 cm) and the three densities (2, 3, and 4 plants box-1, corresponding to 6, 9 and 12 plants m-2, respectively). For the control treatment, there was only one plant per box (3 plants m-2).
The measurements of height (from the base to the beginning of the branch) and diameter at the stem base of the plants were made at 15, 30, 45, 60, and 75 DAT for A. viridis plants (leaves and inflorescence at the beginning of senescence), and at 15, 30, 45, 60, 75, 90, and 105 DAT for pepper (final ripening period). At the end of each experimental period, the following were evaluated: leaf area (LICOR LI3000), dry mass of leaves, and dry mass of stems (after drying in a greenhouse with forced air circulation at 65°C for three days). For the crop, the number of fruits was also counted, and, after harvest, the fresh and dry mass of fruits and the dry mass of seeds were evaluated.
The data were submitted to analysis of variance (ANOVA) and the means were analyzed by the Tukey test (p> 0.05).
RESULTS AND DISCUSSION
Inter-specific interference between pepper and A. viridis
Regarding the coexistence of pepper with A. viridis, a significant difference was observed in plant height from 60 days after transplantation (DAT) in relation to weed distance (Table 2). This effect was more accentuated by the distance the plants occupy, as it can be observed that the pepper showed a higher height at 5 cm from the weeds, mainly at the highest density, of 12 plants m-2. Notwithstanding, this result does not imply that the plant had an effective growth; the crop may have undergone light competition with weeds, which has caused etiolation.

Etiolation is a response of plants to low ambient light (Franco and Dillenburgl, 2007), in which the plant starts to invest more energy to increase stem growth (increase in length) in search of light (Dousseaun et al., 2007). However, this growth does not provide an increase in the dry mass of the plant, that is, this energy expenditure is not being well used and this may cause changes in plant metabolism (Constantin et al., 2008; Pitelli, 2014) and losses in yield. Moreover, the plant is more fragile and susceptible to breakage and to the attack of pests and diseases, which further hinders its development and cultivation (Silva et al., 2016).
This non-effective growth of the pepper plants is also observed in the other characteristics evaluated. There is a significant relation between the factors and the control for the stem diameter of pepper, and the presence of A. viridis negatively interfered with this characteristic from 45 DAT, decreasing it by 40% at 105 DAT with the density of 12 plants m-2 (Table 3). According to Lima et al. (2008), etiolated plants do not accumulate mass and, therefore, their stem is much thinner when compared to a healthy plant without interference.

For the dry mass of stems and leaves and leaf area (Table 4), a significant difference was observed for the control in relation to the factors, and the presence of weeds, regardless of density and distance, caused a reduction in these characteristics. The reductions reached 74, 71, and 25% for the dry mass of stems and leaves and leaf area at the density of 12 plants m-2, respectively.
The plants with higher height did not present higher values ​​of diameter and dry mass of stems and leaves, increasing the chances that the growth occurred as a function of the shading caused by weeds, that is, the pepper plants had etiolation. However, the leaf area was higher with 12 m-2 plants than in the other densities. Under low luminosity, plants tend to expand leaf size to compensate for or better use low light (Lima et al., 2008). However, this change in morphology implies other changes in the leaf, such as a decrease in leaf thickness (Benincasa, 2003), which explains the reduction of dry mass of leaves in the pepper plants, a phenomenon that Lima et al. (2008) found in Caesalpinia ferrea seedlings.

The interception of solar radiation by weeds, preventing the passage of light into the crop, depends on the composition, density, and distribution of these plants (Pitelli, 2014). Marcolini et al. (2010) verified that the beet crop was very sensitive to the interference imposed by A. viridis plants, showing a significant reduction in leaf area, number of leaves, dry mass of leaves, mean root diameter, and fresh mass of roots even at low densities of weed populations.
Based on the analysis of dry mass of leaves (Table 5), there was interaction between the factors distance and density of plants m-2. Comparing the densities of slender amaranth plants for each distance, it was observed that at the distance of 5 cm, there was no significant difference between the treatments. For the distance of 10 cm, it was observed a difference between the treatments with 3 and 12 plants m-2, where the first treatment resulted in a higher dry matter production, probably due to the competition for resources being lower.

For the number of fruits (Table 6), there was a significant difference for the control in relation to the factors. The presence of A. viridis caused a mean reduction of 68% at the densities of 3 and 6 plants m-2 (Table 6), and 81.5% at the other densities. It was observed that the distance of 5 cm provided higher losses for the number of fruits (Table 7) as density increased, reaching a reduction of 56%.

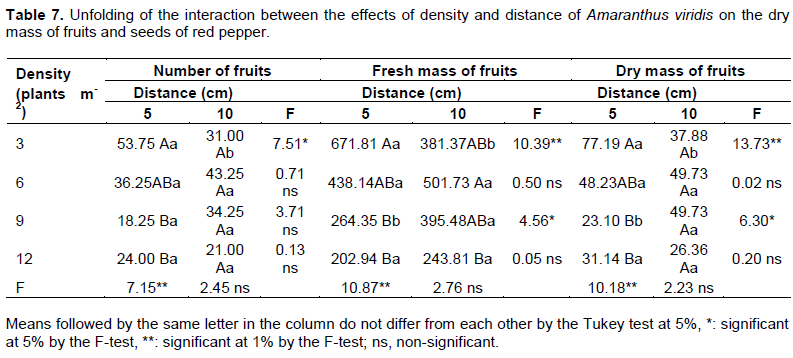
Regarding the fresh mass of fruits (Table 7), the control significantly differed from the factors; at the distance of 5 cm, the density interfered with fruit production. The treatment with 3 plants m-2 was similar only to that with 6 plants m-2.
Regarding the dry mass of fruits, the factors also differed from the control (Table 6). It is observed that the density had greater influence, since the greater the number of A. viridis plants, the smaller the dry mass of fruits. This effect is clearly observed in the unfolding of the treatments, where the reductions reach 60% at the distance of 5 cm in the second crop (Table 7).
For the dry mass of seeds (Table 6), the treatments differed from the control, and it was verified that the density had a greater influence than the distance.
When plants are competing both for space and for resources in the environment, those with faster growth and greater uptake of solar radiation occupy the space faster, hindering the growth and development of the others (Pitelli, 2014). In agricultural areas, weeds are considered more competitive, since they occur at a higher density, distribution, and with greater adaptive and reproductive capacity, causing greater damage to the cultivated plant (Pitelli, 2014; Bianchi et al., 2006).
Studying the periods of interference of weeds in pepper (C. annuum, cv. 'Mirasol') during three years in Mexico, with A. palmeri as one of the main weeds, Amador-Ramirez (2002) obtained that the period previous to interference (PPI) was 2.1 weeks after transplantation, tolerating a 5% loss in total production, or 0.9 weeks considering marketable production, with total period of weed interference (TPWI) of 12.2 and 12.3 weeks, respectively. Consequently, critical periods of weed interference (CPWI) were 2.1 to 12.2 and 0.9 to 12.3 weeks, that is these would be the periods in which the crop would have to be kept free of the presence of weeds to obtain at least 95% of the estimated total and marketable fruit production, respectively. In the present study, red pepper coexisted with slender amaranth plants for 105 days after transplantation, that is, 15 weeks, and this period exceeds the critical period obtained by Amador-Ramirez (2002), thus confirming the sensitivity of the crop to weed interference. In the case of this work, it was observed that red pepper suffered with the presence of A. viridis, since fruit production was impaired. The higher energy expenditure that pepper plants had to correct cell growth processes and recover the loss of resources (Paulus et al., 2015), especially light, may have led to a decrease in the photosynthetic rate and a lower translocation of energy and sugars for fruit formation.
Another important factor that may have caused the low fruit production is the fact that A. viridis has a C4 carbon fixation pathway (Carvalho and Christofoleti, 2008), and this photosynthetic mechanism confers several advantageous characteristics in relation to C3 plants, mainly in hot and humid environments (Paul and Elmore, 1984), which contributes to the presence of Amaranthus species in areas of production of oil plants such as pepper.
Omazine and Silva (2016) verified that with increasing density of A. lividus plants, there was a significant reduction in the cumulative dry mass of C. annuum. There was a reduction of 80.8% in the dry mass of pepper when cultivated with only one A. lividus plant, and a reduction of 97.7% when the treatment consisted of four A. lividus plants, showing that the crop growth was hampered due to the conviviality among the species, even at a low density. In addition, the authors point out that although the greatest damage occurs at high weed densities, the species A. lividus is more aggressive on pepper when it is in a smaller number of individuals than at a high population rate, since the competition among plants harms their own development.
Intra- and inter-specific interference of A. viridis
When analyzing the height of A viridis plants coexisting with pepper (Table 8), it is observed that the control (a single slender amaranth plant in the experimental unit, without pepper) differed from the other treatments from 45 DAT. It was also verified that from 60 DAT, the density caused a decrease in weed height, which reached up to 28% at 12 plants m-2.

The same behavior was observed in relation to the stem diameter (Table 9), in which significant differences were observed from 45 DAT between the treatments and the control. It can be verified that with the increase of the density of plants in coexistence with the pepper crop, there was a decrease in the diameter, but the distance between the plants did not show differences.

Regarding the dry mass of stems and leaves (Table 10), it was observed that the distance did not cause interference in plant growth, while the density gradually reduced the dry matter production. With 12 plants m-2, the reduction in dry mass was 70 and 75% for stem and leaves, respectively, for the distances of 5 and 10 cm. The same behavior was obtained for the leaf area of ​​the plants, with a 35% decrease in the treatment and highest density in relation to the control.

When analyzing the interaction between C. annuum and A. lividus plants, Omezine and Silva (2016) verified that the dry matter accumulation of A. lividus was modified according to the density and the coexisting species. The authors observed that the highest dry matter accumulation of A. lividus occurred in the treatment with four A. lividus plants associated with a single C. annuum plant (52.33 g/pot), while the lowest accumulation occurred in the treatment with one A. lividus plant growing in the presence of one C. annuum plant (35.77 g/pot).
When A. viridis was cultivated alone, that is, only under intra-specific interference, the plants showed a significant difference in plant height as a function of distance only at 30 DAT (Table 11), but density was the factor that mostly interfered with this parameter of plant growth. Notwithstanding, at 75 DAT, only the treatment with the highest number of plants differed from the control, with a 21% reduction in height. In the unfolding at 30 DAT (Table 12), it is observed that when the density is 12 plants m-2, the distance does not interfere with the plant response; instead, the number of competing individuals plays a key role. An important factor in the competition between plants is the availability of resources in the environment, in which the absence of a resource can cause reduction in biomass accumulation and yield (Pitelli, 2014).


As for the diameter under these conditions (Table 13), plant density interferes with plants at 45 DAT; at 75 DAT, the reductions reached up to 27.5% in the treatments with 6 and 9 plants m-2, and 45% in the treatment with 12 m-2 plants.

Regarding the dry mass of stems and leaves and leaf area (Table 14), it was verified that the factors differed significantly from the control, and that the greatest influence in the coexistence of plants is due to density: the higher the plant density, the smaller the results obtained for the three parameters. When analyzing the lowest and the highest density (6 and 12 plants m-2, respectively), the reductions were 53 and 78% for dry mass of stems; 47 and 71% for dry mass of leaves; and 21 and 45% for leaf area, respectively.

According to Omezine and Silva (2016), for the species A. lividus, intra-specific competition may be more important than inter-specific competition, since they verified that the accumulated dry matter of this species decreased when there was an increase in its density, in the same way as the total plot dry matter (C. annuum + A. lividus) decreased with increasing density of A. lividus. Magro et al. (2011), in turn, found that Cyperus difformis biotypes are more sensitive to coexistence with rice plants (Oryza sativa) than with plants of the same species, that is, inter-specific competition is more important. In contrast, for the rice crop, intra-specific competition was more important.
The results indicate that the factor density caused greater damage to the growth and development of both pepper and A. viridis plants, due to morphological changes in the plants of both species and a decrease in the production of pepper. The distance between plants did not prove to be a significant factor in the coexistence of plants. The increase in the density of A. viridis, mainly with 12 plants m-2, caused a reduction in stem diameter, leaf area, and in the number and mass of pepper fruits, besides causing plant etiolation; it also led to a reduction in height, stem diameter, leaf area, and dry mass of leaves of A. viridis.

CONFLICT OF INTERESTS
The authors declare that they have no conflict of interest.
REFERENCES
|
Amador-Ramirez MD (2002). Critical period of weed control in transplanted chilli pepper. Weed Research 42(3):20-209. |
|
|
Barroso AAM, Yamauti MS, Nepomuceno MP, Alves PLCA (2012). Efeito da densidade e da distância de caruru-de-mancha e amendoim-bravo na cultura do feijoeiro. Planta Daninha 30:47-53. |
|
|
Bezerra FT, Dutra AS, Bezerra MAF, Oliveira Filho AF, Barros GL (2014). Comportamento vegetativo e produtividade de girassol em função do arranjo espacial das plantas. Revista Ciência Agronômica 45:335-343. |
|
|
Bianchi MA, Fleck NG, Lamego FP (2006). Proporção entre plantas de soja e plantas competidoras e as relações de interferência mútua. Ciência Rural 36(5):1380-1387. |
|
|
Carvalho SJP, Christoffoleti PJ (2008). Competition of Amaranthus species with dry bean plants. Scientia Agricola 65(3):239-245. |
|
|
Christoffoleti PJ, Victoria Filho R (1996). Efeitos da densidade e proporção de plantas de milho (Zea mays L.) e caruru (Amaranthus retroflexus L.) em competição. Planta Daninha 14:42-47. |
|
|
Constantin J, Machado MH, Cavalieri SD, Oliveira Jr RS, Rios FA, Roso AC (2008). Influência do glyphosate na dessecação de capim-braquiária e sobre o desenvolvimento inicial da cultura do milho. Planta Daninha 26(3):627-636. |
|
|
Dousseaun S, Alvarenga AA, Santos MO, Arantes LO (2007). Influência de diferentes condições de sombreamento sobre o crescimento de Tapirira guianensis Alb. Revista Brasileira de Biociências 5(2):477-479. |
|
|
Eure PM, Culpepperhttps AS (2017). Bell pepper and weed response to dimethyl disulfide plus chloropicrin and herbicide systems. Weed Technology 31(5):694-700. |
|
|
Franco MAS, Dillenburgl LR (2007). Ajustes morfológicos e fisiológicos em plantas jovens de Araucaria angustifolia (Bertol.) em resposta ao sombreamento. Hoehnea 34:135-144. |
|
|
Filgueira FAR (2008). Novo manual de olericultura: agrotecnologia moderna na produção e comercialização de hortaliças. Viçosa: Editora da UFV. 421 p. |
|
|
Lima JD, Silva BMS, Moraes WS, Dantas VAV, Almeida CC (2008). Efeitos da luminosidade no crescimento de mudas de Caesalpinia ferrea Mart. ex Tul. (Leguminosae, Caesalpinoideae). Acta Amazônica 38(1):5-10. |
|
|
Lorenzi H (2000). Plantas Daninhas do Brasil: Terrestres, aquáticas, parasitas e tóxicas. Nova Odessa: Instituto Plantarum 608p. |
|
|
Magro TD, Shaedler CE, Fontana LC, Agostinetto D, Vargas L (2011). Habilidade competitiva entre biótipos de Cyperus difformis L. resistente ou suscetível a herbicidas inibidores de ALS e destes com arroz irrigado. Bragantia 70(2):294-301. |
|
|
Marcolini LW, Carvalho LB, Cruz MB, Alves PLCA, Cecílio Filho AB (2010). Interferência de caruru-de-mancha sobre características de crescimento e produção da beterraba. Planta Daninha 28(1):41-46. |
|
|
Omezine A, Silva JAT (2016). Competitive ability of Capsicum annuum L. relative to the weed Amaranthus lividus L. Journal of Horticultural Research 24(1):79-91. |
|
|
Paul R, Elmore CD (1984). Weeds and the C4 syndrome. Weeds Today 15:3-4. |
|
|
Paulus D, Valmorbida R, Santin A, Toffoli E, Paulus E (2015). Crescimento, produção e qualidade de frutos de pimenta (Capsicum annuum) em diferentes espaçamentos. Horticultura Brasileira 33(1):91-100. |
|
|
Pereira FECB, Torres SB, Silva ML, Grangeiros LC, Benedito CP (2014). Qualidade fisiológica de sementes de pimenta em função da idade e do tempo de repouso pós-colheita dos frutos. Revista Ciência Agronômica 45:737-744. |
|
|
Pitelli RA (2014). Competição entre Plantas Daninhas e Plantas Cultivadas. In: Aspectos da Biologia e Manejo de Plantas Daninhas (ed. by Monquero PA). Rima, São Carlos pp. 61-82. |
|
|
Rigoli RP, Agostinetto D, Schaedler CE, Dal Magro T, Tironi S (2008). Habilidade competitiva relativa do trigo (Triticum aestivum) em convivência com azevém (Lolium multiflorum) ou nabo (Raphanus raphanistrum). Planta Daninha 26(1):93-100. |
|
|
Robinson DE, McNaughton K, Soltani N (2008). Weed management in transplanted bell pepper (Capsicum annuum) with pre-transplant tank mixes of sulfentrazone, s-metolachlor, and dimethenamid-p. Hortscience 43(5):1492-1494. |
|
|
Rufino JLS, Penteado DCS (2006). Importância econômica, perspectiva e potencialidades no mercado para pimenta. Informe Agropecuário 27:7-15. |
|
|
Silva FJ, Hisatugo EY, Souza JP (2016). Efeito da luz na germinação e desenvolvimento de plântulas de pinhão-manso (Jatropha curcas L.) de distintas procedências. Hoehnea 43(2):195-202. |
|
|
Van Raij B, Cantarella H, Quaggio JA, Furlani AMC (1997). Boletim Técnico 100 – Recomendações de adubação e calagem para o Estado de São Paulo. Campinas: Instituto Agronômico/Fundação IAC. 258 p. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


