Full Length Research Paper

ABSTRACT
Development of diseases in cultivated crops depends on the complex inter-relationship between host, pathogen and prevailing environmental conditions. The significant role of nematodes in the development of nematode–fungus interaction is demonstrated in many crops throughout the world. However, there is scanty research information in Ethiopia. Therefore, the main objectives of this study were to: (i) investigate the effect of Meloidogyne incognita (MI)-Fusarium oxysporum f.sp. lycopersici (FOL) interaction on selected tomato genotypes based on their order of inoculation and (ii) evaluate the reaction of selected tomato genotypes against the MI-FOL interaction. The greenhouse experiment was laid out in a completely randomized design (CRD) factorial with four replications. Three-week-old tomato seedlings were inoculated with MI suspension at a rate of 3000 second-stage juveniles (J2) and 10 ml FOL suspension (3x106 conidia/ml/pot) around the root rhizosphere. Tomato growth, biomass and pathogen related data were recorded starting first week after inoculation to eight weeks of post inoculation. The result revealed that simultaneous inoculation of MI and FOL (NF) and FOL 10 days after MI (N1F2) was found significantly (p ≤ 0.05) reducing tomato growth, biomass and pathogen related parameters compared to single pathogen or un-inoculated control. Among the three tomato genotypes tested, Assila was moderately resistant as measured by the lower number of root gall and egg mass per plant, that it could be of a good choice to manage this disease complex or interaction. Performance evaluation study at MI-FOL hot spot farmers’ field should be investigated in the near future.
Key words: Fusarium oxysporum f.sp. lycopersici, Meloidogyne incognita, resistance, management, synergistic effect and tomato varieties
INTRODUCTION
Tomato (Solanum lycopersicum L.) is one of the most important vegetable crops across the world next to potato (McGovern, 2015). The fruits of tomato are popular throughout the world, are used in all kinds of vegetables as raw salad, and as processed products such as paste and juice. Ripe tomato fruit has high nutritive value and a good source of vitamin A, B, C and minerals (MoARD, 2009). Recently, it started gaining more medicinal value because of its high content of antioxidant including carotenoids, ascorbic acid, phenolics and lycopene (Oduor, 2016). It also serves for export, making a significant contribution to the national economy of Ethiopia with most of the exports to Djibouti, Somalia, South Sudan, Middle East and European markets (Tabor and Yesuf, 2012).
Tomato production reached to more than 163.4 million tons cultivated on more than 4.6 million hectares of land worldwide (FAOSTAT, 2016). In Ethiopia, it is being grown in about 6,298.63 hectares with an annual production of 28,364.83 tones (CSA, 2017). Large-scale production of tomato is being carried out in the central rift valley (CRV) under irrigated and rain-fed conditions having unlimited potentials for expansion if certain production constraints are avoided (Wondimeneh et al., 2013). Despite its rapid spread across the different localities and agro-ecologies, the production and productivity remains low. This is attributed to several biotic and abiotic factors. Among the biotic factors, plant diseases caused by plant-parasitic nematodes (PPN) are a costly burden. There are over 4100 species of PPN currently identified collectively causing an estimated loss of $80–$118 billion per year in damage to crops. Of these species, 15% of them are the most economical species directly targeting plant roots of major agricultural crops and prevent water and nutrient uptake resulting in reduced agronomic performance, overall quality and yields (Bernard et al., 2017). In nature plants are rarely, if ever, subject to the influence of only one potential pathogen and this is especially true of soil-borne pathogens like fusarium wilt (Fusarium oxysporum) whereby further opportunities exist for interactions with other microorganisms occupying the same ecological niche (Back et al., 2002).
The combined effect of wilt causing fungi and PPN causes serious damage to different economically important crops worldwide (Mai and Abawi, 1987; Chen et al., 2004). Based on its worldwide distribution, extensive host range and involvement with fungi, bacteria and viruses in disease complex, root knot nematodes (RKN) rank first among the top 10 damaging genera of PPN affecting the world’s food supply (Jones et al., 2013). Obtaining optimum crop quality and economic production of tomato depends on development and exploitation of an eco-friendly, sustainable, economical and alternative methods of nematode-wilt disease complex management. Being aware of the array of organisms influencing the crop and the nature of various organisms’ interactions is therefore essential (Webster, 1985). This indicates that development of simple technology for efficient and reliable integrated management of soil-born plant pathogens is highly dependent on knowledge of this interaction effect. Hence, there is an urgent need to generate basic information regarding disease complex involving RKN and soil-born fungi. The main objectives of this research work were to: (i) investigate the effect of the interaction between Meloidogyne incognita (MI) and Fusarium oxysporum f.sp. lycopersici (FOL) on selected tomato genotypes based on their order of inoculation and (ii) evaluate the reaction of selected tomato cultivars with different degree of resistance to M. incognita against M. incognita-F. oxysporum f.sp. lycopersici (MI-FOL) co-infestation under greenhouse conditions.
MATERIALS AND METHODS
Fungal isolates
Monoconidial isolates of F. oxysporum f.sp. lycopersici (FOL-W, FOL-P and FOL-V) were used in this study. These isolates were collected from infected tomato plant from the Central Rift Valley (CRV), Ethiopia and were characterized according to Leslie and Summerell (2006). Diseased plant specimens (stem bases and roots) were subjected to running tap water and then rinsed with distilled water. Diseased specimens were cut into pieces (2 cm) and surface sterilized with 2% NaOCl for 2 min followed by three changes of sterile distilled water and dried in between two sterilized blotting paper. Sterilized and dried specimens were plated out on potato dextrose agar (PDA) media in sterile Petri-dishes and incubated at 25 ± 2°C for 7-9 days.
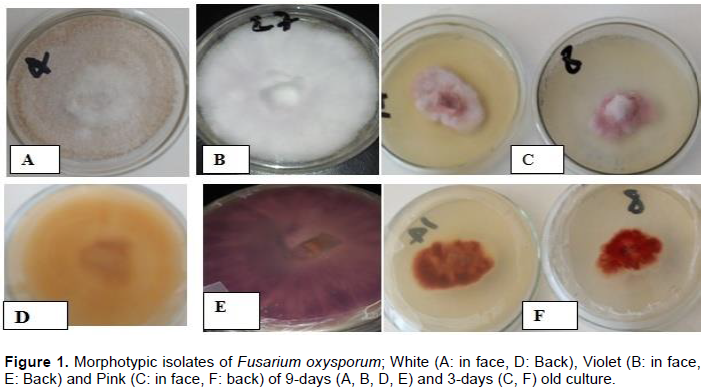
Pathogenicity test was used to confirm the identified F. oxysporum formae specials. Three-week-old susceptible (Marmande) tomato genotype seedlings were inoculated by standard root dip method (Srivastava et al., 2009). Four treatments: (i) Marmande + White FOL isolate, (ii) Marmande + Pink FOL isolate, (iii) Marmande + Violate FOL isolate and (iv) un-inoculated check with five replications were set under controlled greenhouse conditions. Pure culture of the aggressive fungal isolate (FOL-W) was used as a starting culture for the disease complex or interaction study (Figure 1). The test fungus was multiplied with PDA medium on 9 cm diameter Petri-dishes to get enough inoculum to help initiate the actual experiment. The three tomato genotypes, resistant (Assila), moderately resistant (Cochoro) and susceptible (Moneymaker) were inoculated with FOL conidia suspension based on the treatment requirement. Inoculum density (conidia concentration) of the pathogen was adjusted to 3x106 conidia/mL/plant (Lobna et al., 2016) using a Haemocytometer and 10 mL of this solution was delivered into holes in the soil surface of pots.
Meliodogyne incognita population
Molecularly (DNA-based and Isozyme techniques) identified M. incognita population by Seid et al. (2017) was used as a starting pure culture. This species originated from the central rift valley area from the 2016 tomato-growing season. The root galls containing the egg masses were used to inoculate three-week-old susceptible (Moneymaker) tomato seedlings grown in sterilized soil for mass multiplication of nematode under aseptic conditions. The culture was multiplied on several pots using the same genotype to get enough inoculums. After giving sufficient time of 70-80 days to complete 2-3 generations, the plants were de-potted carefully and root system was washed free of soil. Then roots free of attached soil particles were submerged on Phloxine B (0.15 g/L) for 15 min (Holbrook et al., 1983) to clearly observe the egg-masses and facilitate counting.
Second stage juveniles (J2) were obtained from egg masses by incubating large number of egg masses at room temperature in water. After 10 days of incubation, every two days the J2 in water were collected and kept in a refrigerator (8°C) in a 100 mL beaker and volume of water was made up to 50 mL. The nematode suspension (water containing J2) was fetched with the help of 10 mL pipette and an aliquot of one mL was transferred to counting dish for counting the juveniles under dissecting-binocular microscope. For inoculation, juvenile levels were adjusted with water so as to add equal volume of nematode suspension (3000 J2) in each treatment and were added to pot seedlings to the 2 cm deep holes around the roots of the tomato seedlings or rhizosphere soil.
Treatments and experimental design
The experiment was conducted in greenhouse with 18 treatment combinations using a completely randomized design (CRD) factorial with four replications (Table 1). Plants were maintained in a greenhouse at the temperature of 26 ± 2°C. The experiment lasted a total of two months and at the end, each treatment was hand harvested. The following data on M. incognita and F. oxysporum f.sp. lycopersici and plant related parameters were collected starting from 7 days after inoculation (DAI).
Pathogen related parameters
After cutting the top parts of the plants, all the pots were turned upside down with care, to discharge the soil and the roots were made free of soil. Finally, the plant roots were gently washed with tap water to remove adhering soil particles. Then the number of root-galls per root system was counted manually aided with hand lens. Roots containing egg-masses were soaked in Phloxine B (0.15 mg/L tap water) solution for 15-20 min and then the roots were rinsed in tap water to remove residual stain. The egg-masses were stained pink to red and observed and counted (Coyne, 2007) to determine number of egg-masses per plant (EMPP). Root gall index (RGI) and egg-mass index (EMI) per plant were determined from each pot and based on 0 to 5 rating scale (Taylor and Sasser, 1978); where, 0 = no galls or egg masses; 1: 1-2 galls or egg-masses; 2: 3-10 galls or egg-masses; 3: 11-30 galls or egg-masses; 4: 31-100 galls or egg-masses and 5: over 100 galls or egg-masses.
The final nematode population density (Pf) was estimated from organic (root) and mineral (soil) fraction per pot. The mean number of J2 in the roots was estimated from the whole root system after extracting nematodes from a sub-sample of 5 g roots per plant based on (Hussey, 1973). Nematodes from soil samples were extracted using a modified Baermann funnel technique. It was expressed as J2 per 100 gram of soil. The Pf of M. incognita was counted by transferring the suspension to nematode counting dish under a stereo microscope.
Disease severity was assessed weekly (visual observation) starting one week after inoculation up to eight weeks of post inoculation, where the final estimation was recorded and rated on a scale of 0-4 (Song et al., 2004); where, 0: no infection; 1: slight infection which is about 25% of full scale (one or two leaves become yellow); 2: moderate infection (two or three leaves became yellow, 50% of leaves become yellow); 3: extensive infection (all plant leaves became yellow, 75% of leaves become wilting) and 4: complete infection (the whole plant leaves became wilting, and growth was inhibited). Area under the disease progress curve (AUDPC) was calculated according to the method of Shaner and Finney (1977) using the formula:
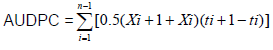
Plant related parameters
Plant height (PH) was measured from the soil level to the main apex of the plant eight weeks after transplanting and mean values were calculated per treatment and expressed in cm. Root length (RL) was taken after the adhering soil was gently washed away from the roots using tap water and excess water was removed after blotting with tissue paper. The root length per plant was measured from the soil level to the tip of 75% of roots end and expressed in centimeter.
The tomato plant was cut at the crown level in each pot and the fresh shoot weight (FSW) was measured (in gram) using electronic balance soon after cutting. After cutting the top parts of the tomato plants, all the pots were turned upside down with care, to discharge or dislodge the soil and the roots were made free of adhering soil. Finally, the plant roots were gently washed with tap water to remove adhering soil particles. Then, the fresh root weight (FRW) was measured (in gram) using electronic balance. The shoots were put in paper bag and brought to laboratory just after taking the fresh weight and kept in an oven at 105°C for 24 h, and allowed to come to room temperature and the dry shoot weight was measured (in gram) using electronic balance to determine its dry shoot weight (DSW).
Data analysis
Data obtained from the greenhouse experiment were subjected to analysis of variance (ANOVA) and means were separated using LSD at P=0.05 by GenStat software (16th edition).
RESULTS
Identification of Fusarium oxysporum
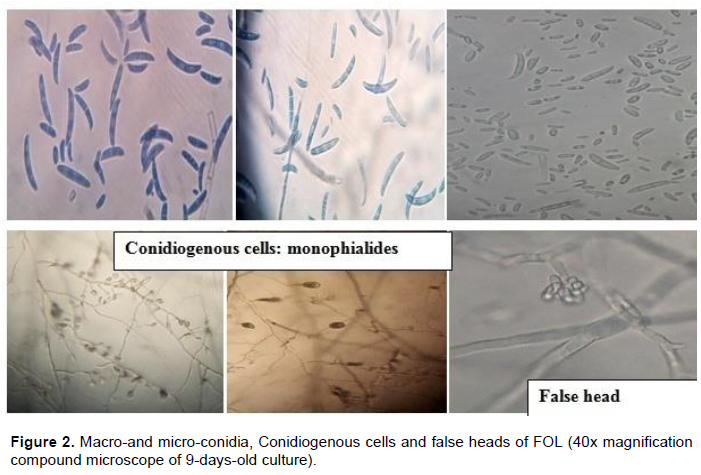
The length and breadth of macroconidia usually varied between (15.9-46.98 x 1.83-4.88 µm) and that of microconidia was 6.75-13.56 x 1.93-3.4 µm under 40 x compound microscope magnification. The number of septation of macroconidia ranges from 1.5-4.3. The shape of macroconidia varied from straight, slightly curved to sickle shaped. The shape of microconidia was oval, elliptical or kidney and usually 0-septated with infrequent occurrence of a single septation (Figure 1). False head structures with short monophialides were there in the aerial mycelium while it was observed under compound microscope without disturbance of the existing mycelium (Figure 2).
Pathogenicity of Fusarium oxysporum
Various symptoms on aerial parts and within the stem tissues of tomato plants infected with F. oxysporum were noted starting at 33 DAI. Yellowing of the lower leaves at early stage of the plant and leaf necrosis and later, dropping due to the infection were the most prominent symptoms. However, there was no statistically significant difference in virulence among the isolates in mean disease severity and area under disease progress (Table 2). As expected, however, there was significant difference (p ≤ 0.05) between the inoculated treatments and the un-inoculated check regardless of the isolates (Table 2).
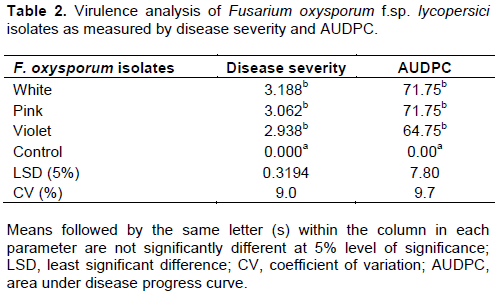
Number of root gall and root gall index
Data presented (Table 3) revealed that mean number of root gall per plant (RGPP) has significantly (p ≤ 0.01) increased in the treatment where the nematode was inoculated simultaneously with the fungus (NF) as compared to the treatment which received nematode 10 days later to fungus inoculation (F1N2) and the control (C) on Moneymaker genotype. The maximum mean number of RGPP (496.8) was recorded from treatments that received both pathogens simultaneously followed by FOL inoculated 10 days after M. incognita, N1F2 (453.8) and M. incognita alone, N (360.2) on susceptible genotype Moneymaker, although no significant variation among one another. However, the minimum number of root galls was observed on roots of Assila and Moneymaker genotypes when fungus infection preceded nematode in 10 days (F1N2).
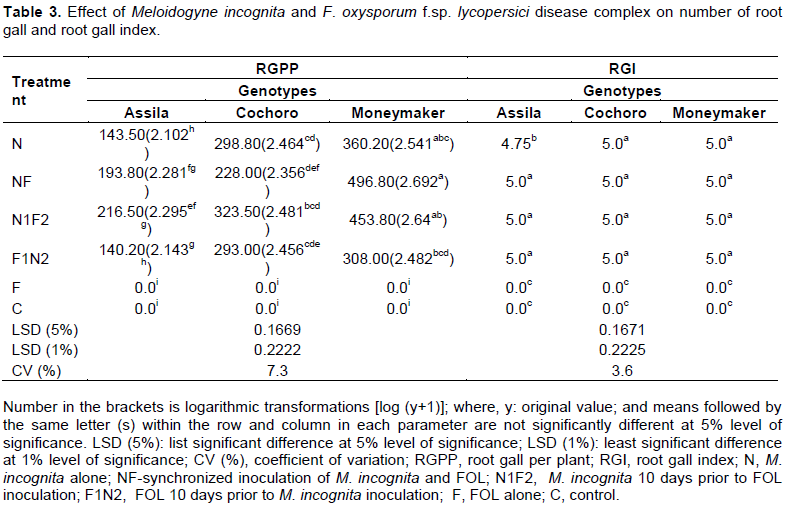
There was a significant difference in number of RGPP among the selected tomato genotypes when M. incognita inoculated with F1N2. A statistically highly significant (p ≤ 0.01) difference in mean number of RGPP was also found between Assila and Moneymaker tomato genotypes in all the treatments containing M. incognita. However, there was no statistically significant difference between Assila and Cochoro genotypes while they were inoculated with nematode and fungus concomitantly (NF). The minimum number of RGPP (140.2) was recorded on Assila genotype when inoculated with F1N2 followed by N (143.5). It is also possible to clearly notice from Table 3 that RGPP varied according to the susceptibility gradient of the cultivars used.
The interaction effect of the three selected tomato genotypes and disease complex involving M. incognita and FOL on the root gall index (RGI) was insignificant (p ≤ 0.05) except when inoculated with M. incognita alone (N). The effect of the treatments was also insignificant in root gall index except on Assila genotype where it was infested with the nematode, M. incognita alone (N) as compared to other treatments, including the un-inoculated check.
Number of egg-mass and egg-mass index
The highest number of egg-mass per plant, EMPP (347.2), was recorded from the treatment inoculated with N1F2 followed by inoculation of N (297.8) on the susceptible, Moneymaker tomato genotype (Table 4). A statistically highly significant difference (p ≤ 0.01) in the mean number of EMPP was observed on Moneymaker genotype when it was inoculated with N1F2 as compared to other treatments and the control except the treatment that received N. A statistically highly significant difference (p ≤ 0.01) in mean number of egg mass was also noted among the selected tomato genotypes but of between Cochoro and Moneymaker, while they were inoculated with NF (Table 4). Less mean number of EMPP (42.0) was recorded when inoculated with F1N2, followed by inoculation of N (89.2) on the resistant genotype (Assila).
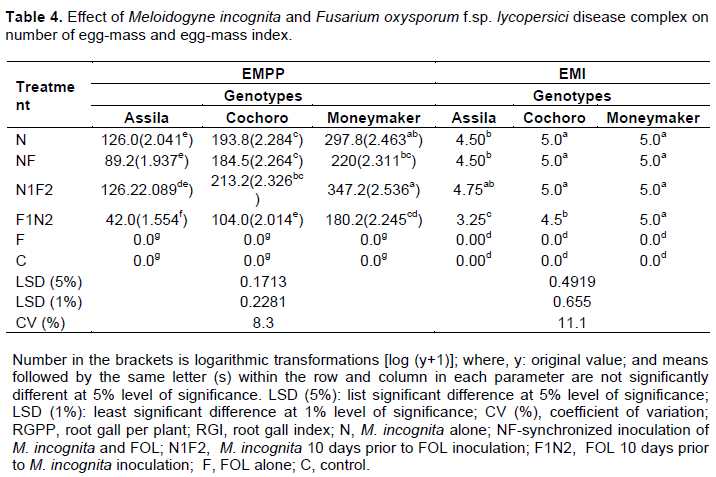
Generally, treatments receiving N1F2 and N resulted in increased and comparable number of egg mass across all the genotypes tested. There was invariably reduced number of EMPP in all the genotypes that received F1N2 (Table 4). The lowest egg mass index, EMI (3.25) was observed from the treatment inoculated with F1N2 on the resistant tomato genotype. Highly significant (p ≤ 0.01) interaction effect of the disease complex and tomato genotypes was also noted when fungus inoculation precedes nematode by 10 days in this study (Table 4).
Final population density (Pf) and reproduction factor (Pf/Pi)
The maximum mean final nematode population density [72002 (J2+eggs)] and reproduction factor (24) from 100cc soil and the entire root system was observed on Moneymaker genotype when it was inoculated with N1F2 (Table 5). In contrast, the lowest mean M. incognita count [18552 (J2 + eggs)] and reproduction factor (6.18) was on the resistant genotype, Assila where the reciprocal (F1N2) treatment was applied. There was highly significant variation (p ≤ 0.01) in nematode population density among treatments on all the genotype tested. However, this variation was not significant between treatments, which received NF and N on Assila genotype. The same is true between the treatments on genotype Moneymaker when N1F2 and N. As indicated in Table 5, the final population density of M. incognita in the treatment that received both pathogens concomitantly (NF) was numerically lower compared to nematode alone (N) inoculated treatment though not statistically significant.
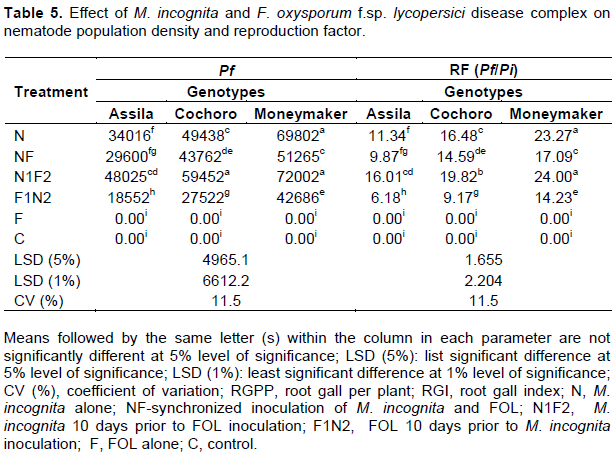
The interaction effect of disease complex and selected tomato genotypes was noted significant except between Cochoro and Moneymaker where the genotypes were inoculated with N1F2 (Table 5). However, the highest reproduction was on susceptible cultivar when nematode preceded fungus by 10 days. The reproduction of M. incognita invariably increased on all genotypes used when inoculated with both pathogens in the sequence of N1F2 or N.
Plant height (PH) and root length (RL)
The result indicated that when M. incognita was inoculated simultaneously with the FOL (68 cm) and FOL 10 days after M. Incognita (71.75 cm) reduced PH to a significant level (p ≤ 0.05) as compared to the control (106.5 cm) and the rest of the treatments except the treatment, which received M. incognita alone on Cochoro genotype (Table 6). The same is true in case of Moneymaker genotype where by the lowest mean PH (78.25 cm) was recorded from NF inoculated treatment. In contrast, the highest PH (113.0 cm) and (106.5 cm) were recorded from the control treatment on Moneymaker and Cochoro genotypes, respectively.
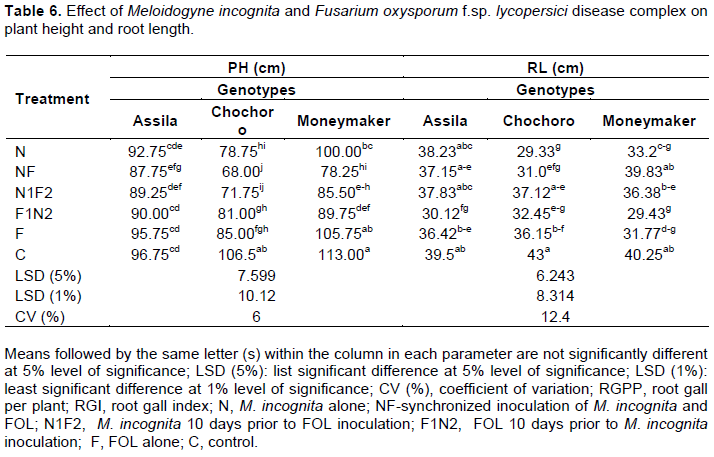
The result of the current study also showed statistically insignificant variation (p ≤ 0.05) between treatments receiving the two pathogens simultaneously and fungus after 10 days to nematode with respect to PH on Cochoro and Moneymaker genotypes. On the other hand, PH was highly significantly (p ≤ 0.01) affected by synchronized inoculation of the two pathogens regardless of the genotypes used, including the resistant one, Assila (Table 6). The interaction effect of M. incognita and FOL disease complex and the genotypes were statistically insignificant between Assila and Moneymaker except when inoculated with NF. However, this is not true in case of Assila and Cochoro genotypes.
The minimum (29.33 cm) mean root length was recorded from M. incognita alone inoculated treatment on Cochoro genotype, followed by nematode inoculated 10 days after FOL (29.43 cm) on the genotype Moneymaker. The maximum root length (43 cm) was from un-inoculated check on the same genotype. There was no statistically significant difference in mean root length among the three selected tomato genotypes, except in between Assila and Cochoro when inoculated with nematode alone.
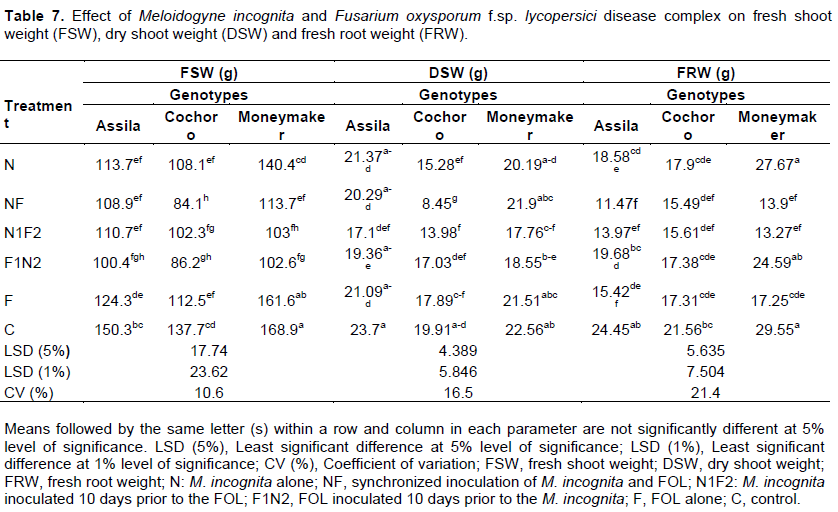
Fresh shoot weight (FSW)
The minimum (84.1 g) mean FSW was counted from pots inoculated with M. incognita and FOL simultaneously (NF) on Cochoro genotype. This was followed by the treatment, where the M. incognita was inoculated 10 days after FOL (F1N2); even though, there is no significant difference between each other. The maximum (168.9 g) fresh shoot weight was observed from un-inoculated treatment on the same genotype. Significant (p ≤ 0.05) interaction effect between treatments and genotypes were also observed where the genotypes were inoculated with NF, N and F. All the treatments inoculated with M. incognita and FOL significantly (p ≤ 0.05) reduced fresh shoot weight (FSW) than treatments, which received either of the pathogens alone, and the control on the susceptible genotype (Table 7).
Dry shoot weight (DSW)
The minimum (8.45 g) DSW, from M. incognita and FOL simultaneously inoculated pot and maximum (23.75 g) DSW, from un-inoculated pots were recorded on Cochoro and Assila genotypes, respectively. A significant (p ≤ 0.05) synergistic interaction effect of treatments and genotypes in terms of DSW were noted if the genotypes are infected with NF and N (Table 7).
Fresh root weight (FRW)
The lowest (11.47 g) mean FRW was recorded on simultaneously inoculated treatment with M. incognita and FOL (FN) on Assila genotype followed by treatment received FOL after 10 days to M. incognita, N1F2 (13.27 g) on Moneymaker genotype. There was significant (p ≤ 0.05) variation among the genotypes selected when exposed to M. incognita with absence of FOL (Table 7). Fresh root weight in M. incognita alone inoculated treatment was comparable to the un-inoculated treatment on Cochoro and Moneymaker genotype. Significant (p ≤ 0.01) reduction in FRW invariably across the genotypes was noted when it was inoculated two pathogens simultaneously and N1F2 and there is insignificant variation among the genotypes in this regard. The interaction effect of treatments and the genotypes was observed significant (p ≤ 0.05) if the genotypes are inoculated with M. incognita alone (Table 7).
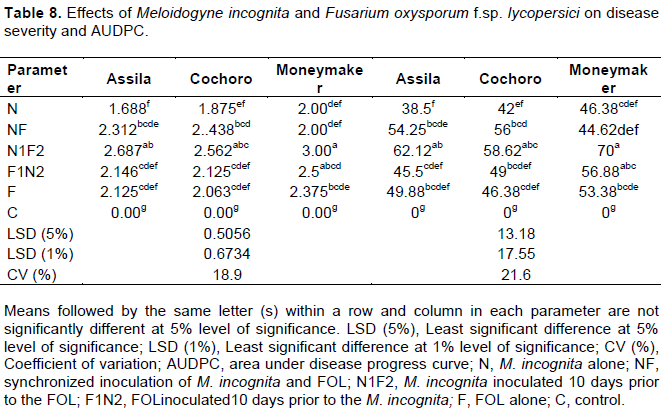
Disease severity and area under disease progress curve
The interaction effect of disease complex and genotype on mean disease severity score was insignificant (p ≤ 0.05). The main effect of the genotypes was also found to be insignificant and here under the main effect of the treatments, only is presented. This was invariably true in case of the resistant genotype, Assila and moderately resistant genotype, Cochoro on which highest mean disease score, 2.687 and 2.562 and area under disease progress curve, 62.12 and 58.62 respective order, was recorded from treatments infected with FOL 10 days later to M. incognita (Table 8).
DISCUSSION
This research result showed that the number of root gall and galling index caused by M. incognita has increased in the presence of fusarium wilt (FOL) either concomitantly (NF) or nematode 10 days prior to the fungus (N1F2). This might be attributed to increased penetration rate of M. incognita juveniles (J2) into the roots due to co-infection of both pathogens. Similar result was obtained by Al-Hazmi and Al-Nadary (2015); whereby synchronized inoculation of M. incognita race 2 and Rhizoctonia solani (N + F) increased the index of rhizoctonia root rot and the number of root galls on green beans (Phaseolus vulgaris L.). Minimum number of root gall in F1N2 treatment might indicate the unsuitability of root for J2 penetration and the fungus damaged lack of support for the nematodes to establish within the root system as it. F. oxysporum f.sp. lycopersici infection and establishment in plant roots previously infected by nematodes (N1F2) was enhanced as the developing root galls associated with nematode feeding may act as a nutrient sink. Elevated major organic constituents of root exudates mainly, carbohydrates and nitrogenous compounds during the first fourteen days after nematode infection is well established fact (Van Gundy et al., 1977; Mai and Abawi, 1987). These organic constituents are considered to be major nutrient consumptions for different fusarium wilt inciting fungal species like F. oxysporum that co-infect the same host plant. This result also supports /depicts these established research reports as determined by synergistic interaction effect of the two pathogens on the number of root galls. This is generally in line with nematode induced physiological modification/change theory of nematode fungus interaction in the host plant tomato. Maximum number of root gall on treatments when nematode was inoculated 14 days prior to the fungus and on synchronized inoculation of both pathogens with no statistically significant difference between each other on the same experimental host plant and pathogens was also reported (Khpalwak, 2012). Statistically insignificant variation between the resistant and moderately resistant tomato genotypes in number of root gall during co-infection of the two pathogens (NF) probably indicates loss of potential genetic resistance in resistant genotype due to co-infection of these pathogens and signifies the negative impact of the disease complex on the resistant potential of tomato plant as it was measured by the number of root gall. However, increased number of RGPP along with the susceptibility gradient of tomato genotypes implied that M. incognita resistant genotypes would also be promising for the management of the disease complex.
Similar trends of increase in number of egg-mass and egg-mass index in N1F2 and NF treatments across all the genotypes tested were observed. The influence of FOL and time of its application on M. incognita egg-mass development on tomato was most pronounced with N1F2. This clearly depicts the negative impact of FOL and its time of inoculation on the development of nematode egg. Similar previous findings of Yang et al. (1976), Al-Hazmi and Al-Nadary (2015) and Kumar et al. (2017) are in line with this result. This might be also attributed to reduced food sources for nematode as the root system is affected with the fungus in F1N2 inoculated treatment. Timing of application of nematode and fungi seems to matter the relationship between the invasions of tomato by M. incognita as it was also supported by Back et al. (2006) who prove the relationship between the invasion of potato roots by potato cyst nematodes and the percentage of stolon affected by Rhizoctonia solani was strongest 6 and/or 8 weeks after planting. Lowest EMI in F1N2 treatment however is against the previous result of Pauline (2016), who reported lower EMI in combined inoculation of F. oxysporum and Meloidogyne species as compared to inoculation of Meloidogyne species alone and highest EMI viz. single inoculation of Meloidogyne species on the same experimental host plant. This might be attributed to the genetic nature of tomato genotype used in the experiment as it was depicted with significant interaction effect of Assila and the treatments.
Measurement of nematode reproduction (host efficiency) and yield or growth is vital to quantify the reaction of plants to RKN infection (Mai and Abawi, 1987). High nematode population density and reproduction factor from N1F2 treatment, which was low with the reciprocal treatment (F1N2), may be explained by the nutrient competition effect of the two pathogens co-habiting the common host plant. Sugars in root exudates from M. incognita infected tomato increased from to 836% over un-inoculated check (Wang and Bergeson, 1974).
This indicates huge nutritional advantage of the nematode (M. incognita) obtained from the host plant response during M. incognita infection and the entire disease cycle. However, this nutritional advantage of the nematode could be suppressed if other pathogen of the same nutritional requirement existed on the same ecological niche that is soil rhizosphere and plant rhizoplane. Similar previous report in which infection of the fungus Verticillium albo-atrum resulted in no enhancement effect on reproduction of neither stubby root nematode (Trichodorus christie) nor M. incognita on or in the root of the host plant (tomato) was also reported (Conroy and Green, 1974). Invariably increased nematode population on all genotypes used when inoculated with both pathogens in the sequence of N1F2 indicates the unfavorable impact of M. incognita-FOL interactions on nematode reproduction probably due to the colonization of giant cells with fungi and thereby disturbing their function. The probable inhibitory effects of fungi metabolites on hatching of nematode eggs are also reported by Zahid et al. (2002). The decline in nematode populations involved in a disease complex with fungi may also be explained by competition for nutrients and root space between the two organisms (Back et al., 2002).
Invariably significant synergistic effect of the two pathogens on tomato plant growth and biomass was explained by reduction of plant height, root length, fresh shoot weight, and dry shoot weight with concomitant infection (NF). Similar previous research finding (Goswami and Agrawal, 1987; Johnson and Littrell, 1970; Kumar, 2008; Kumar et al., 2017; Lobna et al., 2016; Negron and Acosta, 1989) also indicated similar research finding. However, the interaction effect of the two pathogens on plant height has been also affected by the existing varietal variation. Assila is a genotype with determinate growth habit (bushy type that grows 60-90 cm tall) whereas Moneymaker is genotype with indeterminate growth habit, vining type tomato that grows 1.5-3 m tall. Root length was found unaffected when inoculated simultaneously with both pathogens (NF) which is similar with previous research report of Kumar et al. (2017) and against the research results of Goswami and Agrawal (1978).
The increase of fusarium wilt severity in the presence of M. incognita may be due to the fact that infection by this endo-parasitic nematode (M. incognita), whether prior to the fungal infection (N1F2) or simultaneously (NF), causes physiological and anatomical changes in the root tissues predisposing the plants to increased fungal infection (Al-Hazmi and AL-Nadary, 2015). This result is also in line with the finding of Katsantonis et al. (2003) on which invasion of the roots of cotton by M. incognita enhanced disease severity, as measured by the height of vascular browning in the stem, following inoculation of F. oxysporum f.sp. vasinfectum. The result also indicates the importance of timing of nematode infection and plant defense mechanisms for the establishment of an interaction as supported by increased disease symptoms when M. incognita are inoculated 10 days before or together with the FOL. The fungus might utilize the feeding tracts of the nematode to infect the tomato plants. Research findings of (Yang et al., 1976) indicated promoted wilt development viz. M. incognita and F. oxysporum f. sp. vasinfectum interaction on cotton. Highest wilt disease incidence due to infection of the nematode before the fungus on tomato plant (Khpalwak, 2012) is also proved, experimentally.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Al-Hazmi AS, Al-Nadary SN (2015) Interaction between Meloidogyne incognita and Rhizoctonia solani on Green Beans. Saudi Journal of Biological Sciences 22:570-574. |
|
|
Back M, Haydock P, Jenkinson P (2006). Interactions between the potato cyst nematode Globodera rostochiensis and diseases caused by Rhizoctonia solani AG3 in potatoes under field conditions. European Journal of Plant Pathology 114(2):215-223. |
|
|
Back MA, Haydock PP, Jenkinson P (2002). Disease complexes involving plant parasitic nematodes and soil borne pathogens. Plant Pathology 51(6):683-697. |
|
|
Bernard GC, Egnin M, Bonsi C (2017). The Impact of Plant-Parasitic Nematodes on Agriculture and Methods of Control. In: shah MM, Mohamood M., eds., Nematology Concepts, Characteristics and Control, InTech, London 121. |
|
|
Chen ZX, Chen SY, Dichson DW (2004). Nematology Advances and Perspectives: Nematode Management and Utilization. Tsinghua University Press, Beijing. |
|
|
Conroy JJ, Green RJ (1974). The Stubby Root Nematode Trichodorus christiei with Verticillium. Phytopathology 64:1118-1121. |
|
|
Coyne DL (2007). Practical plant nematology: a field and laboratory guide. IITA |
|
|
Central Statistical Agency (CSA) (2017). Agricultural sample survey report on area and production of major crops (private peasant holdings, Meher season). Vol I. Addis Ababa, Ethiopia. |
|
|
FAOSTAT (2016). Crop production: Food and Agricultural Organization of United Nations. |
|
|
Goswami BK, Agrawal DK (1978). Interrelationships between species of Fusarium and root knot nematode, Meloidogyne incognita, in Soybean. Nematologia Mediterranea 6(6):125-128. |
|
|
Holbrook CC, Knauft DA, Dickson DW (1983). A technique for screening peanut for resistance to Meloidogyne arenaria. Plant Disease 67(9):957-958. |
|
|
Hussey RS (1973). A comparison of methods of collecting inocula of Meloidogyne spp., including a new technique. Plant Disease Report 57:1025-1028. |
|
|
Johnson AW, Littrell RH (1970). Pathogenicity of Pythium aphanidermatum to chrysanthemum in combined inoculations with Belonolaimus longicaudatus or Meloidogyne incognita. Journal of Nematology 2(3):255. |
|
|
Jones JT, Haegeman A, Danchin EG, Gaur HS, Helder J, Jones MG, Kikuchi T, Manzanillaâ€López R, Palomaresâ€Rius JE, Wesemael WM, Perry RN (2013). Top 10 Plantâ€Parasitic Nematodes In Molecular Plant Pathology. Molecular Plant Pathology 14(9):946-961. |
|
|
Katsantonis D, Hillocks RJ, Gowen S (2003). Comparative effect of root-knot nematode on severity of Verticillium and Fusarium wilt in cotton. Phytoparasitica 31(2):154-162. |
|
|
Khpalwak W (2012). Interaction between Fusarium oxysporum f.sp. lycopersici and Meloidogyne incognita in tomato. Dissertation, University of Agricultural Sciences, Dharwad. |
|
|
Kumar B (2008). Studies on root Knot and wilt complex in Coleus forskohlii (Wild.) Briq. Caused by Meloidogyne incognita (Kofoid and White) Chitwood and Fusarium chlamydosporum (Frag. and Cif.) Booth. Doctoral dissertation, University of Agricultural Sciences, Dharwad. |
|
|
Kumar N, Bhatt J, Sharma RL (2017). Interaction between Meloidogyne incognita with Fusarium oxysporum f. sp. lycpersici on Tomato. International Journal of Current Microbiology and Applied Sciences 6:1770-1776. |
|
|
Leslie JF, Summerell BA (2006). The Fusarium Laboratory Manual. Blackwell Publishing, Iowa. |
|
|
Lobna H, Hajer R, Naima MB, Najet HR (2016). Studies on disease complex incidence of Meloidogyne javanica and Fusarium oxysporum f. sp. lycopersici on resistant and susceptible tomato cultivars. Journal of Agricultural Science and Food Technology 2(4):41-48. |
|
|
Mai WF, Abawi GS (1987). Interactions among root-knot nematodes and Fusarium wilt fungi on host plants. Annual Review of Phytopathology 25(1):317-338. |
|
|
McGovern RJ (2015). Management of tomato diseases caused by Fusarium oxysporum. Crop Protection 73:78-92. |
|
|
MoARD (Mintery of Agriculture and Rural development) (2009). Improved production technology of tomatoes in Ethiopia. |
|
|
Negron JA, Acosta N (1989). The Fusarium oxysporum f. sp. coffeae-Meloidogyne incognita complex in 'Bourbon'coffee. Nematropica 19(2):161-8. |
|
|
Oduor TK (2016). Agro-morphological and nutritional characterization of tomato landraces (lycopersicon species) in Africa. MSc Thesis, University of Nairobi. |
|
|
Pauline KM (2016). Host resistance and interaction between root knot nematodes and fusarium wilt of tomato. MSc Thesis, Kenya University. |
|
|
Seid A, Fininsa C, Mekete TM, Decraemer W, Wesemael WM (2017). Resistance screening of breeding lines and commercial tomato cultivars for Meloidogyne incognita and M. javanica populations (Nematoda) from Ethiopia. Euphytica 213(4):97. |
|
|
Shaner G, Finney RE (1977). The effect of nitrogen fertilization on the expression of slow-mildewing resistance in Knox wheat. Phytopathology 67(8):1051-6. |
|
|
Song W, Zhou L, Yang C, Cao X, Zhang L, Liu X (2004). Tomato Fusarium wilt and its chemical control strategies in a hydroponic system. Crop Protection 23(3):243-247. |
|
|
Srivastava R, Sharma SK, Singh JP, Sharma AK (2009). Evaluation of tomato (Solanum lycopersicum L.) germplasms against Fusarium oxysporum f. sp. lycopersici, causing wilt. Pantnagar Journal of Research (India). |
|
|
Tabor G, Yesuf M (2012). Mapping the current knowledge of carrot cultivation in Ethiopia. Carrot Aid, Denmark, pp. 1-20. |
|
|
Taylor AL, Sasser JN (1978). Biology, Identification and Control of Root-Knot Nematodes. International Nematology Project, North Carolina State University, Graphics, Raleigh 111. |
|
|
Van Gundy SD, Kirkpatrick JD, Golden J (1977). The nature and role of metabolic leakage from root-knot nematode galls and infection by Rhizoctonia solani. Journal of Nematology 9(2):113. |
|
|
Wang EL, Bergeson GB (1974). Biochemical changes in root exudate and xylem sap of tomato plants infected with Meloidogyne incognita. Journal of Nematology 6(4):194. |
|
|
Webster JM (1985). Interaction of Meloidogyne with fungi on crop plants. AGRIS: International information system for the agricultural science and technology |
|
|
Wondimeneh T, Sakhuja PK, Tadele T (2013). Root-knot nematode (Meloidogyne incognita) management using botanicals in tomato (Lycopersicon esculentum). Academia Journal of Agricultural Research 1(1):009-016. |
|
|
Yang H, Powell NT, Barker KR (1976). Interactions of concomitant species of nematodes and Fusarium oxysporum f. sp. vasinfectum on cotton. Journal of Nematology 8(1):74. |
|
|
Zahid MI, Gurr GM, Nikandrow A, Hodda M, Fulkerson WJ, Nicol HI (2002). Effects of root- and stolon-infecting fungi on root-colonizing nematodes of white clover. Plant Pathology 51:242-250. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


