Full Length Research Paper

ABSTRACT
Ectomycorrhizal fungi inoculation can increase the sustainability of planted forests hence the inoculant production should be optimized. The objective of this study was to determine the agitation speed and time of growth for maximum production of mycelium of D216 isolate of Pisolithus sp. The treatments were established by factorial 6x3, being six agitation conditions and three different times (days) of growth. The kinds of agitation used were: Without agitation, gentle manual agitation every two days and orbital agitation speed of 50, 100, 200 and 250 rpm. The times of growth were 14, 21 and 28 days. The mycelium mass was generally single and remained on the surface of the culture medium when grown without agitation or with manual agitation. Spherical mass units formed under agitation and grew submerged. The dry mass production of mycelium of D216 isolate was higher when grown under agitation of 200 rpm, followed by 250 and 100 rpm. For all agitation system, the production of mycelium did not increase after 14 days. The maximum production of mycelium dry mass of D216 isolate of Pisolithus sp. was obtained in the agitation speed of 200 rpm for 14 days. The elevation of the agitation to 250 rpm decreased the mycelium dry mass production of Pisolithus sp. In the absence or in lower agitation speeds, the mycelium dry mass was minimal. Using the ideal speed for maximum mycelium production and adjusting other growing conditions can decrease the risk of contamination, the production time and cost of the inoculant and encourage the use of this biotechnology.
Key words: Ectomycorrhizal fungi, inoculation, mycorrhiza, planted forests.
INTRODUCTION
The ectomycorrhizal fungi (EMF) are well known due to the association ability with planted trees species, such as eucalyptus and pines, increasing the survival and growth of the same (Chen et al., 2006; Quoreshi et al., 2008; Rivero et al., 2009; Costa et al., 2015, 2019). The inoculation of these trees with selected ectomycorrhizal fungi is a biotech strategy that can increase productivity in the forestry sector and reduce the excessive use of chemical fertilizers, which contribute to the high production cost and environmental impacts (Alberton et al., 2014; Costa et al., 2019). In ectomycorrhizae, fungi increase the plants ability to absorb water and nutrients and to withstand biotic and abiotic stresses and plants provides carbohydrates to the fungi (Garbaye, 2000; Chalot et al., 2002).
Inoculation of selected ectomycorrhizal fungi can reduce the use of fertilizers by up to 33% and increase coniferous seedlings growth, thus contributing to the reduction of environmental impacts associated with the production and use of chemical fertilizers in nurseries (Khasa et al., 2001). The beneficial effect of inoculation of ectomycorrhizal fungi in nursery was also observed in Eucalyptus seedlings (Chen et al., 2006; Costa et al., 2019), as well as the improvement in the quality and performance of transplanted seedlings (Quoreshi et al., 2008; Rivero et al., 2009). Inoculation of Pisolithus microcarpus isolate UFSC-Pt116 in Eucalyptus cutting increased plant growth by 13.1% at commercial nursery (Costa et al., 2019). Furthermore, seedlings of Pinus radiata inoculated with Rhizopogon roseolus and Scleroderma citrinum (Ortega et al., 2004) and seedlings of Pinus pinea inoculated with Rhizopogon luteolus and Rhizopogon roseolus (Parladé et al., 2004) survived and grew more than those not inoculated after planting in the field. Despite these results, the lack of investment by both government agencies and companies in the forestry sector limits the production and use of ectomycorrhizal inoculants in commercial nurseries in Brazil (Rossi et al., 2007).
The mycelium is the most appropriate way for these fungi inoculation (Brundrett et al., 1996), hence it must be used directly in in vitro tests, in selection tests in a greenhouse and commercially in the nursey seedlings inoculation (Alves et al., 2001). Thus, for its large-scale use, it is necessary to produce mycelium from pure cultures in culture medium (Pokojska et al., 1996).
Based on the assumption that the ectomycorrhizal fungus requires contact with nutrients and oxygen to grow, and considering the possibility of biotechnological application of biomass produced and the various uses of mycelium, the cultivation in a liquid culture medium is the most suitable for the mycelium biomass production (Rosado et al., 2003; RepáÄ, 2011). This cultivation can also be done in less space and less time when compared with the solid medium (Martin, 1983; Papagianni, 2004; Albaek et al., 2011). In addition, it offers ease of mycelium separation from the culture medium, allows the growth medium sterilization, maintenance of the aseptic culture and greater cultivation conditions control during the production process (Guillén-Navarro et al., 1998).
The physical effects of shaking speed during the fungi cultivation in a liquid culture medium require a great deal of attention. There are several reports that the agitation speed can change the morphology, differentiation of filamentous species and the time of growth and mycelium production of the fungi (Park et al., 2002b; Kim et al., 2003; Kelly et al., 2004; El-Enshasy et al., 2006). The agitation is important for the homogenization of liquid culture medium, mixing of nutrients, mass transfer and heat between the different phases present in the culture and maintenance of physical and chemical conditions suitable for mycelium production (Mantzouridou et al., 2002; Gupta et al., 2003). Furthermore, it is directly linked to the aeration rate of the culture medium, which may be beneficial for the growth and performance of the cells (Park et al., 2002a; Kim et al., 2003).
On the other hand, the exposure of the mycelium to high agitation speeds create shear forces, which undermine the fungi in several ways, reducing the growth and, so, the mycelium biomass production (Park et al., 2002b; Gupta et al., 2003; Kelly et al., 2004; El-Enshasy et al., 2006). However, there are also reports that the increase of agitation speed increases mycelium production (Cui et al., 1997; Kim et al., 2003). Although the fungi responses to unrest have been quite varied, the general trend for both the agitation and aeration is a positive correlation with the mycelium production (Albaek et al., 2011).
The use of the ideal agitation speed for the growth of each fungus isolate can increase mycelium production in less time and thus reduce production costs (Amanullah et al., 2000; Albaek et al., 2011). Among the ectomycorrhizal fungi, the species of the Pisolithus are among the most studied regarding the mycorrhizae formation (Barros et al., 1978; Campos et al., 2011; Alberton et al., 2014). The influence of agitation speed at the growth rate and mycelium formation should be examined, with the purpose of finding which agitation speed results in increased mycelium production in less time, causing less damage to the fungal cells. In this context, the objective of this study was to determine the agitation speed and the time of growth for maximum production of mycelium of D216 isolate of Pisolithus sp. in liquid culture medium.
MATERIALS AND METHODS
Location of experiment and fungal isolate
The study was carried out in the Laboratory of Soil Microbiology from the Federal University of Jequitinhonha and Mucuri Valleys - UFVJM, in Diamantina, Minas Gerais, Brazil (with headquarters at 18.20°S and 43.57°W).
The D216 isolate of the Pisolithus sp. used isolate that was obtained from EMF collection of the Laboratory of Soil Microbiology – UFVJM. Pure isolate cultures were originally obtained from basidiomas sampled in Eucalyptus spp. plantations in Jequitinhonha Valley – Minas Gerais. It was selected by presenting, in previous experiments, satisfactory growth in solid and liquid culture medium and by promoting benefits to eucalyptus plants in vitro tests (Costa et al., 2015) and in the nursery (Gomes, 2016).
Isolate growth, experimental design and treatments
The isolate was grown in Petri dishes (∅= 100 mm) with 20 mL of solid MNM culture medium for 29 days at 25ºC. Subsequently, discs of 5 mm diameter were removed from the edges of the isolate colonies, transferred to Petri dishes and kept for three more days to allow the damaged mycelium recovery during transplanting, confirm the feasibility, demonstrate the absence of contamination and perform the pre-growth for cultivation in liquid medium.
The experiment was carried out in a completely randomized design and treatments were established by factorial 6x3, being six agitation conditions and three different times (days) of growth. The agitations used were: without agitation, gentle manual agitation for three seconds every two days and orbital agitation speed of 50, 100, 200 and 250 rpm. The times of growth were 14, 21 and 28 days. The experimental unit was composed of one Erlenmeyer containing 50 mL of liquid Melin-Norkrans modified culture medium (MNM), pH 6,5 (Marx, 1969) inoculated with 10 discs of 5-mm diameter culture medium with pre-grown mycelium of D216 isolate of Pisolithus sp. and incubated at 25°C. Each treatment had eight repetitions, totaling 144 experimental units. The MNM culture medium was added of 20 mg L-1 of chloramphenicol to prevent bacterial contamination.
Determination of the mycelium dry mass and data analysis
To each time of cultivation, the fungal mycelium originated from each Erlenmeyer was collected in a sieve with a mesh of 53 μm, washed with distilled water. At the time of washing, the mycelium was visually analyzed for density, branching and fragmentation. The material was transferred to 30 mL plastic containers previously weighed and dried in an oven at 60ºC for three days until constant weight (Brundrett et al., 1986). Then, the set was weighed on an analytical balance (Marte, AY220). The mycelium dry mass was determined by the difference between the final weight of the plastic container with dried mycelium and the initial weight of the empty plastic container and was expressed in milligrams by 50 mL of culture medium (experimental unit).
Dried mass of mycelium was analyzed for distribution (Lilliefors test) and homogeneity (Cochran and Bartlett test). Then, using the Sisvar software, dates were subjected to two-way ANOVA and the means compared by the Tukey test (5% significance).
RESULTS
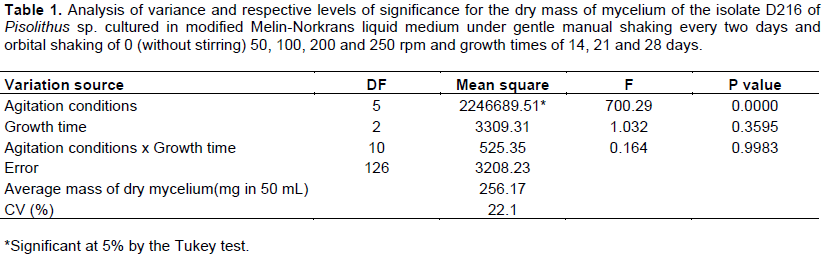
The agitation conditions influenced the mycelium dry mass production of D216 isolate of Pisolithus sp., however, the agitation with time interaction was not significant (Table 1).
The mycelia morphology, mycelium density or branching frequency, of the D216 isolate was visually different in the agitation condition evaluated. When grown without agitation or manual agitation some of the mycelium discs placed in the medium did not grow, the mycelium mass (densely interwoven mycelium masses referred to here as pellet) was generally single and remained on the surface of the culture medium.
Differently, under agitation, several spherical pellets units were formed and these grew submerged. At the end of the cultivation, these pellets differed in size and mycelium density; at a speed of 50 rpm, they were smaller and some fragmented hyphae filaments also occurred dispersed. At the speed of 100 rpm, mycelia pellets were larger than the previous ones and they had a hairy hyphal layer. At the speed of 200 rpm, the mycelia pellets were larger than the previous ones and had a more mycelium density with a more branching frequency. At the speed of 250 rpm, the mycelia pellets were smaller, irregular, and there were more hyphae fragmented.
The dry mass production of mycelium of D216 isolate was higher when grown under agitation of 200 rpm (747 mg), followed by 250 rpm (510 mg) and 100 rpm (157 mg) (Figure 1). For all agitation conditions, the production of mycelium did not increase after 14 days. The maximum production of mycelium dry mass (757 mg) of D216 isolate of Pisolithus sp. was obtained in the agitation speed of 200 rpm at 14 days of growth, while in the absence or low agitation speed the mycelium mass production was not affected (Figure 1).
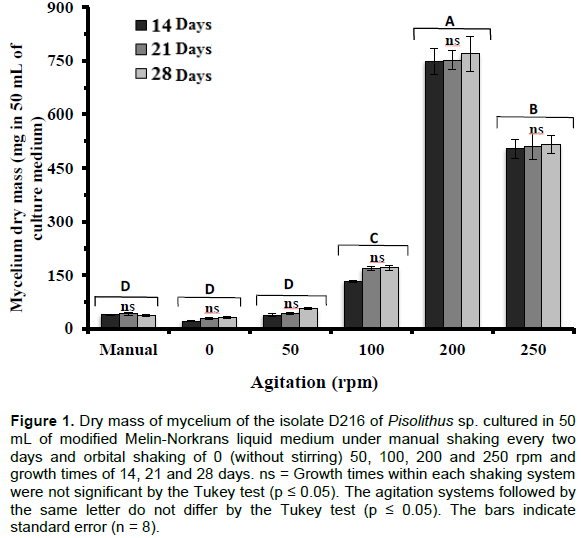
Elevation of the agitation speed from 200 to 250 rpm resulted in a decrease in the mycelium production of Pisolithus sp. (Figure 1).
DISCUSSION
The influence of agitation speed on the morphology of filamentous fungi growing in submerged liquid culture medium was also observed for Blakeslea trispora (Mantzouridou et al., 2002), Cordyceps militaris (Park et al., 2002b), Paecilomyces sinclairii (Kim et al., 2003) and Aspergillus niger (Kelly et al., 2004). This effect of the agitation speed on the fungi morphology confirms that these are morphologically complex organisms, differing in structure at different times in their life cycle, differing in form between surface and submerged growth, differing also with the nature of the growth medium and physical environment (Papagianni, 2004). Filamentous fungi growing in low agitation speeds, in general, produce larger pellets and lower mycelium dry mass, probably due to the lower nutrient availability in the internal parts of the pellet (El-Enshasy et al., 2006).
Regardless of the agitation conditions, there was dry mass production of the D216 isolate of Pisolithus sp. until the 14 days of growth (Figure 1). The insignificant growth of mycelium after 14 days of cultivation may be due to changes in the pH of the culture medium or depletion of some nutrient. This higher growth rate until the 14 days is better than what is cited in the literature for Pisolithus sp., which in general,is grown between 20 and 30 days for the inoculant production (Fernandes et al., 2014; Costa et al., 2015). For other faster growth fungi, as A. niger and Ceriporiopsis subvermispora, the greater quantity of mycelium dry mass is produced in a shorter time, from four to five days (Gupta et al., 2003; Kelly et al., 2004). Those authors proposed that the reduction in the growth rate was due to the depletion of the glucose.
The growth of the D216 isolate of Pisolithus sp. was minimal when incubated without agitation, agitated manually every two days or under agitation speed of 50 rpm even grown for 28 days (Figure 1). At a speed of 50 rpm, they were smaller and some fragmented hyphae filaments also occurred dispersed. This means that the same requires agitation for its greater growth in liquid medium. The lack of homogeneity of the liquid culture medium may have interfered with the aeration and distribution rate of nutrients, hindering the mycelium growth and consequently the dry mass production of Pisolithus sp. The agitation speed influences the nutrient availability and aeration rates, which reflects, in the mycelium morphology and increases the hyphae length, and consequently also the mycelium dry mass production (Amanullah et al., 1999, 2000; Park et al., 2002a, b; Mantzouridou et al., 2002; Gupta et al., 2003; Kelly et al., 2004; El-Enshasy, 2006). The agitation results in better culture medium mixing and maintains the concentration gradient between the cells’ interior and exterior. This gradient promotes the satisfactory supply of sugars and other nutrients to the cells, besides facilitating the removal of gases and other catabolism byproducts from the cellular microenvironment, resulting in greater mycelium dry mass production (Mantzouridou et al., 2002).
The agitation speed increase for 100 and 200 rpm increased the visible number of mycelium pellets and mycelium dry mass of Pisolithus sp., reaching maximum production at a speed of 200 rpm (Figure 1). At the speed of 200 rpm, the mycelia pellets were larger than the previous ones and had a more mycelium density and seemingly more branching frequency. The 747 mg of dry mycelium mass obtained in this work at a speed of 200 rpm was 3.9 times greater than that observed for Laccaria laccata grown in 100 mL of liquid Lamb’s medium inoculated with one 10-mm in diameter disc without agitation (Pokojska et al., 1996) and was 21 times greater than that observed for the ectomycorrhizal fungi Cadophora finlandia in solid MNM modified (Azaiez et al., 2018). This effect of increased agitation speed was also observed for the pathogenic fungus B. trispora (Mantzouridou et al., 2002) and the parasitic fungus C. militaris (Park et al., 2002b). The authors’ proposed that increase of mycelium dry mass was due to better air supply to the cells. Generally, aerobic fungi have an increased growth rate when moderate aeration is combined with an increase in the agitation speed (Mantzouridou et al., 2002).
With the additional increase of the agitation speed to 250 rpm, the mycelia pellets were smaller, irregular, and there were more hyphae fragments and decreased mycelium dry mass production (Figure 1). Although, for other fungi, Aspergillus awamori (Cui et al., 1997) and P. sinclairii (Kim et al., 2003), the agitation speed of 250 rpm increased the mycelium dry mass production. The increase in the number of pellets under higher agitation speeds was also observed in A. niger growing at speeds of 200 to 800 rpm (El-Enshasy et al., 2006). Most of the time, agitation speeds lead to greater energy dissipation, linked to high shear risk, which can result in cell fragmentation and damage, decreasing the mycelium production (Park et al., 2002b; Gupta et al., 2003; Kelly et al., 2004; El-Enshasy et al., 2006). In addition, it can increase the culture medium’s apparent viscosity, due to the increase in the quantities of free filaments, causing the reduction of the efficiency in mass transfer and limitingoxygen and nutrient absorption by the cells (Kelly et al., 2004).
The results obtained in this study confirmed that agitation speed is an important parameter in the mycelium development and dry mass production of Pisolithus sp., and indicates the agitation speed of 200 rpm as the ideal for growth and production of mycelium dry mass of 216 isolate of Pisolithus sp. These results corroborate with those found in literature, where it is reported that in appropriate aeration conditions, the optimal agitation speed for mycelium mass production in general is within the range of 150 to 250 rpm (Papagianni et al., 2001; Mantzouridou et al., 2002; Park et al., 2002a; Kim et al., 2003; El-Enshasy et al., 2006). The agitation speed within this range provides a balance between the forces of cohesion and disintegration; besides allowing the formation of small pellets favorable to a higher mycelium dry mass production during prolonged cultivation (Papagianni et al., 2004; El-Enshasy et al., 2006).
CONCLUSION
The mycelium dry mass production of D216 isolate of Pisolithus sp. is maximum at 14 days of growth under 200 rpm of agitation. Using the ideal speed for maximum mycelium production and adjusting other growing conditions can decrease the risk of contamination, the production time and cost of the inoculant and encourage the use of this biotechnology.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors appreciate the Universidade Federal dos Vales do Jequitinhonha e Mucuri (UFVJM) for analyses of performance, and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) for financial support.
REFERENCES
|
Albaek MO, Gernaey KV, Hansen MS, Stocks MS (2011). Modeling enzyme production with Aspergillus oryzae in pilot scale vessels with different agitation, aeration, and agitator types. Biotechnology Bioengineering 108(8):1828-1840. |
|
|
Alberton O, Aguiar D, Gimenes MRT, Carrenho R (2014). Meta-analysis for responses of eucalyptus and pine inoculated with ectomycorrhizal fungi in Brazil. Food Agriculture Environment Journal 12(2):1159-1163. |
|
|
Alves JR, Souza O, Podlech PAS, Giachini AJ, Oliveira VL (2001). Efeito de inoculante ectomicorrízico produzido por fermentação semi-sólida no crescimento de Eucalyptus dunnii Maiden. Pesquisa Agropecuária Brasileira 39(4):307-313. |
|
|
Amanullah A, Blair R, Nienow AW, Thomas CR (1999). Effects of agitation intensity on mycelial morphology and protein production in chemostat cultures of recombinant Aspergillus oryzae. Biotechnology Bioengineering 62(4):434-446. |
|
|
Amanullah A, Jüsten P, Davies A, Paul GC, Nienow AW, Thomas CR (2000). Agitation induced mycelial fragmentation of Aspergillus oryzae and Penicillium chrysogenum. Biotechnology Bioengineering 5(2):109-114. |
|
|
Azaiez A, Nadeau MB, Bertrand A, Khasa DP (2018). In vitro selection of ecologically adapted ectomycorrhizal fungi through production of fungal biomass and metabolites for use in reclamation of biotite mine tailings. Mycologia 110(6):1017-1032. |
|
|
Barros NF, Brandi RM, REIS MS (1978). Micorriza em eucalipto. Uma revisão sobre a morfologia, a fisiologia e os efeitos mútuos da associação fungo-planta. Revista Árvore 2(2):130-140. |
|
|
Brundrett M, Bougher N, Dell B, Grove T, Malajczuk, N (1996). Working with Mycorrhizas in Forestry and Agriculture. Canberra: ACIAR Monograph 32 - Australian Centre for International Agricultural Research 374 p. |
|
|
Campos DTS, Silva MCS, Luz JMR, Telesfora RJ, Maria CMK (2011). Colonização micorrízica em plantios de eucalipto. Revista Árvore 35(5):965-974. |
|
|
Chalot M, Javelle A, Blaudez D, Lambilliote R, Cooke R, Sentenac H, Wipf D, Botton B (2002). An update on nutrient transport processes in ectomycorrhizas. Plant and Soil 244(2):165-175. |
|
|
Chen YL, Kang LH, Malajczuk N, Dell B (2006). Selecting ectomycorrhizal fungi for inoculating plantations in South China: effect of Scleroderma on colonization and growth of exotic Eucalyptus globulus, E. urophylla, Pinuselliottii, and P. radiata. Mycorrhiza. New Phytologist 16(4):251-259. |
|
|
Costa LS, Grazziotti PH, Grazziotti DCFS, Silva AC, Rossi MJ, Silva EB, Costa VHD, Gomes ALF (2015). In vitro evaluation of Eucalyptus ectomycorrhizae on substrate with phosphorus doses for fungal pre-selection. Revista Árvore 39(1):127-136. |
|
|
Costa LS, Grazziotti PH, Silva AC, Fonseca AJ, Gomes ALF, Grazziotti DCFS, Rossi MJ (2019). Alginate gel entrapped ectomycorrhizal inoculum promoted growth of cuttings of Eucalyptus clones under nursery conditions. Canadian Journal of Forest Research 48(8):978-985. |
|
|
Cui YQ, Van der Lans RGJM, Luyben KCAM (1997). Effect of agitation intensities on fungal morphology of submerged fermentation. Biotechnology Bioengineering 55(5):15-725. |
|
|
El-Enshasy H, Kleine J, Rinas U (2006). Agitation effects on morphology and protein productive fractions of filamentous and pelleted growth forms of recombinant Aspergillus niger. Process Biochemistry 41(10):2103-2112. |
|
|
Fernandes MCS, Costa LS, Grazziotti PH, Grazziotti DCFS, Santos JB, Rossi MJ (2014). Pisolithus sp. tolerance to glyphosate and isoxaflutole in vitro. Revista Árvore 38 (3):461-468. |
|
|
Garbaye J (2000). The role of ectomycorrhizal symbiosis in the resistance of forest to water stress. Outlook on Agriculture 29(1):63-69. |
|
|
Guillén-Navarro GK, Márquez-Rocha FJ, Sanchez-Vázquez JE (1998). Producción de biomasa y enzimas ligninolíticas por Pleurotus ostreatus en cultivo sumergido. Revista Ibero americana de Micología 15:302-306. |
|
|
Gupta R, Gigras P, Mohapatra H, Goswami VK, Chauhan B (2003). Microbialα-amylases: biotechnological perspective. Process Biochemistry 38(11):1599-1616. |
|
|
Kelly S, Grimm LH, Hengstler J, Schultheis E, Krull R, Hempel DC (2004). Agitation effects on submerged growth and product formation of Aspergillus niger. Bioprocess and Biosystems Engineering 26:315-323. |
|
|
Khasa PD, Sigle L, Chakravarty P, Dancik BP, Erickson L, Mc Curdy D (2001). Effect of fertilization on growth and ectomycorrhizal development of container-grown and bare-root nursery conifer seedlings. New Forests 22:179-197. |
|
|
Kim SW, Hwang HJ, Xu CP, Choi JW, Yun JW (2003). Effect of aeration and agitation on the production of mycelial biomass and exopolysaccharides in an enthomopathogenic fungus Paecilomyces sinclairii. Letters Applied Microbiology 36(5):321-326. |
|
|
Mantzouridou F, Roukas T, Kotzekidou P (2002). Effect of the aeration rate and agitation speedon -carotene production and morphology of Blakesleatrisporain a stirred tank reactor: mathematical modeling. Biochemical Engineering Journal 10(2):123-135. |
|
|
Martin AM (1983). Submerged production of edible mushroom mycelium. Canadian Institute Food Science Technology Journal 16(3):215-217. |
|
|
Marx DH (1969). The influence of ectotrophicmycorrhyzal fungi on the resistence of pine roost to pathogenic infections and antagonism of mycorrhizal to root pathogenic fungi and soil bacteria. Phytopathology 59(2):153-163. |
|
|
Ortega U, Duñabeitia M, Menendez S, Gonzalez-Murua C, Majada J (2004). Effectiveness of mycorrhizal inoculation in the nursery on growth and water relation of Pinus radiata in different water regimes. Tree Physiology 24(1):65-73. |
|
|
Papagianni M, Nokes SE, Filer K (2001). Submerged and solid-state phytase fermentation by Aspergillus niger: Effects of agitation and medium viscosity on phytase production, fungal morphology and inoculum performance. Food Technolology and Biotechnology 39(4):319- 326. |
|
|
Papagianni M (2004). Fungal morphology and metabolite production in submerged mycelial processes. Biotechnology Advances 22(3):189-259. |
|
|
Park JP, Kim YM, Kim SW, Hwang HJ, Cho YJ, Lee YS, Song CH, Yun JW (2002a). Effect of aeration rate on the mycelial morphology and exo-biopolymer production in Cordyceps militaris. Process Biochemistry 37:1257-1262. |
|
|
Park JP, Kim YM, Kim SW, Hwang HJ, Cho YJ, Lee YS, Song CH, Yun JW (2002b). Effect of agitation intensity on the exo-biopolymer production and mycelial morphology in Cordyceps militaris. Letters Applied Microbiology 34(6):433-438. |
|
|
Parladé J, Luque J, Pera J, Ricón AM (2004). Field performance of Pinuspinea and P. halepensis seedlings inoculated with Rhizopogon spp. and outplanted in formerly arable land. Annals Forest Science 61(6):507-514. |
|
|
Pokojska A, Kampert M, Różycki H, Strzelczyk E (1996). Effects of vitamins, temperature and pH on the biomass production by ectomycorrhizal fungi. Acta Mycologica 31(1):55-65. |
|
|
Quoreshi AM, Piché Y, Khasa DP (2008). Field performance of conifer and hardwood species 5 years after nursery inoculation in the Canadian Prairie Provinces. New Forest 35(3):235-253. |
|
|
RepáÄ I (2011). Ectomycorrhizal inoculum and inoculation techniques. In. Rai M, Varma A (Eds.), Diversity and Biotechnology of Ectomycorrhizae: Soil Biology, pp. 43-63. |
|
|
Rivero SHT, Moorillón VGN, Borunda EO (2009). Growth, yield, and nutrient status of pecans fertilized with biosolids and inoculated with rizosphere fungi. Bioresource Technology 100(6):1992-1998. |
|
|
Rosado FR, Germano S, Carbonero ER, Costa SMG, Iacomini M, Kemmelmeier CD (2003). Biomass and exopolysaccharide production in submerged cultures of Pleurotus ostreatoroseus Sing. and Pleurotus ostreatus "florida" (Jack.: Fr.) Kummer. Journal Basic Microbiology 43(3):230-237. |
|
|
Rossi MJ, FurigoJr A, Oliveira VL (2007). Inoculant production of ectomycorrhizal fungi by solid and submerged fermentations. Food Technology Biotechnology 45(3):277-286. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


