Full Length Research Paper

ABSTRACT
INTRODUCTION
Solenostemon rotundifolius [(Poir.) J. K. Morton] (Lamiaceae), is commonly called Chinese potato, Innala, Hausa potato, Zulu round potato, Sudan potato, Saluga, Fabirama or Frafra potato. It is believed to have originated from Central or East Africa but spread throughout tropical Africa and into South-east Asia. It is cultivated as tuber crop in many African countries including Burkina Faso, Ghana, Nigeria, Togo, Mali (in west Africa), Cameroon and Chad (in central Africa) and some parts of South and East Africa (Schippers, 2000; Gouado et al., 2003; Sugri et al., 2013). In Asia, S. rotundifolius is reported to be cultivated in Sri Lanka, South India and Java (Jayakody et al., 2005).
S. rotundifolius is an annual herbaceous plant, 15 to 30 cm high, with ascending or prostrate stems and thick leaves having aromatic smell (Sugri et al., 2013). It is specially adapted to the Sahelian region of West Africa. Local varieties of S. rotundifolius produce many (up to 70/plant) small sized tubers; 3.78 cm long and 1.53 cm width (Nanéma et al., 2009). The potential yield reported in West Africa ranged from 7 to 20 T/ha (Enyiukwu et al., 2014). The tubers contain significant rate of reducing sugar (26 mg/100g), protein (13.6 to 14.6 mg/100g), crude fat (1.2%), crude fiber (1.6%), phosphorus (36 mg/100g), calcium (29 mg/100g), vitamins A and C, respectively 13.6 mg/100g and 10.3 mg/100g, and antioxidants (Anbuselvi and Balamurugan, 2013; Anbuselvi and Priya, 2013). They are commonly consumed as a curry, baked or fried, or cooked (Agyeno et al., 2014).
Besides its nutritional attributes, S. rotundifolius holds strong economic potential and could be financially rewarding to the farm economy (Enyiukwu et al., 2014). During the period of availability, the quantity of tubers sold varied from 16 to 32 Kg/day/person in the main markets of Ouagadougou in Burkina Faso. According to the period and the tubers availability, the prices varied from 1.2 to 3 USD/Kg. This important variation of prices (250%) showed the economic potential of frafra potato (Nanéma et al., 2017).
S. rotundifolius is a very important food crop which can contribute to improving food security. Besides its agricultural importance, it has ornamental, medicinal, culinary and many other uses (Kwarteng et al., 2017). Therefore, as mentioned by Chivenge et al. (2015) and Mamadalieva et al. (2017), there are many neglected crops that have the potential to contribute to food security but investigation should be done to clearly demonstrate their potentialities and the priorities in term of research on these crops. It is now a consensus that understanding variability within plant genetic resources is one of the key steps for a sustainable use and conservation of its potentialities. Some research activities were already carried out on S. rotundifolius germplasm and contributed to identify a set of useful traits that could be used as descriptors for this crop (Opoku-Agyeman et al., 2007; Nanéma et al., 2009). Some other works focused on the identification of the main morphotypes within S. rotundifolius genetic resources based on tuber skin color (Prematilake, 2005; Sugri et al., 2013).
A description based on one trait could lead to some confusions between the morphotypes. The objective of this study is to contribute to a better description of the main morphotypes of S. rotundifolius cultivated in Burkina Faso based on a set of qualitative morphological traits. This will give some useful tools for the identification of the morphotypes for research activities on agronomic, economic or nutritional potentialities of S. rotundifolius.
MATERIALS AND METHODS
Plant material
Three accessions (E02, E35 and E20) representing three morphotypes of S. rotundifolius (respectively A, B and C) were used for this study (Table 1). These accessions were identified based on morphological variation observed during our previous research on S. rotundifolius genetic resources. The accession E02 was collected in the province of Passoré in the North region of Burkina Faso. The accessions E35 and E20 were collected in the province of Nahouri in the South region of Burkina Faso. For each accession, 30 tubers were randomly selected for the experiment (Table 1).

Study area and experimental design
The research was carried out in the research farm of the Faculty of Earth and Plant Sciences of the University Ouaga I Joseph KI-ZERBO in Ouagadougou (12°21′56″ N; 1°32′01″ W). A total rainfall of 665.1 mm was registered during the period of the experiment (July 2016 to January 2017). The experiment was laid out in Fisher blocks with three replications. The replication consisted in three lines of height plastic buckets perforated at the bottom to improve drainage. Each plastic buckets (9 L) contained a mix of sun (1/3) and potting soil (2/3). The spacing was 50 cm between the lines, 40 cm between the plants and 50 cm between the blocks. One tuber was planted per bucket on 25 July 2016. An additional irrigation was bring after the rainy season from October 2016 to January 2017.
Morphological traits
A total of 25 qualitative morphological traits were assessed at different stages of plant growth. These traits were identified during our previous research activities. The different variants were identified and scored per plant. At all, the morphological traits were observed on 24 plants per morphotype.
Morphological traits observed at vegetative stage
Height morphological traits were observed during the vegetative stage. The color of stems (COSt) and the color of leaves (COL1) of young plant were scored at juvenile stage (Table 2). During the plant growth, the foliage color (FCO), the thickness of leaves (TLE), and the color of apical leaves (CAL) were observed. The variation of leaves color when injured by insects (LCI), the main stem (SMS) and the secondary stems (SSS) stature were observed before flowering period.

Morphological traits observed at flowering stage and after harvesting
A total of seventeen morphological traits were observed on the inflorescence, the stems and the tubers after flowering. Nine of them were observed on the inflorescence. These traits were the number of flower buds per cluster (NFB), the color of flower buds (CFB), the type of inflorescence (TIN), the level of branching of inflorescence (LBI), the arrangement of clusters of flowers on the rachis (AFR) and the flowers persistence on the rachis (FPE). The color of sepals (COSe), and the color of petals (CPE) were observed on three randomly selected flowers per plant. The color of rachis (CRA) was also observed. Besides the morphological traits observed on the inflorescence, two were observed on the stems and the leaves. These were the shape of the section of main stems (SSS) and the color of leaves of adult plant (COL2). Six traits were observed on the tubers. After harvesting, the position of tuberous roots (PTR), the tuber shape (TSH), the tuber skin color (TSC), the tuber flesh color (TFC), the presence of lateral tubers (PLT) and the texture of tuber skin (TTS) (Table 2).
Data analysis
The individuals with missing data were eliminated. A consensus value for each morphological trait was identified based on the individual score of each plant. The morphotypes were then described based on the consensus value for all the morphological traits.
RESULTS
Variability of the morphotypes at juvenile stage
A variability was observed between young plant of the morphotypes for stems and leaves color (Table 3). The stems and leaves of young plants of morphotypes “A” and “C” were green (Figures 1 and 2) while the young plants of morphotype “B” were red (Figures 3 and 4).




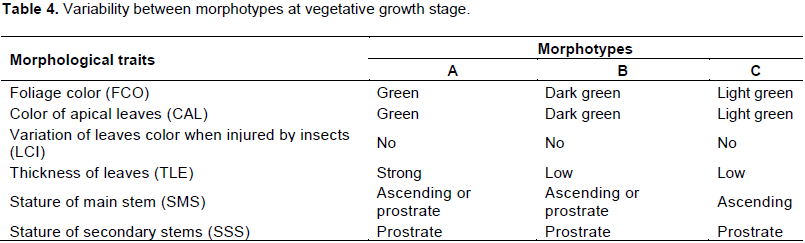
Variability of the morphotypes at vegetative growth stage
Variability was observed between the morphotypes at vegetative stage (Table 4). The color of foliage was green for morphotype “A”, dark green for morphotypes “B” and light green for morphotype “C”. The color of apical leaves did not vary from the color of whole foliage. No particular color was observed on the leaves when injured by insects. The morphotypes also varied for the level of leaves thickness. The leaves of the morphotype “A” were very thick while those of the morphotypes “B” and “C” were relatively less thick (Figure 5).

Variability of the morphotypes at flowering stage
The morphological traits observed at flowering stage revealed important variability between the morphotypes. The inflorescence was an apical spike. Some inflorescences with primary branching were observed for the morphotypes “A” and “B” (Table 5). The flower buds color was green with reddish pigmentation for the morphotype “B” (Figure 6) but it was green for the morphotypes “A” (Figure 7) and “C” (Figure 8). The number of flower buds per cluster was four for the morphotypes “A” and “C” but it was three for the morphotype “B”. The clusters of flowers were opposite on the rachis for all the morphotypes. The flowers buds and the flowers were no persistent on the rachis. After flowering, the color of rachis was reddish for the morphotype “A” but was green for the morphotypes “B” and “C”. The morphotype “A” developed elongated flowers with green-reddish sepals and white-purple petals (Figure 9). The sepals of the flowers of the morphotype “B” was reddish and the petals were very purple (Figure 10). For the morphotype “C”, the sepals were green and the petals were reddish (Figure 11). The morphological traits observed on the stems and the leaves after flowering did not vary. The section of stems was quadrangular for all the morphotypes and the leaves were dark green.




Variability of the morphotypes for the morphology of the tubers
The tuber skin and flesh color, and the tuber shape were the discriminant traits observed on tubers (Table 6). The tubers of the morphotype “A” were black skin (Figure 12) and the flesh color was white (Figure 13). The tubers of the morphotype B, skin color was red (Figure 14) and the tuber flesh color was white-yellow (Figure 15). The tubers of the morphotype “C” skin color was white-yellow (Figure 16) and the flesh color was yellow (Figure 17). Tubers were oblong for morphotypes “A” and ovoid for the morphotypes “B” and “C”. For all the morphotypes, the tuberous roots were observed on the upper parts of the roots. The tubers were rough and some tubers presented lateral tubers.



DISCUSSION
Three morphotypes of S. rotundifolius were described based on morphological traits observed on leaves, stems, inflorescences and tubers. At juvenile stage, the presence red coloration on young plant was identified to be the main difference between the morphotypes. The variation of young plant color was also observed by Nanéma et al. (2009) as a discriminant traits within S. rotundifolius genetic resources. According to Price and Sturgess (1938), the reddish color observed on Lamiaceae species are due to the presence of anthocyanins (mainly cyanidine saccharides).
During the vegetative growth stage, the main variation was observed for leaves thickness and the stature of main stem. All the morphotypes produced thick leaves. As suggested by Edison et al. (2006), leaves morphology is an important parameter for water regulation in plant tissues. The strong thickness of leaves of the morphotype “A” could be considered as a particular adaptation potential of this morphotype to water scarcity conditions. Besides the leaves morphology, the stature of stems was identified as a discriminant parameter. Previous research activities revealed the presence of pigmentation on S. rotundifolius leaves (Agyeno et al., 2014).
The most significant discriminant traits were observed on the inflorescence. These parameters can be sufficient to identify the morphotypes. Our previous works on S. rotundifolius variability mentioned the color of the petals of the morphotypes “A” and “B” (Nanéma et al., 2009). This work is the first report on the flowering of the morphotype “C”. The other morphological traits observed after flowering did not reveal differences between the morphotypes. It was the stem section, that was quadrangular and the color of leaves that was green. The quadrangular stem section of S. rotundifolius was also mentioned by Agyeno et al. (2014). However, variability of stem section can be observed within the same morphotype (Nanéma et al., 2009). A pigmentation on leaves was also mentioned at flowering stage. These traits seem to be influenced by S. rotundifolius growth conditions.
In addition to the tuber skin color, a variability was observed for tuber flesh color. Current literature mentioned three morphotypes according to the tuber skin color: - the morphotype with white skin tuber; the morphotype with red skin tuber; and the morphotype with black or brown skin tuber (Prematilake, 2005; Nanéma, 2010; Sugri et al., 2013). Chevalier and Perrot (1905) mentioned these morphotypes respectively as varieties “alba”, “rubra” and “nigra”. These authors suggested the presence of a fourth variety with black skin tuber called “javanicaminum”. Some research activities were carried out on S. rotundifolius based on the tuber skin color as the main criteria of identification of the varieties (Jayakody et al., 2005; Priya and Anbuselvi, 2013, Taye et al., 2012). After harvest, the tuber skin color is the most accessible criteria for identification of the morphotypes but any error of color appreciation could lead to ambiguous results. Additional morphological traits could be the tuber flesh color and tuber shape.
The tubers of all the morphotypes were rough and glabrous. Other research activities in West Africa also mentioned the lack of hairiness on tubers of S. rotundifolius (Opoku-Agyeman et al., 2007; Agyeno et al., 2014).
In Asia, some varieties of S. rotundifolius produce hairy tubers (Jayakody et al., 2005). This difference could be due to genetic factor or to growth conditions. For all the morphotypes, many branched tubers were identified. Solenostemon rotundifolius produced small size tubers and the presence of branched tubers make peeling very difficult. According to Enyiukwu et al. (2014), branched tubers make the crop unattractive and reduce the marketability of the tubers.
The described morphotypes were identified in S. rotundifolius germplasm in Burkina Faso. Based on some morphological traits used in previous works, the three morphotypes were reported to be cultivated in West Africa (Sugri et al., 2013; Agyeno et al., 2014). The morphotype “A” and “B” are cultivated in Sri Lanka respectively as “Bola” and “Dik” (Prematilake, 2005, Jayakody et al., 2005). Some research activities highlighted the influence of genetic variability on tuber nutritional, medicinal potentialities and their behavior in conservation (Jayakody et al., 2005; Priya and Anbuselvi, 2013; Parmar et al., 2017; Mamadalieva et al., 2017; Azad et al., 2017). A clear classification of accessions used in future research on S. rotundifolius could contribute to a better analysis of the results. This include nutritional, agronomical and economical potentialities as well as genetic diversity.
Future research activities could also be focused on the existence of variability within the morphotypes. Other aspects could be the description of flower morphology. Similar studies already led to the revision of Borassus classification (Bayton et al., 2006).
CONCLUSION
This study described three morphotypes of S. rotundifolius cultivated in Burkina Faso based on a set of qualitative morphological traits. These morphological traits can be used as descriptors for S. rotundifolius. It is an important step towards full description of the morphotypes of this important tuber crop. The future studies on morphological variability could focused on flower structure and microscopic description of plant tissues. A complete description of the morphotypes will provide useful information for a sustainable management of genetic resources of S. rotundifolius.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
This work was supported by the University Ouaga I Pr Joseph KI-ZERBO.
REFERENCES
|
Agyeno OE, Jayeola AA, Ajala BA, Mamman BJ (2014). Exo-morphology of vegetative parts support the combination of Solenostemon rotundifolius (Poir) J. K. Morton with Plectranthus esculentus N. E. Br. Natal (Lamiaceae) with insight into infra-specific variability. Advances in Agriculture and Botanics-International Journal of the Bioflux Society 6(1):16-25. |
|
|
Anbuselvi S, Balamurugan T (2013). Nutritional and antinutrional consituents of Manihot esculentus and Plectranthus rotundifolius. International Research Journal of Pharmacy 4(9):97-99. |
|
|
Anbuselvi S, Priya HM (2013). Nutritional and anti-nutritional constituents of Plectranthus rotundifolius. International Journal of Pharmaceutical Sciences Review and Research 22(1):213-215. |
|
|
Azad AK, Kabir H, Eaton TEJ, Soren EB (2017). Storage potentialities of some exotic potato varieties at farmers' condition in Bangladesh. Agricultural Sciences 8(2):183-193. |
|
|
Bayton RP, Ouédraogo A, Guinko S (2006). The genus Borassus (Arecaceae) in West Africa, with a description of a new species from Burkina Faso. Botanical Journal of the Linnean Society 150:419-427. |
|
|
Chevalier A, Perrot E (1905). Les Coleus à tubercules alimentaires. In : Les végétaux utiles de l'Afrique Tropicale française. Etudes scientifiques et agronomiques, Fascicule I, Paris 1:100-152. |
|
|
Chivenge P, Mabhaudhi T, Modi AT, Mafongoya P (2015). The potential role of neglected and underutilised crop species as future crops under water scarce conditions in sub-saharan Africa. International Journal of Environmental Research and Public Health 12(6):5685-5711. |
|
|
Edison S, Unnikrishnan M, Vimala B, Pillai SV, Sheela MN, Sreekumari MT, Abraham K (2006). Biodiversity of tropical tuber crops in India national biodiversity authority. National Biodiversity Authority Chennai, Tamil Nadu, India 31 p. |
|
|
Enyiukwu DN, Awurum AN, Nwaneri JA (2014). Potentials of Hausa Potato (Solenostemon rotundifolius (Poir.) JK Morton and Management of its tuber rot in Nigeria. Greener Journal of Agronomy, Forestry and Horticulture 2(2):027-037. |
|
|
Gouado I, Fotso M, Djampou EJ (2003). Potentiel nutritionnel de deux tubercules (Coleus rotundifolius et Solenostemon ssp.) consommés au Cameroun. 2ème atelier international, voies alimentaires d'amélioration des situations nutritionnelles, Ouagadougou 13-28/11/2003, pp. 85-90. |
|
|
Jayakody L, Hoover R, Liu Q, Weber E (2005). Studies on tuber and root starches. I. Structure and physicochemical properties of innala (Solenostemon rotundifolius) starches grown in Sri Lanka. Food Research International 38(6):615-629. |
|
|
Kwarteng AO, Ghunney T, Adu Amoah R, Nyadanu D, Abogoom J, Nyam KC, Ziyaaba JZ, Danso EO, Whyte T, Asiedu DD (2017). Current knowledge and breeding avenues to improve upon Frafra potato (Solenostemon rotundifolius (Poir.) J. K. Morton). Genetic Resources and Crop Evolution 65(2):1-11. |
|
|
Mamadalieva NZ, Akramov DK, Ovidi E, Tiezzi A, Nahar L, Azimova SS, Sarker SD (2017). Aromatic medicinal plants of the Lamiaceae family from uzbekistan: ethnopharmacology, essential oils composition, and biological activities. Medicines 4(8):1-13 |
|
|
Nanéma KR, Traoré RE, Bationo/Kando P, Zongo JD (2009). Morphoagronomical characterization of Solenostemon rotundifolius (Poir J. K. Morton) (Lamiaceae) germplasm from Burkina Faso. International Journal of Biological and Chemical Sciences 3(5):1100-1113. |
|
|
Nanéma KR (2010). Ressources génétiques de Solenostemon rotundifolius (Poir.) JK Morton du Burkina Faso : Système de culture, variabilité agromorphologique et relations phylogénétiques entre ses différents morphotypes cultivés au Burkina Faso. Thèse de Doctorat Unique, Univ. Ouaga., Burkina Faso. 122p. |
|
|
Nanéma KR, Sawadogo N, Traoré ER, Ba AH (2017). Marketing potentialities and constraints for frafra potato : case of the main markets of Ouagadougou (Burkina Faso). Journal of Plant Science 5(6):191–195. |
|
|
Opoku-Agyeman MO, Bennett-Lartey SO, Vodouhe RS, OSEI C, Quarcoo E, Boateng SK, Osekere EA (2007). Morphological characterization of frafra potato (Solenostemon rotundifolius) germplasm from the savannah regions of Ghana. Plant genetic resources and food security in West and Central Africa. Regional Conference, Ibadan, Nigeria, 26-30 April, 2004, pp. 116-123. |
|
|
Parmar A, Gebre BA, Legesse A, Demelash Y, Fladung K, Hensel O (2017). Nutritional comparison of white and red Coccinia abyssinica (Lam.) Cong. accessions: an under-utilised edible tuber of the ethiopian highlands. Foods 6(8):71. |
|
|
Prematilake DP (2005). Inducing genetic variation of innala (S. rotundifolius) via in vitro callus culture. Journal of the National Science Foundation of Sri Lanka 33(2):123-131. |
|
|
Price JR, Sturgess VC (1938). A survey of anthocyanins. VI. Biochemical Journal 32(9):1658-1660. |
|
|
Priya HM, Anbuselvi S (2013). Physico chemical analysis of Plectranthus rotundifolius. Journal of Chemical and Pharmaceutical Research 5(3):12-14. |
|
|
Schippers RR (2000). African indigenous vegetables. An overview of the cultivated species. Natural Resources Institute/ACP-EU Technical Centre for Agricultural and Rural Cooperation, Chatham, United Kingdom 214 p. |
|
|
Sugri I, Kusi F, Kanton LAR, Nutsugah KS, Zakaria M (2013). Sustaining frafra potato (Solenostemon rotundifolius Poir.) in food chain; current opportunities in Ghana. Journal of Plant Sciences 1(4):68-75. |
|
|
Taye M, Lommen MWJ, Struik PC (2012). Ontogeny of the tuber crop Plectranthus edulis (Lamiaceae). African Journal of Agricultural Research 7(30):4236-4249. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


