Full Length Research Paper

ABSTRACT
This work aims to identify the impacts of vermicompost leachate (VCL) toward its salt resistance stress in plants. Young Capsicum annuum L seedlings were subjected to salinity (160 mM NaCl added to a nutrient solution) over a course of seven days during or following the adding of 7 mL L−1 VCL. The exposure to salt stress resulted in a notable decrease of dry and fresh leaf weights, K+ concentrations, and leaf water content; whereas leaf and root Na+ concentrations were remarkably increased. Furthermore, the salt stress yielded reduced instantaneous transpiration, net photosynthesis, maximum PSII photochemistry efficiency, stomatal conductance (gs) in the state of photochemical quenching, dark adaptation, and photochemical efficiency of PSII (ΦPSII). Moreover, VCL application in the state of salt stress led to an increase of leaf fresh weight and gs, while the leaf osmotic potential remained consistent regardless of the proline content increase seen in the salt-treated plants. Meanwhile, it minimized the Na+ concentration in the leaves by 21.4%, and increased in the roots by 16.9%. Accordingly, a pre-treatment regiment with VCL and the ensuing salt stress yielded higher efficiency compared to its concomitant application with salt stress due to the pre-treatment recording the highest osmotic adjustment. VCL pre-treatment resulted in sugar concentration in the roots (9.3 µmol g−1 FW) and the maximum level of proline in the leaves (51 µmol g−1 FW). Additionally, the fluorescence-linked parameter reaffirmed superior outcomes for the PSII stability and efficiency in the case of VCL pre-treatment on salt-stressed plants in contrast to those subjected to concurrent VCL salt stress exposure. Hence, VCL is proven to be an effective defense medium towards enhancing the property of salt-stress resistance in bell peppers.
Key words: Vermicompost leachate (VCL), priming, bell pepper, Sodium, NaCl.
INTRODUCTION
Salinity and water deficiency are two major abiotic determinants (Wang et al., 2009; Vibhuti et al., 2015; of Bargali and Bargali, 2016) due to the high magnitude their impact and wide occurrence (Bartels and Sunkar, 2005). Salt stress is reported as a serious problem (Nedjimi and Daoud, 2006) which directly or indirectly affects the physiological status of an organism by altering its metabolism, growth and development (Jnandabhiram and Sailen, 2012) and adversely affects agricultural productivity (Shahi et al., 2015; Bartels and Sunkar, 2005).
Generally, the parameter of salinity is deemed a key limitation in today’s global agrarian industry. In particular, salt stress can be described in terms of two primary groups of physiological stress in plants; the first group constitutes the feature of NaCl increase causing a reduced water availability in plants, specifically due to external osmotic potential decrements (Benazzouk et al., 2018). Meanwhile, the second group consists of the induction of ionic stress following an increased NaCl concentration, which is attributable to the higher toxic ion absorption (e.g., Na+) and less essential element absorption (e.g., K+ and P). As a result, the property of plant salt-stress resistance is a direct reflection of their demand for a sustained water status via osmotic adjustment, thereby facilitating the turgidity maintenance while sustaining the K+/ Na+ selectivity and allowing the sequestration of vacuolar Na+. Collectively, they will hinder any toxic ion interference throughout the biochemical process undertakings occurring in the cell cytosol (Coelho et al., 2020). Normally, the species of the plant kingdom yields notable variations of their salt stress resistance.
Majority of the cultivated plants consist of glycophyte species, which are comparatively sensitive to this particular parameter (Wang et al., 2019). Furthermore, such property is highly accurate for bell peppers or Capsicum annuum L., which is otherwise one of the primarily ingested fruits globally (Erel et al., 2020). In fact, salt stress may potentially pose a highly detrimental influence on the bell pepper production. Moreover, the halophytic plant species are made up of several closely-related plant species for the Capsicum genus, thereby rendering interspecific crosses a potential alternative for salt-stress-resistant hybrids (Parađiković et al., 2019). Unfortunately, such goal is highly costly, time-consuming, and requires concerted efforts in order to choose the best material equipped with the required agrarian attributes (Erel et al., 2020). Besides, one may further opt for genetic engineering to carry out specific gene transfer; however, it comes with varying population-based issues and is less pragmatic as a solution. This is due to salt-stress resistance being a polygenic trait dependent upon the interactions of multiple genes (Saghafi et al., 2019). Additionally, genetically enhancing the cultivated material is yet another straightforward and cost-effective process towards optimizing the growth conditions for bell peppers, which are similarly deemed a great alternative for a short-term salt-stress resistance (Liu et al., 2019).
To this particular end, biostimulants are highly favorable as an option. The EBIC (European Biostimulants Industry Council) has stated that plant-based biostimulants have micro-organisms and/or substances capable of stimulating various natural processes when subjected to plants or the rhizosphere. As a result, the parameters of nutrient uptake and efficiency and plant stress tolerance are enhanced, thus yielding in quality crops. Besides, such products are accessible across different types, whereby a majority of them consist of an intricate amalgamation of various active substances (Hajizadeh et al., 2019). In particular, vermicompost is a form of eco-friendly organic fertilizer generally manufactured by earthworm actions, namely via organic waste digestion.
Decomposition of organic matter is enhanced by the microbes and earthworms which influence the nutrient cycling and enrich the soil (Bargali et al., 1993; Bargali, 1996; Bargali et al., 2015); it later becomes available for the plant uptake and could enhance the production of crops and make the soil system sustainable. The fertilizer is inherently robust due to its important nutrients, hormones, active humic substances, and phenolic compounds, thereby collectively yielding significant impacts on the plant physiology (Mohapatra et al., 2019; Moridi et al., 2019). Following earthworms digesting the organic matter, solid vermicompost is generated, as well as a liquid phase due to leaching processes. In bell peppers, the application of vermicompost is geared toward enhancing the plant growth, fruit production (Rorat and Vandenbulcke, 2019), and quality. Furthermore, it is proven to yield an improved security against root-knot nematode (Singh et al., 2010) and minimize water stresses and temperature in the seedlings (Piovan et al., 2019). Regardless of this, information detailing vermicompost influence on bell pepper resistance to salt stress remains inadequate (Kar et al., 2019). Accordingly, VCL’s capacity towards minimizing salt stress in bell peppers has been substantiated, specifically by highly effectual compatible solute build-up (e.g., soluble sugar and proline) and high concentrations of photosynthetic pigments.
Majority of such works implemented the product application concurrently with environmental limitation. Despite this, salt stress is not a particularly indefinite element in the context of natural field conditions; instead, it may present as an outcome of insufficient irrigation practices (Liu et al., 2019). Therefore, one should prioritize the determination of VCL application is akin to vermicompost implementation prior to the former’s exposure to salt stress and the latter’s concurrent salt stress outcomes. Hence, the research hypothesizes that VCL may be afforded a defense mechanism against salt stress in bell peppers in which its impacts vary according to the stress application timings. For the purpose of analyzing the possible ‘protective’ impacts of vermicompost on resistance of salt-stress in C. annuum, this work opts to assess the cultivation of young plants in a nutrient solution, whereby VCL is put in either prior or within salt stress. The plant growth is henceforth assessed in the context of ion concentration, plant water status regulations, and their photosynthetic attributes.
MATERIALS AND METHODS
Growth conditions, vermicompost leachate and plant materials
The seedlings of Capsicum annuum L. or bell peppers (cv. Romance, Rijk Zwaan) were subjected to germination in a greenhouse, whereby they were placed in the jars and implemented loam as the substrate. After 15 days, they were transferred to a growth chamber with a hydroponic culture system and subjected to a photoperiod of 16 h and under the light intensity (160 µmol m−2 s−1). The default temperatures were allotted to 26°C at daytime and 19°C at nighttime, whereas the humidity was set at 80 and 90%, respectively. All seedlings were allocated at the rate of six seedlings per tank into 12 tanks containing 1.4 L of aerating nutrient solutions placed in a thoroughly randomized block design. We secured these seedling, on the aerated hydroponic nutrient solution having polystyrene plates floating, which contains:1.5 mM (NH4)6Mo7O24, 0.75 mM MgSO4, 33 µM Fe-EDTA, 3 mM KH2PO4, 25 µM H3BO4, 6 µM MnSO4, 4 mM KNO3, 1.5 µM CuSO4, 1.5 mM Ca(NO3)0 and1.5 µM ZnSO4. The solution pH level was subjected to daily adjustment (5.9) and the solution itself was replaced every seven days.
In this study, the vermicompost utilized was manufactured in the greenhouse of the Ain Shams University located in Cairo, Egypt in 2018. This was done via food waste vermicomposting and by using Eisenia fetida, otherwise known as earthworms, in accordance with the suggestions of Ndegwa and Thompson (Ndegwa and Thompson, 2001). A prior work has shown that the VCL yield contains the following elements accordingly: anthocyanins (1.5 mg L−1), C-glycosides (90 mg L−1), K (46 mg L−1), humic acids (679 mg L−1), polyphenols (42 mg L−1) and P (4.8 mg L−1). Following a 7-day period of acclimatization, all plants in the 12 tanks were subjected to maintenance in the nutrient solution for seven days. However, four tanks received an additional VCL (7 mL L−1) for pretreatment, whereas the remaining eight tanks were subjected to either one of other prescribed treatments over the course of seven days. The treatments included: (1) nutrient solution and addition of 160 mM NaCl (2) nutrient solution and addition of 160 mM NaCl (for control of salt stress); (3) nutrient solution and the addition of 160 mM NaCl and 7 mL L−1 VCL both (4) 7 mL L−1 VCL plus nutrient solution (for control) and (5) nutrient solution only (as control). Concurrently, plants subjected to the VCL pre-treatment also went through either one of these treatments for the subsequent seven days, namely, or (6) nutrient solution only (as VCL pre-treated control). All of the treatments were subjected to every plant across two tanks.
Osmotic potential, plant growth and water status
Subsequent to the six treatment options, the roots and shoots of 4 arbitrarily selected plants per treatment were differentiated accordingly. Following the fresh (FW) determination, the roots and leaves were measured for their dry weight (DW) after a period of incubation of 48 h at 72°C. In terms of the osmotic potential parameter (Ψs), the samples were divided into multiple segmentations and put in the prepared microcentrifuge tube (MyFuge 5 MicroCentrifuge USA). These tubes were characterized with perforations, namely four small holes, and then placed within another microcentrifuge tube, which was intact. Then, the samples were subjected to three cycles of freeze/thaw and centrifugation at 15,000×g for 15 min at 4°C. Next, an analysis of the plant tissue sap was done to obtain the Ψs, whereas the osmolarity assessment was undertaken by using a vapor pressure osmometer (Osmomat 3000 Germany). Then in accordance with the Van’t Hoff equation: Ψs (MPa)=–c(mosmol kg−1)×2.58 10−3 was converted to MPa (Lutts et al., 1996).
Na+ and K+ contents determination
To this end, the root and leaf tissues of 4 plants for each treatment was first subjected to oven-drying at 71°C for a period of two days and then reduced to powder form via grinding in liquid nitrogen. Then, digestion of the samples occurred at 75°C and in 5 mL HNO3 34%, following which the remnants were dissolved again in aqua regia (64% HNO3:36% HCl; 1:3), then filtered (Whatman, 12 mm). Next, quantification of the ion concentration has been undertaken via FAAS (flame atomic absorption spectrophotometry) (LabGenius Single Beam 4nm UV/VIS Spectrophotometer USA).
Net photosynthesis, Stomatal conductance and Chlorophyll fluorescence
Measurements of the leaves stomatal conductivity (gs) were done by using the 4th completely expanded leaves of 6 leaves per treatment by utilizing a diffusion porometer (AP4 Leaf Porometer from Delta-T ALGETE Spain). They were undertaken between the timing of 13 and 15 h. Next, the rate of net carbon assimilation (A) was measured under intercellular CO2 concentration (ci) and 500 (µmol m−2 s−1) PPFD by utilizing the prior infrared gas analyzer system and in accordance with Vandoorne et al. (2012). Again, the measurements were taken among the timing of 11:00 and 13:00 h using 6 plants for each treatment.
Chlorophyll fluorescence quantification was undertaken by utilizing a portable pulse-modulated chlorophyll fluorimeter (Model OS30p+ - Handheld, USA) in which the values were obtained using the fourth leaf and performed using the middle section of the leaflet. The leaflets portions were then subjected to acclimatization in darkness for 20 min accordingly. First, the maximum fluorescence image (Fm) was identified with PSII reaction center at 8600 µmol m−2 s−1 using a 0.6-s saturating pulse. Then, the minimum fluorescence image (F0) was assessed via 0.2 (µmol m−2 s−1) modulated light measurements. The leaves were then illumined for 2 min using 230(μmol m− 2 s−1) white actinic light, while, Fs (steady state fluorescence) was recorded.
Statistical treatment
Data analysis was conducted via one-way analysis of variance (ANOVA), which allowed the determination of the primary impacts of the varying treatments undertaken in this work towards the physiological and biochemical plant parameters.
Determination of osmocompatible solute
First, the roots and leaves of 4 plants for each treatment have been subjected to powdering in N2 (liquid) and then kept in storage at −80°C until further analysis was undertaken. In particular, free proline quantification was done per the method suggested by Bates et al. (1973) and Adamipour et al. (2019); whereas soluble sugar extraction was undertaken per (26) before it was measured using the anthrone technique per (Yakhin et al., 2017).
RESULTS
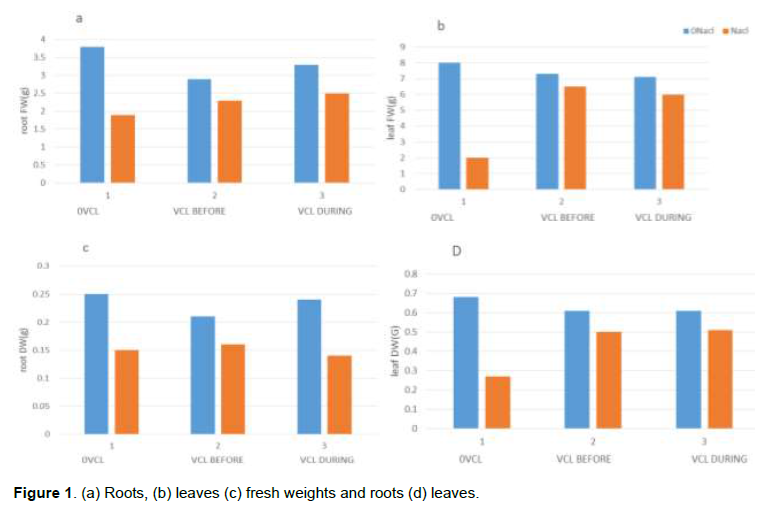
According to the findings, salt stress (that is, 160 mM NaCl) successfully triggered notable decrease in the root (Figure 1a) and leaf (Figure 1b) FWs in comparison with the untreated control (nutrient medium only). Meanwhile, when there is no salt stress, using VCL (7 mL L−1) has no effect on the parameters. Then, VCL application during and prior salt-stress hindered the leaf FW decrease, which was generally an outcome of salt stress (Figure 1b). Furthermore, salt stress yielded less significant effect on the root DW (Figure 1c) despite the notably reduced leaf DW (Figure. 1d). Upon repeated VCL application prior or during salt stress, the salt-stress-induced inhibitory effect was inhibited and reflected in the leaf growth. Both parameters of salt stress alone and VCL alone did not result in any notable impacts on the root WC. However, salt stress caused a notable decrease of leaf WC and the plants subjected to VCL either prior or during the experiment revealed nearly comparable plants WC to that of unstressed leaves. Moreover, as shown in Table 1, salt stress minimized the leaves and roots osmotic potentials (Ψs). Similarly, VCL resulted in a decrease of Ψs root in the salt-stressed plants, which was higher in extent for those treated with VCL prior to salt stress in comparison with the plants subjected to VCL only during salt stress. Additionally, the leaves Ψs value was less in VCL-treated plants prior to salt stress, whereas those subjected to VCL during salt stress yielded no significant leaf Ψs values. Per the initial expectations, consistently low root and leaf Na+ contents were noted when salt stress was absent, while VCL treatment yielded no impact on the particular parameter (Figure 2). However, salt stress resulted in an increase of the root Na+ amount (Figure 2a), whereas VCL treatment during or prior salt stress revealed contrasting impacts. Accordingly, the VCL pretreatment generated a notable reduction of the root Na+ concentration, whereas a concurrent VCL treatment yielded a significant increase for the parameter. Contrary to this, both VCL treatments resulted in a significant reduction of the Na+ build-up in the leaves of the salt-stressed plants (Figure 2b). Similarly, a slight decrease of the K+ concentration was noted in the leaves, which was absent in the plant roots (Figure 2c, d). Then, the salt-stressed plants subjected to VCL pretreatment showed an increase of K+ concentration in their leaves; whereas concurrent VCL treatment caused the K+ concentration in the roots to rise. As shown in Figure (3a, b), the proline content of the roots and leaves did not change during VCL treatment despite the lack of salt stress.
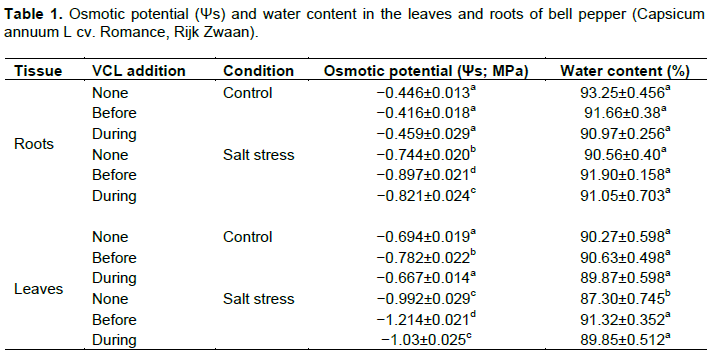

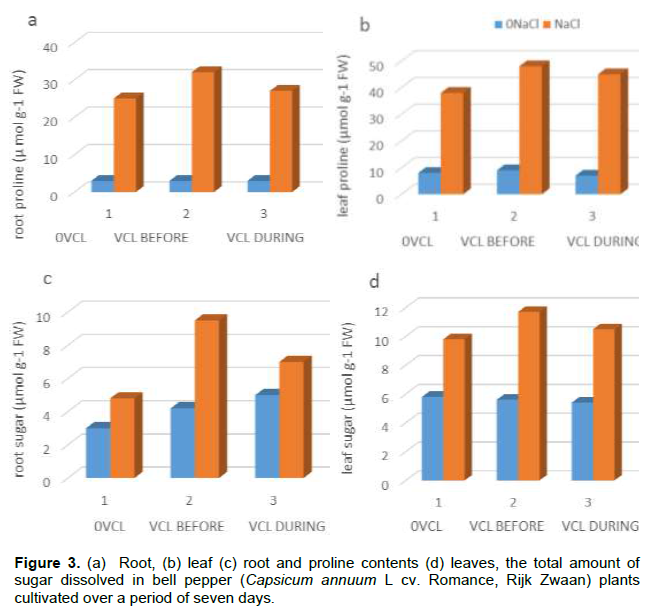
Accordingly, salt stress resulted in the leaves and roots proline content increase, whereby the increment values were higher for salt-stressed plant subjected to treatment by VCL. Similarly, VCL pre-treatment caused a bigger outcome for the leaf proline content under salt stress conditions compared to its concomitant application with salt stress (Figure 3b). In terms of the sugar content, the lack of salt stress rendered the VCL treatment not to affect the leaves but it increased root sugar content regardless. Here, the salt stress resulted in an increase of the root sugar content, particularly when subjected to VCL pre-treatment (Figure 3c). Contrary to this, as shown in Figure 3d, no impacts were seen on the leaf sugar content upon VCL treatment subjected in salt-stressed plants.
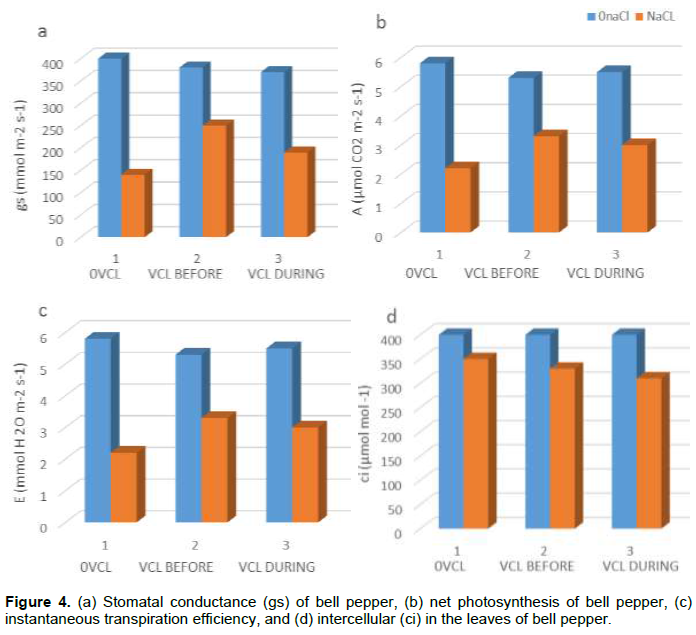
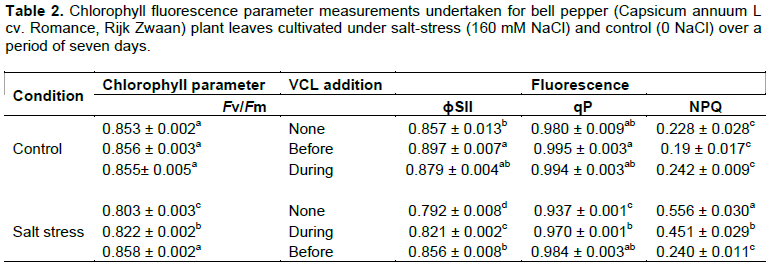
However, salt stress minimized the stomatal conductance (gs) (Figure 4a). VCL treatment caused a significant increase of gs in salt-stressed plants, whereby its effect was greater when undertaken pre-treatment rather than during concomitant application with salt-stress. Furthermore, salt stress minimized the instantaneous transpiration (E) and net photosynthesis (A) (Figure 4b, c), whereby a significant reduction of their salt stress-induced negative impacts was due to the VCL pre-treatment. As shown in Figure (4d), in plants that were cultivated without VCL treatment, salt stress did not affect the Ci (intercellular CO2 mole fraction), but in the VCL treated plants, it reduced Ci. Similarly, according to Table 2, the parameters that decreased in response to salt stress were photochemical quenching (qP) and the greatest amount of quantum yield for the dark acclimated leaves (Fv/Fm). Therefore, the VCL pretreatment posed a protective mechanism against the harmful effects of salt stress upon such parameters, whereas salt-stressed plants subjected to concurrent VCL treatment revealed an intermediate behavior. Merely the presence of salt stress could increase the non-photochemical quenching (NPQ), which could be minimized by VCL pre-treatment; whereas concomitant VCL treatment and salt stress resulted in less protective effect. Regardless, similar observations were made regarding the photochemical efficiency of photosystem II (ΦPSII): salt stress minimized its yield and remained high in VCL-pretreated plants in contrast to those treated with VCL and subjected to salt stress concomitantly.
The incoming few decades can be associated with the primary goal of improved marginal land productivity, particularly in agrarian areas impacted by salinity or drought. In such locations, the crop would have yielded comparably low output and optimal fertilization might potentially aid the plants in overcoming certain harmful effects of abiotic stresses. Unfortunately, specific localities in the world affected by such negative environmental conditions may be burdened by the inaccessibility to chemical fertilizers or small farms being unable to pay for their expensive cost. In addition to this, these products may pose a risk toward the ecosystem stability. Therefore, vermicomposting processes are currently highlighted as a potential alternative for such chemical fertilizers (Gupta et al., 2019). The eco-friendly product obtained via the post-thermophilic organic material biodegradation as a result of earthworm-microorganism interactions is particularly suited to be added into soil (Rorat and Vandenbulcke, 2019). Despite this, solid vermicompost also poses a big drawback if it is to be employed in salt-impacted areas, namely the risk of soil salinity increase as a result of its high electrical conductivity (Singh et al., 2010). Contrary to this, VCL is characterized by its electrical conductivity, thereby offering its advantageous homogeneity if applied in plant production (Bolong and Saad, 2020). Regardless, the implementation of vermicompost and VCL have been found to result in an improved bell pepper yield and quality alike, despite their usage across various systems, ranging from fields to greenhouses and hydroponic systems (J. M. P. H. E. R. and 2020 n.d.). Additionally, plenty of other plant species have been linked with similar data and evidence.
The current work showed the manner in which no apparent benefits were noted when VCL was applied in bell pepper plant cultivation with salt stress absence. In particular, the parameters of growth, ion contents, and photosynthesis-linked components were unaffected by such application, whereby merely a slight increase of the root sugar content was observed following VCL addition in non-salt-stressed plants. However, prior works have the capacity of vermicompost or VCL to yield a notable bell pepper plant growth stimulation when salt stress is absent, specifically in the context of the stem and root lengths, stem diameters and root and shoot FWs (Greco et al., 2019). In specific circumstances, these plants are cultivated in soil and any performance improvements are a result of the enhanced soil microbial population and soil structure (Morton et al., 2019). Recent works have further noted the varying differences of bell pepper growth performance due to VCL, which is highly reliant upon the type of soil utilized. This worked commenced by having all plants fixed and maintained in the selected nutrient solution, thereby rendering any VCL effects on the aggregates size, soil structure and water holding capacity of the substrate to be irrelevant to the discussion. Liu et al. (2019)have further revealed VCL’s role in improving P and K deficiencies seen in the bell pepper seedlings. This work opted to maintain the unstressed seedlings in an optimal nutrient situation capable of providing all components and elements to ensure their growth and development. As a result, the plants did now show any form of nutrient deficiency. Furthermore, VCL and its influence on the fruit size and quality have been delineated in various works, which implement vermicompost over a long duration spanning from the seedling phase to the reproductive phase. However, this work emphasized the seedling phase only, opting for a short VCL exposure duration (seven days). Similarly, Benazzouk et al. (2020) have described the effect of VCL to be different according to the bell pepper cultivars being sample, whereas have denoted its composition variability to be due to the organic waste characteristics following their usage in the composting processes (Bolong and Saad, 2020). These prior works have underlined the manner in which varied VCL preparations may possibly influence the bell pepper plant growth differently. Such differences across the culture system, sampled cultivar, and VCL composition may potentially delineate the reason behind the lack of apparent VCL impact on the seedlings seen in this work, which occurs when salt stress is absent.
Here, the situation was noted to be different for the plants subjected to salt-stress. In particular, VCL application prior or through salt-stress successfully diminished certain harmful markers of salt stress, leading to significant leaf FW and DW decrease, as well as minimized leaf WC and highly increased root and leaf Na+ concentrations. Apart from the reduced K+ concentration, it also yielded a decrease of the gs, ΦPSII, A, Fv/Fm, E, and qP, values, whereas the NPQ rose. Moreover, the VCL application through salt-stress eliminated the leaf FW decrease, which was triggered by the salt stress, thereby confirming its beneficial influence on the salt-stress resistance in bell peppers (Javaid et al., 2019). Such protective mechanism has also been noted in the case of sunflower and peppermint (Jabeen and Ahmad, 2017). Moreover, this work revealed the improved leaf WC values following the application of VCL during salt stress, thereby substantiating its capacity to minimize the water stress element of salt stress. Regardless, such outcome cannot be attributed to stomatal closure as its application led to an increased gs as opposed to its decrease. Therefore, it has been proposed its capability to enhance the osmotic adjustment in case of water-stressed circumstances in the context of proline and soluble sugar content build-up. According to Shelke et al. (2019), VCL application during salt stress triggers a hike in proline content in contrast to plants exposed to salt stress in its absence, but such increase is inadequate to cause a significant leaf Ψs decrease. Contrary to this, VCL-treated roots concurrently subjected to salt stress resulted in less Ψs compared to those subjected to salt stress in its absence, which may be partially attributable towards the soluble sugar accumulation.
Moreover, the biochemical foundation of VCL-induced increase for the proline content is yet to be ascertained but it has been noted when stress is absent in the bell peppers (Bulgari et al., 2019a). Regardless of the increased glutamine synthase (EC 6.3.1.2) and glutamate dehydrogenase (EC 1.4.1.2) activities thus significantly affecting proline precursor synthesis in salt-stressed plants, Ezzat et al., 2019)have substantiated the lack of effect exerted by vermicompost on this enzymatic activity in salt-stressed sunflowers. Here, have reported on vermicompost’s protective mechanism on Mentha haplocalyx and Silybum marianum against salt stress in which their respective proline content is diminished as a result of global salt stress intensity reduction (Benazzouk et al., 2019).
Furthermore, the monovalent cation absorption and root-to-shoot translocation may be an alternative goal in the context of VCL-induced advantages salt-stressed plants. Here, the application minimized the leaf Na+ concentration, while the root Na+ content was increased. Therefore, this indicates its capacity to minimize Na+ translocation to the aerial components of the plant, thereby being an effective approach for salt-stress resistance in the bell pepper (Javaid et al., 2019). Contrary to this, VCL-treated plants subjected to salt stress yielded an increased root K+ concentration, whereas it showed a slight decrease in salt-stressed plants when VCL was absent. VCL is made up of K, resulting in its common usage as an ‘NPK’ fertilizer (Singh et al., 2010; Pandyaswargo and Premakumara, 2014). Regardless, the root K+ content noted in the VCL-treated plants showed an increase alongside salt stress, which was not seen when VCL was absent. Therefore, this suggests the interaction between salt stress and VCL towards improving the K+ nutrition. Moreover, its inherent humic substances may interact with membrane transporters, which has been reported previously, as well as with H+-ATPases (Schiavon et al., 2019) and nitrate Additionally, these humic substances may be capable of stimulating the gene expression, specifically those encoding two putative maize nitrate transporters (du Jardin et al., 2020). Therefore, VCL interaction with K+ transporters should not be dismissed, despite NaCl-induced salt stress and its effect on such interaction possibly necessitating future investigation.
DECISION
It should be noted that the VCL application prior to salt stress yielded better efficiency than its concomitant use with salt stress in the context of salt-stress resistance improvements. As shown in Table 1, Such preventive mechanism resulted in the best osmotic adjustment values towards the end of the stress period. Furthermore, both root and shoot Na+ concentrations were reduced, suggesting Na+ absorption was subjected to a modification under such conditions, as opposed to Na+ translocation. Besides, such protective nature yielded a higher proline and K+ concentration in the leaves, while the roots depicted the highest level of sugar concentration. Collectively, this influenced the fluorescence-related parameters and emphasized the VCL-treated salt-stressed plants that resulted in greater efficiency and PSII stability upon their exposure to salt stress as opposed to those subjected to VCL and salt stress concurrently. Furthermore, the application of VCL prior to salt stress caused the stomatal to open during the period of salt stress, which would maintain net photosynthesis and gaseous exchange without causing any harmful effect on the plant water status. Such status is noted to be effectively controlled via osmotic adjustment. Even though VCL application without salt stress present did not trigger any morphological adjustments, it was shown to alter the physiological status of the plants, causing them to adapt to the salt stress in a better and effectual manner. Therefore, it may be viewed as an effectual protective element capable of reinforcing plant resistance against salt stress, whereby such modifications and their underpinning mechanisms can be attributed to VCL properties. Additionally, humic acid extraction from earthworm compost has been recorded to show auxin properties, while indole acetic acid can congregate by the supramolecular arrangement of humic acid (Hajizadeh et al., 2019). Despite certain works have noted abscisic acid to be present in vermicompost, Bulgari et al. (2019b) have stated its level may be reduced by humic acids. Similarly, its endogenous level may be prevented by humic acids as well. In fact, the cytokinin increased linked with a reduced ABA may delineate the sustained high gs outcomes in salt-stressed tissues (Bulgari et al., 2019b). Therefore, one may hypothesize that VCL exposure prior to salt stress can cause high stomatal conductance, thereby allowing the transpiration stream maintenance. As a result, these plants stockpile on the protective phytohormones or antioxidants from the VCL, which will be inherently present upon salt stress exposure. Contrary to this, concurrent contact with salt stress and VCL present the stomatal conductance to be sustained, resulting in its closure due to low external osmotic potential. Following this, putative protective compound translocation and the transpiration stream will be reduced.
CONCLUSION
VCL application on the bell pepper plants was found to partially eliminate the harmful effects of salt stress on their growth and photosynthesis functions, which was correlated with the minimized Na+ accumulation. If it was applied prior to salt stress, the outcomes were better and more effectual compared to its concomitant application with salt stress. Therefore, VCL may be implemented as an effectual protective component towards enhancing the element of salt-stress resistance in bell peppers. Currently, additional works are in the works in order to test such hypothesis, which are undertaken by conducting a thorough hormonal profiling of VCL-treated plants and VCL both, under salt-stress and control circumstances.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Adamipour N, Khosh-Khui M, Salehi H, Rho H (2019). Effect of vermicompost on morphological and physiological performances of pot marigold (Calendula officinalis L.) under salinity conditions. Advances in Horticultural Science 33(3):345-358. |
|
|
Bargali K, Bargali S (2016). Germination capacity of seeds of leguminous plants under water deficit conditions: implication for restoration of degraded lands in Kumaun Himalaya. Tropical Ecology 57(3):445-4. |
|
|
Bargali S, Shukla K, Singh L, Ghosh L, Lakhera M (2015). Leaf litter decomposition and nutrient dynamics in four tree species of dry deciduous forest. Tropical Ecology 56(2):191-200. |
|
|
Bargali S, Singh S, Singh R (1993). Patterns of weight loss and nutrient release from decomposing leaf litter in an age series of eucalypt plantations. Soil Biology and Biochemistry 25(12):1731-8. |
|
|
Bargali SS (1996). Weight loss and N release in decomposing wood litter in a eucalypt plantation age series. Soil Biology and Biochemistry 28(4-5):699-702. |
|
|
Bartels D, Sunkar R (2005). Drought and salt tolerance in plants. Critical Reviews in Plant Sciences 24(1):23-58. |
|
|
Bates LS, Waldren RP, Teare I (1973). Rapid determination of free proline for water-stress studies. Plant and Soil 39(1):205-207. |
|
|
Benazzouk S, Djazouli ZE, Lutts S (2018). Assessment of the preventive effect of vermicompost on salinity resistance in tomato (Solanum lycopersicum cv. Ailsa Craig). Acta Physiologiae Plantarum 40(6):121. |
|
|
Benazzouk S, Djazouli ZE, Lutts S (2019). Vermicompost leachate as a promising agent for priming and rejuvenation of salt-treated germinating seeds in Brassica napus. Communications in Soil Science and Plant Analysis 50(11):1344-57. |
|
|
Benazzouk S, Dobrev PI, Djazouli ZE, Motyka V, Lutts S (2020). Positive impact of vermicompost leachate on salt stress resistance in tomato (Solanum lycopersicum L.) at the seedling stage: a phytohormonal approach. Plant and Soil 446(1):145-62. |
|
|
Bolong N, Saad I (2020). Characterization of University Residential and Canteen Solid Waste for Composting and Vermicomposting Development. Green Engineering for Campus Sustainability: Springer pp. 193-206. |
|
|
Bulgari R, Franzoni G, Ferrante A (2019a). Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 9(6):306. |
|
|
Bulgari R, Trivellini A, Ferrante A (2019b). Effects of two doses of organic extract-based biostimulant on greenhouse lettuce grown under increasing NaCl concentrations. Frontiers in Plant Science 9:1870. |
|
|
Coelho DG, Miranda RS, Paula-Marinho SO, de Carvalho HH, Prisco JT, Gomes-Filho E (2020). Ammonium nutrition modulates K+ and N uptake, transport and accumulation during salt stress acclimation of sorghum plants. Archives of Agronomy and Soil Science pp. 1-14. |
|
|
du Jardin P, Xu L, Geelen D (2020). Agricultural Functions and Action Mechanisms of Plant Biostimulants (PBs). The Chemical Biology of Plant Biostimulants pp. 1-30. |
|
|
Erel R, T Le T, Eshel A, Cohen S, Offenbach R, Strijker T, Shtein I (2020). Root Development of Bell Pepper (Capsicum annuum L.) as Affected by Water Salinity and Sink Strength. Plants 9(1):35. |
|
|
Ezzat K, Pernemalm M, Pålsson S, Roberts TC, Järver P, Dondalska A, Bestas B, Sobkowiak MJ, Levänen B, Sköld M, Thompson EA, Saher O, Kari OK, Lajunen T, Ekström ES, Nilsson C, Ishchenko Y, Malm T, Wood MJA, Power UF, Masich S, Lindén A, Sandberg JK, Lehtiö J, Spetz AL, EL Andaloussi S (2019). The viral protein corona directs viral pathogenesis and amyloid aggregation. Nature Communications 10:2331. |
|
|
Greco C, Comparetti A, Mammano MM, Orlando S (2019). Sustainable, circular and innovative value chains using growing substrata alternative to peat for the cultivation of nutraceutical species. 9th International Scientific Conference Rural Development 2019. At: Kaunas, Lithuania. |
|
|
Gupta C, Prakash D, Gupta S, Nazareno MA (2019). Role of Vermicomposting in Agricultural Waste Management. Sustainable Green Technologies for Environmental Management: Springer pp. 283-295. |
|
|
Hajizadeh HS, Heidari B, Bertoldo G, Della Lucia MC, Magro F, Broccanello C, Baglieri A, Puglisi I, Squartini A, Campagna G, Concheri G, Nardi S, Stevanato P (2019). Expression Profiling of Candidate Genes in Sugar Beet Leaves Treated with Leonardite-Based Biostimulant. High-Throughput 8(4):18. |
|
|
Jabeen N, Ahmad R (2017). Growth response and nitrogen metabolism of sunflower (Helianthus annuus L.) to vermicompost and biogas slurry under salinity stress. Journal of Plant Nutrition 40(1):104-114. |
|
|
Javaid T, Farooq MA, Akhtar J, Saqib ZA, Anwar-ul-Haq M (2019). Silicon nutrition improves growth of salt-stressed wheat by modulating flows and partitioning of Na+, Cl− and mineral ions. Plant Physiology and Biochemistry 141:291-299. |
|
|
Jnandabhiram C, Sailen PB (2012). Water stress effects on leaf growth and chlorophyll content but not the grain yield in traditional rice (Oryza sativa Linn.) genotypes of Assam, India II. Protein and proline status in seedlings under PEG induced water stress. American Journal of Plant Sciences pp. 971-980. |
|
|
Kar S, Chandel A, Pant K (2019). Effect of Different Tree Spacings of Grewia optiva and Combined Doses of Poultry Manure and Vermicompost on Growth and Yield of Bell Pepper under Agrisilviculture System. International Journal of Current Microbiology and Applied Sciences 8(1):2213-23. |
|
|
Liu R, Gu M, Huang L, Yu F, Jung SK, Choi HS (2019). Effect of pine wood biochar mixed with two types of compost on growth of bell pepper (Capsicum annuum L.). Horticulture, Environment and Biotechnology 60(3):313-319. |
|
|
Lutts S, Kinet J, Bouharmont J (1996). Effects of salt stress on growth, mineral nutrition and proline accumulation in relation to osmotic adjustment in rice (Oryza sativa L.) cultivars differing in salinity resistance. Plant Growth Regulation 19(3):207-218. |
|
|
Mohapatra D, Sahoo K, Sannigrahi A (2019). Impact of quantity of paper mill wastes on vermi-conversion time and quality of product. |
|
|
Moridi A, Zarei M, Moosavi AA, Ronaghi A (2019). Influence of PGPR-enriched liquid organic fertilizers on the growth and nutrients uptake of maize under drought condition in calcareous soil. Journal of Plant Nutrition 42(20):2745-2756. |
|
|
Morton MJ, Awlia M, Alâ€Tamimi N, Saade S, Pailles Y, Negrão S, et al (2019). Salt stress under the scalpel-dissecting the genetics of salt tolerance. The Plant Journal 97(1):148-63. |
|
|
Ndegwa PM, Thompson SA (2001). Integrating composting and vermicomposting in the treatment and bioconversion of biosolids. Bioresource Technology 76:107-112. |
|
|
Nedjimi B, Daoud Y (2006). Effect of Na 2 SO 4 on the growth, water relations, proline, total soluble sugars and ion content of Atriplex halimus subsp. schweinfurthii through in vitro culture. Anales de Biología; Servicio de Publicaciones de la Universidad de Murcia 28:35-43. |
|
|
Pandyaswargo AH, Premakumara DGJ (2014). Financial sustainability of modern composting: the economically optimal scale for municipal waste composting plant in developing Asia. International Journal of Recycling of Organic Waste in Agriculture 3(3):4. |
|
|
ParaÄ‘iković N, Teklić T, Zeljković S, Lisjak M, Špoljarević M (2019). Biostimulants research in some horticultural plant species-A review. Food and Energy Security 8(2):e00162. |
|
|
Piovan MJ, Pratolongo P, Donath TW, Loydi A, Eckstein RL (2019). |
|
|
Germination Response to Osmotic Potential, Osmotic Agents, and Temperature of Five Halophytes Occurring along a Salinity Gradient. International Journal of Plant Sciences 180(4):345-355. |
|
|
Rorat A, Vandenbulcke F (2019). Earthworms converting domestic and food industry wastes into biofertilizer. Industrial and Municipal Sludge: Elsevier pp. 83-106. |
|
|
Saghafi D, Delangiz N, Lajayer BA, Ghorbanpour M (2019). An overview on improvement of crop productivity in saline soils by halotolerant and halophilic PGPRs. 3 Biotech 9(7):261. |
|
|
Schiavon M, Ertani A, Francioso O, Nardi S (2019). Manure fertilization gives high-quality earthworm coprolites with positive effects on plant growth and N metabolism. Agronomy 9(10):659. |
|
|
Shahi C, Bargali K, Bargali S (2015). Assessment of salt stress tolerance in three varieties of rice (Oryza sativa L.). Journal of Progressive Agriculture 6(1):50-6. |
|
|
Shelke DB, Nikalje GC, Chambhare MR, Zaware BN, Penna S, Nikam TD (2019). Na+ and Cl− induce differential physiological, biochemical responses and metabolite modulations in vitro in contrasting salt-tolerant soybean genotypes. 3 Biotech 9(3):91. |
|
|
Singh G, Biswas D, Marwaha T (2010). Mobilization of potassium from waste mica by plant growth promoting rhizobacteria and its assimilation by maize (Zea mays) and wheat (Triticum aestivum L.): a hydroponics study under phytotron growth chamber. Journal of Plant Nutrition 33(8):1236-1251. |
|
|
Vandoorne B, Mathieu AS, Van den Ende W, Vergauwen R, Périlleux C, Javaux M, Lutts S (2012). Water stress drastically reduces root growth and inulin yield in Cichorium intybus (var. sativum) independently of photosynthesis. Journal of Experimental Botany 63(12):4359-4373. |
|
|
Vibhuti CS, Bargali K, Bargali S (2015). Seed germination and seedling growth parameters of rice (Oryza sativa L.) varieties as affected by salt and water stress. Indian Journal of Agricultural Sciences 85(1):102-8. |
|
|
Wang WB, Kim YH, Lee HS, Kim KY, Deng XP, Kwak SS (2009). Analysis of antioxidant enzyme activity during germination of alfalfa under salt and drought stresses. Plant Physiology and Biochemistry 47(7):570-577. |
|
|
Wang WY, Chai WW, Zhao CY, Rowland O, Wang BS, Song X, Liu YQ, Ma Q, Wang SM (2019). Under drought conditions NaCl improves the nutritional status of the xerophyte Zygophyllum xanthoxylum but not of the glycophyte Arabidopsis thaliana. Journal of Plant Nutrition and Soil Science 182(4):597-606. |
|
|
Yakhin OI, Lubyanov AA, Yakhin IA, Brown PH (2017). Biostimulants in plant science: a global perspective. Frontiers in Plant Science 7:2049. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


